










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.34 no.2 Bogotá May/Aug. 2016
https://doi.org/10.15446/agron.colomb.v34n2.57193
Doi: http://dx.doi.org/10.15446/agron.colomb.v34n2.57193
Fruit growth characterization of the tomato (Solanum lycopersicum L.) hybrid Ichiban' grown under cover
Caracterización del crecimiento del fruto de tomate (Solanum lycopersicum L.) híbrido 'Ichiban' cultivado bajo cubierta
Pedro José Almanza-Merchán1, Yuly Alexandra Arévalo2, Germán Eduardo Cely R.2, Elberth Hernando Pinzón1, and Pablo Antonio Serrano C.2
1 Group of Agricultural Research (GIA), Faculy of Agricultural Sciences, Universidad Pedagogica y Tecnologica de Colombia (UPTC). Tunja (Colombia). pedro.almanza@uptc.edu.co
2 Research Group GIPSO, Faculty of Agricultural Sciences, Universidad Pedagogica y Tecnologica de Colombia (UPTC). Tunja (Colombia).
Received for publication: 28 April, 2016. Accepted for publication: 30 June, 2016.
ABSTRACT
The tomato (Solanum lycopersicum L.) is one of the more important crops that are consumed worldwide. In Colombia, the department of Boyaca is characterized as being the biggest producer of tomatoes, with production centered in the municipalities of Sutamarchan, Sachica, Santa Sofía and Villa de Leyva, where different cultivars and hybrids are produced that present different physiological behaviors, resulting in changes in conventional agronomic management. This study aimed to evaluate the growth and development of fruits from the tomato hybrid Ichiban under greenhouse conditions. This study evaluated the parameters of polar and equatorial diameters, diameter ratio, fresh mass and dry mass, and absolute and relative growth rates. The results were subjected to a descriptive analysis. Furthermore, the growth model was determined. The Ichiban hybrid had a simple sigmoid behavior, common in fleshy fruits, while the fresh mass and dry mass were fitted to a logistic, nonlinear model. Anthesis was seen at 50 days after transplant and the fruits reached physiological maturity 49 days after anthesis; during this period, 510.2 heating degree-days were accumulated. All of the variables presented significant differences (P≤0.01) and coefficients of determination above 0.9, indicating that the models are suitable for explaining the studied biological phenomena.
Key words: Growth analysis, growth rate, logistic model.
RESUMEN
Dentro de las hortalizas consumidas en el mundo, el tomate (Solanum lycopersicum L.) es una de las de mayor importancia. En Colombia, el departamento de Boyacá se caracteriza por ser el mayor productor de tomate centrando su producción en los municipios de Sutamarchan, Sachica, Santa Sofía y Villa de Leiva, en donde se producen diferentes cultivares o híbridos, que presentan diferente comportamiento fisiológico, lo que implica cambios en el manejo agronómico convencional. El objetivo del trabajo consistió en evaluar el comportamiento del crecimiento y desarrollo del fruto de tomate híbrido Ichiban bajo condiciones de invernadero, para lo cual se evaluaron parámetros como diámetro polar y ecuatorial, relación entre diámetros, masa fresca y seca, y tasa absoluta y relativa de crecimiento, los resultados fueron sometidos a un análisis descriptivo. Además, se determinó el modelo de crecimiento. Se encontró que el hibrido Ichiban presenta un comportamiento sigmoide simple común en frutos carnosos, los diámetros polar y ecuatorial se ajustaron a modelos polinómicos, mientras que la masa seca y fresca se ajustó a un modelo no lineal de tipo logístico. Se presento antesis a los 50 días después del trasplante (ddt) y los frutos alcanzaron la madurez fisiológica 49 días después de la antesis, durante este período se acumularon 510,2 GDD Todas las variables presentaron diferencias estadísticas (P≤0,01) y coeficientes de determinación superiores a 0,9, indicando, que los modelos son adecuados para explicar los fenómenos biológicos estudiados.
Palabras clave: Análisis de crecimiento, tasas de crecimiento, modelo logístico.
Introduction
The tomato (Solanum lycopersicum L.) is the second most consumed vegetable crop, after the potato (Baracaldo et al., 2014). While tremendous improvements in efficiency and cost of production have been made in the tomato processing industry world-wide over the past 100 years, there is still opportunity for innovation (Barrett, 2015). In Colombia, 18 departments cultivate tomatoes, with 94.1% of the domestic production concentrated in Boyaca, Antioquia, Norte de Santander, Cundinamarca, Nariño, Valle de Cauca, Huila, Risaralda and Caldas (Miranda et al., 2009). The production in Boyaca is distributed in the municipalities of Sachica, Sutamarchan, Santa Sofía, Tenza, Pachavita and Villa de Leyva; the latter is the biggest tomato producer at the domestic level (Castellanos et al., 2014).
The tomato is considered a species that has a high number of genotypes from conventional breeding programs, resulting in variation in the morphological, biochemical and physiological behaviors of the plants. Knowledge on these aspects determines the biological efficiency and suitable agronomic management for achieving the best genetic and agronomic potential for new cultivars (Ardila et al., 2011). The department of Boyaca, since it is the biggest producer, is constantly introducing new cultivars and hybrids that have unknown physiological behaviors under the agroecological conditions of the zones where they are cultivated. Among these, the Ichiban hybrid has characteristics such as semi-flat, round fruits that are extra-firm, uniform, smooth with a good appearance, green in color with a semi-dark to intense-red shine at maturation, and weights that oscillate between 160 and 250 g per fruit (Semillas Magna, 2015).
Analyzing the growth and development of species is essential for planning different agronomic practices in crops. According to Rodríguez et al. (2006), this can be carried out taking into account the time intervals for generating calculations, which provides an analysis through a simple computation. Furthermore, a series of growth rates has been created that can be used to interpret behaviors in the best way possible; these rates are calculated from the dry material accumulated by the organ of interest and analyzed with defined time intervals, chronological or physiological (Hunt, 1990). According to this author, among these, the more representative ones in terms of the fruit growth dynamic are the absolute growth rate (AGR) and the relative growth rate (RGR). The AGR indicates the change in size per unit of time, while the RGR expresses the variation rate of the size per initial size unit. The growth conditions for plants have been improved significantly through understanding crop physiology in combination with new technologies (Marcelis et al., 2014).
The growth of any species or organ is calculated over time, which plays a very important role. However, when calculating from the net chronological point of view (hours, days, years etc.), this does not directly demonstrate the growth behavior of plants or some plant organs from the biological perspective because factors such as temperature are not taken into account. The use of thermal-time (growing day degrees) increases the precision in determining the duration of the development and growth cycle of the plant or organ (Ruiz et al., 1998) because it takes into account the threshold temperatures (maximum and minimum) that control the development and growth of the species for the analysis (Ruiz-Corral et al., 2002), even more so if the fact that crop productivity is strongly driven by climatic variables is taken into account. Some crops, such as the tomato, will suffer heavily with the predicted climate change for the current century due to their summer growing cycle (Garofalo and Rinaldi, 2015). Studies carried out on these aspects have demonstrated that temperatures that exceed these limits impede plant development (Trudgill et al., 2005). In the case of the tomato, the minimal threshold temperature or base temperature is found at 10°C (Zotarelli et al., 2009; Ardila et al., 2011). Considering the above, this study aimed to characterize, in thermal-time, the growth of fruits from the tomato hybrid 'Ichiban' cultivated under a plastic cover with the agroecological conditions of the municipality of Sachica (Boyaca).
Materials and methods
This study was conducted between the months of June and September of 2014 on the Andalucia Farm, located in the Ritoque district of the municipality of Sachica (Boyaca-Colombia) and found at 05° 35'07" N and 73° 32'42" W with an altitude of 2,150 m a.s.l.
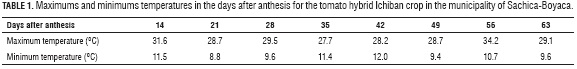
A 1,960 m2 lot was covered with a plastic cover (greenhouse) and cultivated with a commercial 'Ichiban' tomato crop, characterized by a high percentage of fruits with a mean weight of 250 g, uniform maturity, homogenous coloration and excellent postharvest behavior. The greenhouse had a mean temperature of 19.5°C, with daily temperature averaging maximums of 29.7°C and minimums of 10.4°C (Tab. 1).
The plants were sown at a distance of 0.3 m between the plants and 1.1 m between the rows, with a cultivation density of 30,303 plants/ha. The fertilization program was established through an analysis of the soil and was implemented through fertigation. The phytosanitary management and other cultivation tasks were carried out with the technical recommendations of the plant physiology laboratory of the Universidad Pedagogica y Tecnologica de Colombia (UPTC), Tunja.
Three plants were taken per sampling unit, for a total of 24 plants, randomly marking four clusters in each one. Of these, 30 flowers in the anthesis stage were marked. After fruit set, five fruits were taken from each cluster for the respective physical evaluations, which were taken immediately under cold conditions to the UPTC laboratory in Tunja. The measurements were taken every 8 d for a total of eight samplings. The sampling started 14 d after anthesis (DAA) and lasted until the fruits reached grade 5 according to NTC 1103-1 (Icontec, 1995) with a completely red fruit epidermis color, 59 (DAA), thereby completely encompassing the development and growth of the fruits.
In order to determine the thermal-time (accumulation of heating degree-days (GDD)), a humidity/temperature Datalogger RHT-20 (Extech Instruments Waltham, MA) was installed that registered the minimum and maximum temperatures from anthesis to harvest. With the obtained data, the GDD was calculated with Eq. 1, used by Rodríguez and Flórez (2006):
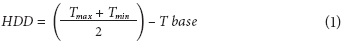
where T max is the maximum daily air temperature; T min is the minimum daily air temperature; and T base is the minimum temperature for the metabolic processes of the plant organs. In the particular case of tomato fruits, the latter is 10°C when using studies conducted under conditions similar to those of the reference, Zotarelli et al. (2009) and Ardila et al. (2011).
The variables evaluated to determine the growth of the fruits included: fresh mass (g), obtained immediately after collecting the samples; dry mass of the fruits (g), obtained after subjecting the fruits to 75°C until reaching constant weight, taken with a 0.01 g precision balance (Ohaus, model AS-120S, Parsippany, NJ); and equatorial and polar diameters (mm), taken with a 0.05 mm precision digital vernier caliper (Mitutoyo Corporation, Kanagawa Japan). With these data, the absolute growth rate (AGR) and the relative growth rate (RGR) were calculated with the methodology employed by Carranza et al. (2009), using a functional approach for this model. The obtained data were analyzed with descriptive statistics by calculating the arithmetic mean and the standard error. The logistic, nonlinear model was created with SAS® v.9.2e (SAS Institute Inc., Cary, NC).
Results and discussion
The statistical analysis demonstrated that, in all of the evaluated variables, there were significant differences (P<0.01) for the employed models. The variables fresh mass and dry mass were fitted to a logistic model; the polar and equatorial diameters were fitted to a polynomial model; and the polar and equatorial diameter ratio was fitted to a linear model. All of the parameters presented coefficients of determination (R2) higher than 0.9, indicating a high goodness of fit in each of the models for explaining the studied biological phenomena as a function of the chronological time and thermal-time (GDD) (Tab. 2). Ardila et al. (2011) found similar results when studying three tomato hybrids. In addition, Casierra-Posada et al. (2007) fitted the studied variables to a cubic model, except for the AGR, which was fitted to an exponential model. However, the response was similar to the one found in the present study.

Growth and development
The tomato hybrid Ichiban, under the studied conditions, presented anthesis at 50 d after transplant (dat) and the fruits reached physiological maturity 49 DAA, seen as the moment when the fruits had developed all of their organo-leptic characteristics; during this period 510.2 GDD were accumulated. These results are similar to those reported by Ardila et al. (2011), who studied three tomato hybrids and found that the accumulation was higher than 700 GDD when obtaining the physiological maturity of the fruits, demonstrating the envirogenomic relationship with the growth of fruits for each of the hybrids of a species. Casierra-Posada et al. (2009), in studies carried out on the tomato hybrids Sofía, Bravona and Granitio under similar conditions, found that physiological maturity was reached at 64 DAA, 5 d later than the hybrid in this study. This led to the conclusion that the studied hybrid could be considered an early-ripening tomato with very good characteristics of adaptation to zones over 2,200 m a.s.l. and a mean temperature of 24°C under a covered production system.
According to De Koning (1994) and Islam (2011), temperature accelerates the fruit development rate, i.e. a high temperature results in an earlier attainment of the maximum growth rate and fruit maturation, in agreement with the present study. Therefore, temperature is one of the more important ambient factors that determine the growth period of fruits, exercising influence on the sink strength of fruits through an increase in the development rate and can be represented through an accumulation of degree-days throughout the fruit growth cycle, from fruit set to maturation (Rodríguez-Fuentes et al., 2012), which confirms the effect of temperature seen in the greenhouse during this research.
According to Jaramillo et al. (2013), tomato production depends mainly on two factors: the physiological condition of the plant and external conditions thereof. The external conditions include: soil and air moisture, radiation and temperature, which influence the physiological condition of the plant. The development of the cluster, flower, pollen viability, fruit shape, plant growth and fruit set of the fruit are affected by temperature and thus affect production (Hatfield and Prueger, 2015). Therefore, the temperature and climate are the main influential factors in most developmental stages and physiological processes of the plant, this depends on the thermal value reached in the greenhouse at each critical period. As the tomato crop of temperate climate, this benefits from mild temperatures. Fluctuations of day-night temperatures (thermal period) or temperature swings (Almanza et al., 2012) benefit growth and development in the case of the tomato, with differences of 6 and 7°C day/night (Maroto, 2000); in the present study, 19°C temperature variations generated by the greenhouse environment were seen.
Likewise, an effect from the temperature oscillations, which can occur over short periods of time, was observed on the sink strength of the vegetative organs through modifications of the metabolic reaction rates, meaning biochemical reactions are accelerated with increases in temperature (De Koning, 1994; Gary et al., 2003).
Fruit diameter
The polar and equatorial diameters of the 'Ichiban' hybrid were fitted to a polynomial model, which described three growth stages: an initial cellular division phase, which coincided with an accumulation of 50 HDD; followed by a rapid accumulation of the diameter due to processes of cellular elongation, from 50 to 300 HDD; and a final phase in which the fruits reached maximum growth when they were harvested with a total accumulation of 510 HDD. The fruits achieved the maximum polar and equatorial diameters at 450 HDD (42 DAA); this behavior was fitted to a simple sigmoid model, which is common in fleshy fruits. In Fig. 1A, the primary low growth phase is not evident because, in order to clearly observe changes, more closely spaced samples must be taken, as indicated by Casierra and Cardoso (2009).
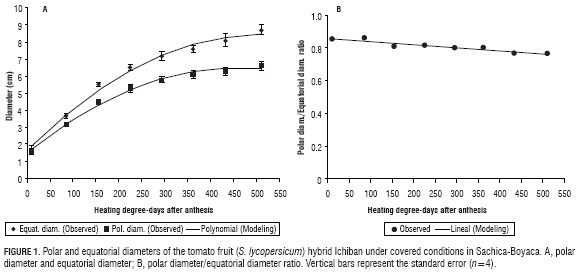
An increase in the diameters and in the volume of tomato fruits depends on the epidermal extensibility (Thompson, 2001). While the epidermal cells divide throughout the entire development of the fruits, the division in the pericarp is limited to a short period of fruit development and is located in the external tissues around which there is vascular tissue and, in the hypodermis once the cellular division is over, cellular expansion starts in order to increase the fruit size (Bertin, 2005).
The ratio between the polar diameter and equatorial diameter was fitted to a linear model with a coefficient of determination of 0.91. The Ichiban hybrid was observed to have a fruit shape that was determined almost at the start of fruit set (Fig. 1B). The fruit, when starting growth, presented a spherical shape in order to increase the polar diameter; therefore, a slightly elongated fruit shape was observed, which was lost during development, resulting in a final, slightly flattened shape that is characteristic of this hybrid. Some tomato hybrids cultivated under similar conditions as the present study present a differential behavior for this parameter, with oblong or flattened fruits. This indicates that fruit shape, in addition to being linked to the solute and seed content, also results from the genetic characteristics of each hybrid (Casierra-Posada et al., 2010).
Fruit shape has a high correlation with the number of seeds, which is seen in some species of the Solanacea family (Peña et al., 2010). Developing seeds is a strong sink for carbohydrates that are provided as sucrose via the phloem (Öpik and Rolfe, 2005). The shape that is acquired by the fruit during the primary growth stages results from the effect of the growth regulators produced by immature seeds, in which, high quantities of abscisic acid (ABA) have been found in the pericarp, in the axils, and in the loculal tissue during the early growth stages of the fruits; however, large quantities of indole acetic acid (AIA) were subsequently seen, which determine the growth rhythm and the fruit shape, with the flow of photosynthates and water being the final determinates of fruit growth, both for size and shape (Kojima, 2005).
Fresh mass and dry mass of the fruits
The variables of fresh mass and dry mass of the fruits were fitted to a logistic, nonlinear model, describing the simple sigmoid type growth found in fleshy fruits (Salisbury and Ross, 1994; Agusti, 2013). These results agree with those reported by Casierra-Posada et al. (2007) for tomatoes under conditions similar to those of the present study. It was observed that the fruits rapidly increased the accumulation of dry mass starting at an accumulation of 90 HDD (7 DAA), achieving the maximum dry mass with the accumulation of 300 HDD (29 daa), followed by entrance into the maturation phase. 'Ichiban' tomato hybrid fruits present three growth phases as described by Opara (2000) both for the fresh mass and the dry mass: an initial phase with a low mass quantity, possibly due to the fact that this phase is dominated by cellular division; followed by fruit filling with a predominance of cellular elongation and a possible decrease in respiration (Kays, 2004); and a final phase of slow growth (Fig. 2), almost imperceptible; during this phase, fruits begin to present physical, physiological, and biochemical changes that are characteristic of the fruit maturation process (Seymour et al., 2013).
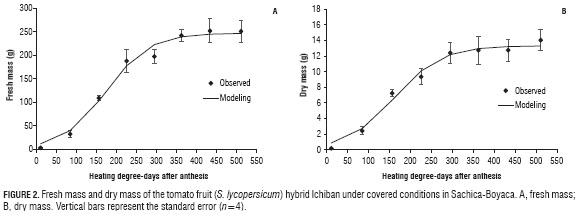
The dry mass accumulated by tomato fruits mostly results from the photoassimilates produced in the leaves and transported to the fruits, according to the dynamic of the source/sink ratio. Therefore, the sink strength of an organ is the potential demand or capacity of the organ for the accumulation of assimilates (Marcelis and Heuvelink, 2007). As such, fruits are established as the principal sink organs and can compete with others, such as the vegetative organs, for the available assimilates (Peil and Gálvez, 2005). It is for this reason that these effects on yield are mainly due to a higher CO2 concentration (Marcelis et al., 2014). The tomato, in its early growth stages, accumulates a minimum amount of dry matter, but this is high when observed in the last stage of full production. This is due to the high metabolic demand that occurs during fruit filling and occurs even when there is a phenotypic difference between cultivars of the same species. This makes, that the order of priority in the allocation of assimilates change with the phenological stage (Casierra-Posada et al., 2007; Andriolo and Falcão, 2000; Hernández et al., 2014).
Absolute growth rate and relative growth rate
In terms of the absolute growth rate (AGR), based on the dry mass of the Ichiban hybrid tomato fruits, a bell-shaped, typical behavior was observed, characteristic of this fruit type, demonstrating an increase from the start of GDD accumulation in the fruit until reaching 150 GDD (16 DAA), with a subsequent decrease until 400 GDD (89 DAA), then stabilizing until harvest, accumulating 510.2 GDD in the chronological time of 49 DAA (Fig. 3A).
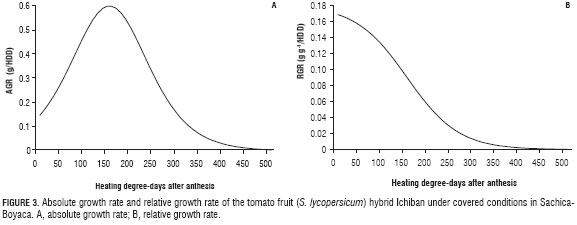
According to Hunt (2003), the AGR is a reliable parameter in terms of the instantaneous accumulation of dry mass, as well as the sink strength of fruits, making it a very important parameter for the planning of agricultural tasks that must be carried out in crops because, when presenting the highest AGR, the fruits are more sensitive to influences from activities such as foliar fertilization, providing water and a higher supply of solar energy (Casierra-Posada and Cardozo, 2009). Likewise, the AGR can be used to investigate the ambient effect on growth and development, which results in a better understanding of the nature of the plant-environment interaction (Mora-Aguilar et al., 2005). Studies carried out by Mazorra et al. (2003) indicated that the AGR, as a growth index used in fruits, is very useful for determining exact values for fruit growth over time, comparing locations and fruit placement on the plants, but demonstrates limitations for evaluating the efficiency of plants.
The relative growth rate (RGR) for the Ichiban hybrid under the conditions of the present study demonstrated a behavior similar to the one reported by Ardila et al. (2011), presenting a constant decrease over time (Fig. 3B) because the RGR is an index of the efficiency that expresses growth in terms of an increased rate in size per size unit over time. In the case of tomato fruits, it indicates the gains obtained as a sink, with the majority of the fruit photoassimilates coming from the leaves (Casierra-Posada and Cardozo, 2009) and a smaller amount coming from the photosynthesis carried out by immature fruits. It is for this reason that the comparison of the growth curves of the AGR and RGR turn out to be important for selecting promising materials for the different environments in which they develop (Gómez de Enciso, 2012), especially given the fact that light, as an important factor in all stages of growth, is reflected in the growth rates and production factor (Flórez et al., 2012).
Conclusions
The Ichiban hybrid presented a simple sigmoid behavior that is common in fleshy fruits. The polar and equatorial diameters were fitted to polynomial models, while the fresh mass and the dry mass were fitted to a logistic, nonlinear model. All of the variables presented statistical differences (P≤0.01) and coefficients of determination above 0.9, indicating that the models are suitable for explaining the studied biological phenomena. The analysis of the growth and development for the thermal-time led to the conclusion that the studied hybrid could be considered an early-ripening tomato with good characteristics of adaptation to zones over 2,200 m a.s.l. and a mean temperature of 24°C under a covered production system. The logistic, nonlinear model facilitated a functional analysis with a high goodness of fit, indicating that it is suitable for estimating biological phenomena such as fruit growth and development.
Literature cited
Agusti, M. 2013. Crecimiento y maduración del fruto. pp. 519-536. In: Azcón-Bieto, J. and M. Talón (eds.). Fundamentos de fisiología vegetal. 2nd ed. Interamericana-McGraw-Hill, Madrid. [ Links ]
Almanza, P.J., P.A. Serrano, and G. Fischer. 2012. Manual de viticultura tropical. Universidad Pedagógica y Tecnológica de Colombia, Tunja, Colombia. [ Links ]
Andriolo, J.L. and L.L. Falcão. 2000. Efeito da poda de folhas sobre a acumulação de materia seca e sua repartição para os frutos do tomateiro cultivado en ambiente protegido. Rev. Bras. Agrometeor. 8, 75-83. [ Links ]
Ardila, G., G. Fischer, and H. Balaguera. 2011. Caracterización del crecimiento del fruto y producción de tres híbridos de tomate (Solanum lycopersicum L.) en tiempo fisiológico bajo invernadero. Rev. Colomb. Cienc. Hortic. 5, 44-56. Doi: 10.17584/rcch.2011v5i1.1252. [ Links ]
Baracaldo, A., R. Carvajal, A.P. Romero, A.M. Prieto, F.J. García, G. Fischer, and D. Miranda. 2014. El anegamiento afecta el crecimiento y producción de biomasa en tomate chonto (Solanum lycopersicum L.), cultivado bajo sombrío. Rev. Colomb. Cienc. Hortic. 8, 92-102. Doi: 10.17584/rcch.2014v8i1.2803. [ Links ]
Barrett, D.M. 2015. Future innovations in tomato processing. Acta Hortic. 1081, 49-55. Doi: 10.17660/ActaHortic.2015.1081.3. [ Links ]
Bertin, N. 2005. Analysis of the tomato fruit growth response to temperature and plant fruit load in relation to cell division, cell expansion and DNA end reduplication. Ann. Bot. 95,439-447. Doi: 10.1093/aob/mci042. [ Links ]
Castellanos, J.N., J.K. Moreno, and J.C. Murcia. 2014. Plan de negocio para la creación de una planta de poscosecha y transformación de tomate en el alto Ricaurte de Boyacá. Undergraduate thesis. Facultad de Ciencias Administrativas y Contables, Universidad de la Salle, Bogotá [ Links ].
Casierra-Posada, F., O.J. Alvarez, and N. Luque. 2010. Calidad de frutos en tomate (Solanum lycopersicum L. cv. Rocío) producidos bajo coberturas reflectiva y plástica. Rev. Colom. Cien. Hort. 4, 67-80. Doi: 10.17584/rcch.2010v4i1.1226. [ Links ]
Casierra-Posada, F. and M.C. Cardozo. 2009. Análisis básico del crecimiento en frutos de tomate (Lycopersicon esculentum Mill, cv. 'Quindío') cultivados a campo abierto. Rev. Fac. Nal. Agr. Medellín 62, 4815-4822. [ Links ]
Casierra-Posada, F., M. Cardozo, and J. Cárdenas-Hernández. 2007. Análisis del crecimiento en frutos de tomate (Lycopersicon esculentum Mill.) cultivados bajo invernadero. Agron. Colomb. 25, 299-305. [ Links ]
Carranza, C., O. Lancheros, D. Miranda, and B. Chaves. 2009. Análisis de crecimiento de lechuga (LactucasativaL.) "Batavia" cultivada en un suelo salino de la Sabana de Bogotá. Agron. Colomb. 27, 41-48. [ Links ]
De Koning, A.N.M. 1994. Development and dry matter distribution in glasshouse tomato: a quantitative approach. PhD thesis, Wageningen Agricultural University, Wageningen, The Netherlands. [ Links ]
Flórez, L.E., G. Gónzalez, S.P. Pulido, K. Wyckhuys, H. Escobar. C. Salamanca, A. Zamudio, J. Jiménez, R. Gil, L.S. Fuentes. N. Niño, L. Fuentes, and C. Bojacá. 2012. Tomate (Solanum lycopersicum L.). pp. 498-528. In: Pinzón, H. (ed.). Manual para el cultivo de hortalizas. Produmedios, Bogotá [ Links ].
Gary, C., P. Baldet, N. Bertin, C. Devaux, M. Tchamitchian, and P. Raymond. 2003.Time-course of tomato whole-plant respiration and fruit and stem growth during prolonged darkness in relation to carbohydrate reserves. Ann Bot. 91, 429-438. Doi: 10.1093/aob/mcq037. [ Links ]
Garofalo, P. and M. Rinaldi. 2015. Impact of climate change and adaptation strategies on irrigation and tomato productivity: a long-term case study in mediterranean environment. Acta Hortic. 1081, 89-96. Doi: 10.17660/ActaHortic.2015.1081.8. [ Links ]
Gómez de Enciso, C. 2012. Ecofisiología de hortalizas. pp. 55-71. In: Pinzón, H. (ed.). Manual para el cultivo de hortalizas. Produmedios, Bogotá [ Links ].
Hatfield, J.L. and J.H. Prueger. 2015. Temperature extremes: effect on plant growth and development. Weather and Climate Extremes 10, 4-10. Doi: 10.1016/j.wace.2015.08.001. [ Links ]
Hernández, M., M. Chailloux, V., Moreno, M. Mojena, and J.M. Salgado. 2014. Relaciones nitrógeno-potasio en fertirriego para el cultivo protegido del tomate (Solanum lycopersicum L.) en la época de primavera-verano. Cult. Trop. 35, 106-105. [ Links ]
Hunt, R. 1990. Basic growth analysis. Plant growth analysis for beginners. Unwin Hyman, Boston, MA. Doi: 10.1007/978-94-010-9117-6. [ Links ]
Hunt, R. 2003. Growth analysis, individual plants. pp. 579-588. In: Thomas, B., D.J. Murphy and D. Murray (eds.). Encyclopedia of applied plant sciences. Academic Press, London. Doi: 10.1016/B0-12-227050-9/00028-4. [ Links ]
Icontec. 1995. Norma técnica colombiana NTC 1103-1, Industrias alimentarias. Tomate de mesa. Instituto Colombiano de Normas Técnicas y Certificación, Bogotá [ Links ].
Islam, M.T. 2011. Effect oftemperature on photosynthesis, yield attributes and yield oftomato genotypes. Int. J. Expt. Agric. 2, 8-11. [ Links ]
Jaramillo, N., G.D. Sánchez, V.P. Rodríguez, P.A. Aguilar, M.A. Zapata, and M. Guzmán. 2013. Tecnología para el cultivo del tomate bajo condiciones protegidas. Ministerio de Agricultura y Desarrollo Rural; Corpoica, Bogotá [ Links ].
Kays, S. 2004. Postharvest biology. Exon Press, Athens, GA. [ Links ]
Marcelis, L.F.M., F. Buwalda, J.A. Dieleman, T.A. Dueck, A. Elings, A. de Gelder, S. Hemming, F.L.K. Kempkes, T. Li, F. Van Noort, and P.H.B. de Visser. 2014. Innovations in crop production: a matter of physiology and technology. Acta Hortic. 1037, 39-45. Doi: 10.17660/ActaHortic.2014.1037.1. [ Links ]
Marcelis, L.F.M. and E. Heuvelink. 2007. Concepts of modelling carbon allocation among plant organs. pp. 103-111. In: Vos, J., L.F.M. Marcelis, P.H.B. de Visser, P.C. Struik, and J.B. Evers. (eds.). Functional-structural plant modelling in crop production. Springer, Amsterdam, The Netherlands. [ Links ]
Maroto, J.V. 2000. Horticultura herbácea especial. Mundi-Prensa, Madrid. [ Links ]
Mazorra, M., A. Quintana, D. Miranda, G. Fischer, and B. Cháves. 2003. Análisis sobre el desarrollo y la madurez fisiológica del fruto de la uchuva (Physalis peruviana L.) en la zona de Sumapaz (Cundinamarca). Agron. Colom. 21, 175-189. [ Links ]
Miranda, D., G. Fischer., C. Carranza., M. Rodríguez., O. Lanchero, and J.C. Barrientos. 2009. Characterization of productive systems of tomato (Solanum lycopersicum L.) in producing zones of Colombia. Acta Hort. 821, 35-46. Doi: 10.17660/ActaHortic.2009.821.2. [ Links ]
Mora-Aguilar, R., J. Ortíz-Cereceres, M. Mendoza, A. Rivera-Peña, H. Lozoya-Saldaña, and M. Colinas-León. 2005. Cinética de la acumulación y distribución de biomasa en genotipos de papa establecidos en condiciones de secano. Rev. Chapingo Ser. Hortic. 11, 135-142. [ Links ]
Rodríguez-Fuentes, H., J.A. Vidales-Contreras, A.I. Luna-Maldonado, J.C. Rodríguez-Ortiz. 2012. Total growth of tomato hybrids under greenhouse conditions. pp. 63-72. In: Luna M., A.I. (ed.). Horticulture. InTech, Rijeka, Croatia. [ Links ]
Rodríguez, W.W. and V. Flórez. 2006. Comportamiento fenológico de tres variedades de rosas rojas en función de la acumulación de la temperatura. Agron. Colomb. 4, 247-257. [ Links ]
Rodríguez, M., H. Arjona, and H. Campos. 2006. Caracterización fisicoquímica del crecimiento y desarrollo de los frutos de feijoa (Acca sellowiana Berg.) en los clones 41 (Quimba) y 8-4. Agron. Colomb. 24, 54-61. [ Links ]
Opara, L.U. 2000. Fruit growth measurement and analysis. Hort. Rev. 24, 373-431. Doi: 10.1002/9780470650776.ch8. [ Links ]
Öpik, H. and S. Rolfe. 2005. The physiology of flowering plants. 4th ed. Cambridge University Press, Cambridge, UK. Doi: 10.1017/CBO9781139164450. [ Links ]
Peil, R. and J. Gálvez. 2005. Reparto de materia seca como factor determinante de la producción de las hortalizas de fruto cultivadas en invernadero. Rev. Bras. Agrociênc. 11, 5-11. Doi: 10.18539/cast.v11i1.1171. [ Links ]
Peña, J., J. Ayala, G. Fischer, B. Cháves, J. Cárdenas, and P.J. Almanza. 2010. Relaciones semilla-fruto en tres ecotipos de uchuva (Physalis peruviana L.). Rev. Colomb. Cienc. Hortic. 4, 43-54. Doi: 10.17584/rcch.2010v4i1.1224. [ Links ]
Ruiz-Corral, J., H. Flores-López, J. Ramírez-Díaz, and D. González-Equiarte. 2002. Temperaturas cardinales y duración del ciclo de madurez del híbrido de maíz H-311 en condiciones de temporal. Agrociencia 36, 569-577. [ Links ]
Ruiz, C., J. Sánchez, and M. Goodman. 1998. Base temperature and heat unit requirement of 49 mexican maize races. Maydica 43, 277-282. [ Links ]
Salisbury, F.B. and C.W. Ross. 1994. Fisiología vegetal. Grupo Editorial Iberoamérica, Mexico DF. [ Links ]
Seymour, G.B., L. 0stergard, N.H. Chapman, S. Knapp, and C. Martín. 2013. Fruit development and ripening. Annu Rev Plant Biol. 64, 219-241. Doi: 10.1146/annurev-arplant-050312-120057. [ Links ]
Semillas Magna. 2015. Características del híbrido de tomate 'Ichiban'. In: http://semillasmagna.com/index.php?page=shop.prod-uct_details&flypage=flypage.tpl&product_id=242&category_id=28&option=com_virtuemart&Itemid=71; consulted: January, 2015. [ Links ]
Thompson, D. 2001. Extensiometric determination of the rheological properties of the epidermis of growing tomato fruit. J. Exp. Bot. 52, 1291-1301. Doi: 10.1093/jexbot/52.359.1291. [ Links ]
Trudgill, D., A. Honek., D. Li, and N. Van Straalen. 2005. Thermal time - concepts and utility. Ann. Appl. Biol. 146, 1-14. Doi: 10.1111/j.1744-7348.2005.04088.x. [ Links ]
Zotarelli, L., J. Scholberg, M. Dukes, R. Muñoz-Carpena, and J. Icerman. 2009. Tomato yield, biomass accumulation, root distribution and irrigation water use efficiency on a sandy soil, as affected by nitrogen rate and irrigation scheduling. Agr. Water Manage. 96, 23-34. Doi: 10.1016/j.agwat.2008.06.007. [ Links ]

