











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
The growth of potato plants depends on the environmental conditions and genotype-environment interaction (Corchuelo, 2005; Cabezas, 2013), where absorption, translocation, and accumulation of essential nutrients in the tuber allow starch accumulation (Corchuelo, 2005; Kumar et al., 2013). Potato stays among the crops of the highest demand for mineral nutrients per kg of dry matter produced (Sierra et al., 2002; White et al., 2009; Kumar et al., 2013). The andigena potatoes are well adapted to soils of low fertility, but respond favorably to NPK fertilization achieving high yields (Ríos Quinchoa et al., 2010; Hernández et al., 2012). According to Grandett and Lora (1979), Villamil (2005), and Gómez and Torres (2012), the macronutrients of the highest requirement in andigena potato plants are K, N, and P followed by Mg, S, and Ca.
In potato, N is the element essential for protein synthesis, respiration, and growth of tubers (Westermann, 2005; Kavvadias et al., 2012). In soil conditions of mountain tropical regions, N availability to the plants could be affected by low mineralization rate, low temperatures, clay soils, and low organic matter content (Castro and Gómez, 2013), although the excess of N may result in luxury consumption and reduce tuberization (Ruza et al., 2013). Deficiency of N caused a reduction in dry matter, reduced leaf area and fewer leaflets that provides less light interception and a lower rate of photosynthesis (Balemi, 2009; Marouani et al., 2015; Li et al., 2016). Phosphorus promotes root growth (Aguilar-Acuña et al., 2006), rapid formation of tubers and starch synthesis (Perrenoud, 1993). In strongly acid soils with pH lower when 5.5 of the Andean regions with cultivated potato, P is fixed and forms precipitates with iron and aluminum, which decreases its availability (Hernández et al., 2012; Castro and Gómez, 2013). Potassium is essential for translocation of sugars to the tubers and starch synthesis, a fundamental processes in the tuber growth and filling (Reis and Monnerat, 2000). Low soil supply of K in sandy soils, excess of Ca (Kavvadias et al, 2012), salinity and clay minerals of 2:1 type (Castro and Gómez, 2013) limit availability of K to potato crop.
The relationship between tuber growth and demand for macronutrients NPK in andigena potatoes is scarcely documented, especially as affected by the changes in edaphic-environmental conditions of Colombian Andean region. This relationship is important to establish a specific nutrition management and efficient use of nutrients, and optimize fertilization moments in order to improve productivity and sustainability of cultivars Supreme and Capiro as economically important for fresh consumption and industrial use. The aim of the study was to evaluate the accumulation of N, P and K in two cultivars spp andigena, under different stages of tuber growth in contrasting soils of the Cundiboyacense plateau. The second objective of the research was to analyze the effect of fertilization on the growth and yield of tuber in order to establish an efficient and specific management of nutrition as dependent on soil type and plant phenology.
Materials and methods
The study was done in 2013 and 2014 in soils contrasting in the level of fertility in the Andean region of Colombia in representative locations in Cundinamarca province characterized with high yield potential (> 50 t ha-1) of potato and different environmental and soil fertility conditions (Tab. 1).
TABLE 1 Edaphoclimatic conditions in the locations of the study.
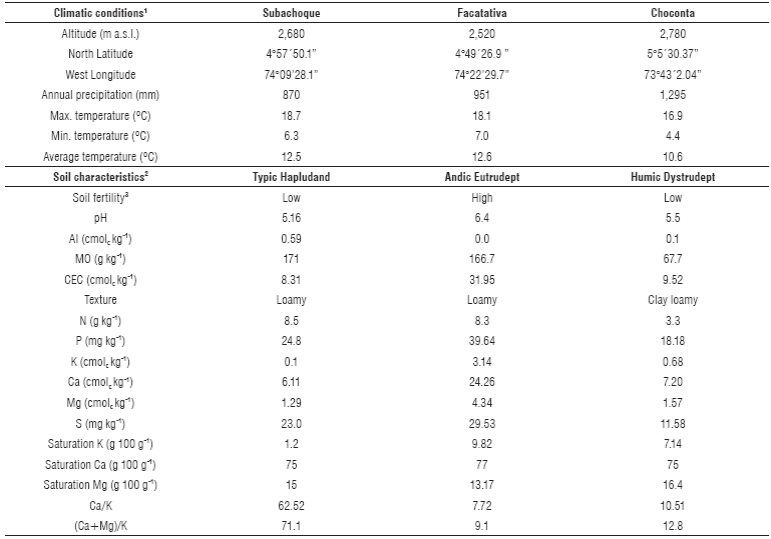
1Environmental data were taken and calculated from IDEAM (2012-2013). 2Physico-chemical characterization of soils in arable layer (0-30 cm) was done according to methodology IGAC (2006). The soils were classified according to USDA classification system (Soil Survey Staff, 2010). 3Chemical fertility potential was assessed in the topsoil 0-30 cm (Castro and Gómez, 2013).
Twelve treatments were evaluated using a repeated measures design with three factors: two cultivars ssp. andigena (Capiro, Supreme), three locations (Subachoque, Facatativa and Choconta), and two levels of fertilization, F0 (unfertilized) and F1 (fertilized). The experimental design was of incomplete nature by differential fertilization by soil type (Tab. 2), with a within-subjects factor associated with four phenological critical stages tuber growth adapted from Segura et al. (2006). These stages were stage II, 70-75 days after planting (dap) (formation of secondary stems and tuberization initiation); stage III, 90-100 dap (bloom, maximum tuberization, and beginning of tuber filling); stage IV, 120-125 dap (final flowering, tuber filling); stage V, 150-160 dap (senescence, maximum filling, and maturation of tubers).
TABLE 2 Doses of mineral nutrients applied with fertilization in the studied locations.
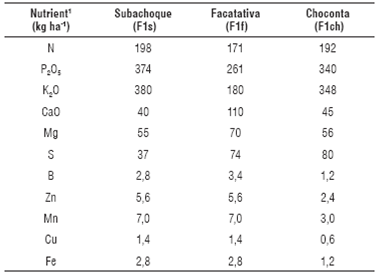
1Fertilizers applied according to the method of soil-plant balance (Castro and Gómez, 2013) and fractioned according to historical management in areas, where the high yields have been obtained (>50 t ha-1): N, 60% sowing, 40% dap 45-50; P, 70% sowing, 30% 45-50 dap; K, 30% sowing and 70% 45-50 dap. The sources of granular fertilizers were: N-P, DAP; K, KCl (0-0-60) potassium sulfate (0-0-50); Ca, calcium nitrate (25% CaO); Mg, Kieserite; Nutricomplet®, a complex micronutrient source B, Zn, Cu, Mn and Fe, based on sulfates.
In each locality, divided plot design with three replicates was used, where the main plot corresponded to the cultivars and subplots corresponded to the two levels of fertilization. The contribution of mineral nutrients, fertilizer sources and fractionation of fertilization (F1) levels in each location were done according to table 1. Planting was done in experimental units of 50 m2 (135 plants/ unit) with a row spacing of 1 m and 0.37 m between the plants, and useful harvest area of 36 m2 with a density of 27,000 plants/ha.
At each stage of tuber growth four plants were taken from each experimental unit and a destructive analysis of tubers and aerial part was carried out. At each stage, fresh weight of tubers (FWT) (kg ha-1) was measured. For dry weight of tubers (DWT), 200 g of fresh plant material were dried to constant weight in oven at 70°C for 72 h and the concentrations of N, P, and K was determined in tubers (IGAC, 2006). The methods used for elemental analysis were Micro-Kjeldahl volumetric for total nitrogen (%), calcination at 475°C colorimetric with molybdate and ammonium vanadate for total phosphorus (g kg-1), calcination at 475°C for K (g kg-1). Using the accumulation of macronutrients per dry weight values the nutritional extraction (kg ha-1) and nutrient (g kg-1) were quantified per growth stage of tubers adapted from Cabalceta et al. (2005) and Gómez and Torres (2012). With the above mentioned variables harvest index (HI) and the rate of nutrient extraction at harvest (EI) (kg of nutrient extracted per ton of harvested tuber) were evaluated.
Principal component analysis was used to perform an exploratory analysis and reduce the dimensionality of the set of variables. Two components were analyzed with a cumulative variability highly significant 87.55% as a result of statistical analysis; the weights of the components were used as new variables for which the multivariate analysis of variance, MANOVA (Ravindra and Dayanand, 1999) was performed. The Pearson correlation matrix of qualitative variables was analyzed and the regression curves were fitted; additionally, the graphs of the interaction of factors that describe the behavior and distribution of nutrients were compared. SAS statistical software version 2014 and Stat graphics version 2010 were used for the graphics.
Results and discussion
Accumulation of N, P and K in tuber phenology
At the beginning of tuberization, 75 dap, Supreme took up 51 kg N ha-1, and 100 kg K ha-1, while Capiro took up about 30 kg N ha-1 and 84 kg K ha-1, with similar uptakes of P (Fig. 1) in Humic Dystrudepts (Choconta) with increased extraction and higher yields than other soil evaluated. These significant differences between the cultivars for N y K (P<0.05) could be attributed to early and stepped tuberization in Supreme and to highly bearing phenotype of this cultivar.
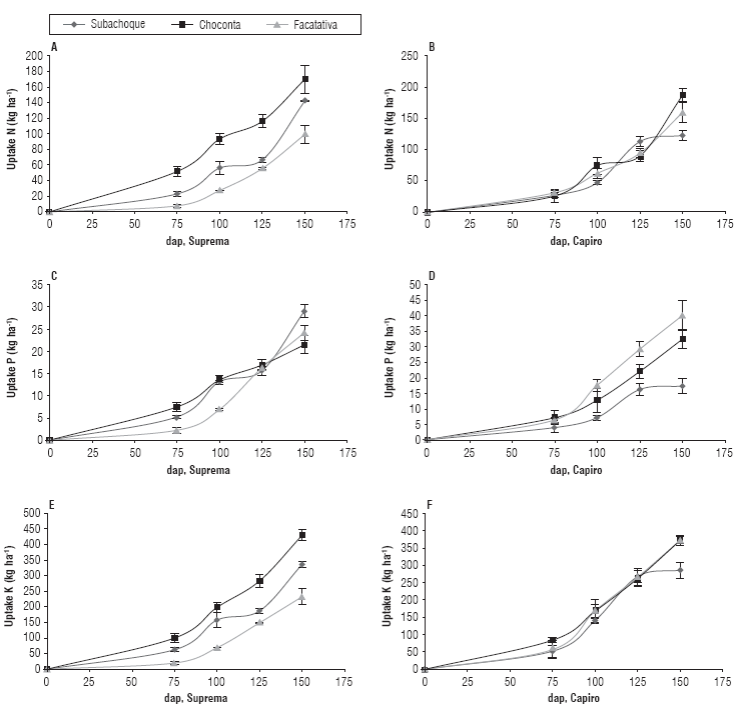
FIGURE 1 Accumulation of NPK in tubers during the growing season in andigena potato, cv. Supreme (left) and cv. Capiro (right) in Typic Hapludands (Subachoque, ♦); Humic Dystrudepts (Choconta, ■) and Andic Eutrudepts (Facatativa, ▲) with fertilization (ferl) in the altiplano Cundiboyacense. dap (day after planting). Error bars indicate standard error.
These plant features, apparently, promoted higher demand for assimilate flow into the tubers at initial growth stages in soils of low fertility, such as of Choconta. Suprema presented better uptake in soils of low fertility Humic Dystrudepts and Typic Hapludands (Fig. 1) with reduced availability of N, P, and K (Tab. 1), while accumulation of Capiro depended directly on increased availability of P in the soil, where the highest uptake of P was obtained in Andic Eutrudepts slightly acidic soil (Fig. 1, Tab. 1).
Therefore, it should be performed earlier and gradual fertilization management of Supreme as related to the fertilization of Capiro that presented accumulation of NPK 75 dap after the beginning of tuberization (Fig. 1B, D and F). After this growth stage, Capiro y Suprema increased the demand of assimilates that coincided with increased nutrient transport to tubers and a localized tuberization at this phenological stage (Figure 1). This is consistent with reports Villamil (2005) for Capiro and comparable with the data found by Sierra et al. (2002) for the cultivar Desiree in Chile and White et al. (2009) for spp tuberosum; where the authors reported increase in NPK accumulation after the beginning of the tuberization. The uptake should be synchronized with the application of NPK fertilizer to promote initial growth associated with the growth of stolons, leaves and stems, increased number of tubers and protein accumulation. In this sense, it is also important to consider factors, such as hormonal balance, low tempera-tures and photosynthetically active radiation (Ruza et al., 2013; Marouani et al., 2015).
In both cultivars, the maximum extraction of N, P, and K by the tubers presented at the tuber filling stage 150-160 dap, except cv. Capiro in Subachoque that presented a maximum extraction at 125 dap (Fig. 1B, D and F). Compared with Facatativa and Choconta, Subachoque likely had more adverse conditions in the availability of N and K (Tab. 1) and water deficit in this locality, which accelerated ripening and affected proportionately losses in yield potential because tuberization of Capiro was grouped into more defined stages 70-100 dap, with greater emphasis on nutrient availability at flowering stages. Therefore, it should be avoided stress affecting organs and metabolism sources for filling of tubers towards the end of the cycle, such as nutritional and hydric deficits, phenomena that have been reported by Monneveux et al. (2013). Equally, cv. Ica Purace presented the earliest maximum accumulation of nutrients at 120 dap (Grandett and Lora, 1979), cv. Pimpernel at 130 dap (Sierra et al., 2002) regarding accumulation at the end of the cycle for cv. Desiree (Sierra et al., 2002) and cv. Atlantic (Coraspe-León et al., 2009). The differences in nutrient accumulation were directly related to soil fertility and tuberization dynamics per cultivar, whether determinate or indeterminate, so it is important to consider these conditions in fertilizer management.
The P accumulation had significant differences (P≤0.05) between localities and cultivars mainly for Capiro, probably because of its higher nutrient requirement and lower extraction on soils poor in P (Typic Hapludands and Humic Dystrudepts) (Fig. 1D). Regarding Supreme, it presented a better response in these soils of low fertility and without differences in extraction in studied soils. Thus, Supreme showed a better adaptation of this cultivar to soils low in P and a lower requirement of P applied with fertilization and probably for its further exploration at the root and mechanisms of adaptation to acid soils by exudates of root organic acids that improve the availability of P fixed. These mechanisms of differential absorption have not yet been investigated for these cultivars.
Phosphorus was the mineral nutrient of less accumulation in tubers as compared to N and K in evaluated cultivars characterized with 17-40 kg ha-1 uptake P (Fig. 1C and D), which coincided with local study by Villamil (2005) for Capiro. The importance of P should not be underestimated, because its deficiency in potato crops dramatically reduced the size of tubers, yields (Aguilar-Acuña et al., 2006; Barben et al., 2007; Hernandez et al., 2012) and tuber quality (Fernandes et al., 2015). Phosphorus deficiency slowed down the apical growth, resulting in small plants and reduced formation of starch in tubers, which manifests itself with necrotic spots distributed in the tubers (Hernández et al., 2012). Several reports indicated that potato plants growing in soils with low P develop less leaf area, which affected light interception, reduced plant growth and differentiation, and generated less stolon bearing tubers (Barben et al., 2007). Results obtained by Balemi (2009) in three genotypes of ssp. tuberosum showed that a low supply of P affected dramatically the plant growth and production of dry matter, also the importance of P in starch accumulation in potato was comparable with that of K (Perrenoud, 1993).
Accumulation of K in tubers of Capiro was similar in three locations, with a maximum extraction of373 kg ha-1 in soils matching high availability of this element in Choconta and Facatativa (soil saturations with K >7%). Supreme presented the maximum extraction of 429 kg ha-1 in Choconta followed by Subachoque (335 kg ha-1) and Facatativa (232 kg ha-1); this cultivar evidenced a differential extraction (P≤0.05) curves according to location and that were similar to those of N (Fig. 1A and E).
The most uptake rate of K by the tubers of this cultivar was more associated with adequate environmental supply with lower temperatures <12°C and greater efficiency in the use of K y N in these soils that allows adequate sink-source management and potassium fertilization in soils of low fertility balance. In soils of high fertility, such as Andic Eutrudepts, potato plants could present luxury consumption of N y K, as it was evidenced in Facatativa, where a similarly extraction was obtained but with greater accumulation of N and K in the aerial parts and similar yields in Choconta, suggesting lower doses of these elements in these soil conditions. In addition, the higher average air temperature in Facatativa over Choconta may cause a higher photosynthetic rate that matches high growth of the aerial part and could encourage higher activity of enzymes related to assimilation of K and, therefore, a high demand for this element.
Importantly, Capiro in soils of low fertility of Subachoque with low concentrations of native soil K (0.1 cmolc kg-1) (Fig. 1F) presented an earlier accumulation of K in the tubers (125 dap) unlike 150 d in other locations (Fig. 1F). These data contrasted in this locality with an accumulation of K at the end of the cycle (150- 160 dds) for Supreme, where tuberization and tuber filling was evident by persistent vegetative development and tuber growth. This implicates a need for more precise handling of supplementary fertilization in soils of low offer of K. It could be suggested after 100 dap to keep photosynthetically active and "green" plant until the end of the cycle avoiding early ripening and to make liquid or foliar fertilization at the late stages of the crop. These technologies have been evaluated by Horvat et al. (2014) as an alternative in the supplementary fertilization management of this crop.
The steps of tuber filling and accumulation of K after 100 dap were related directly to the partition of the dry matter in tubers and were higher than the rest of the plant, which coincided with high harvest index > 75%, which favors high yield potential. Similar rates of harvest were found by Sierra et al. (2002) and Sauceda Acosta (2010) for ssp. tuberosum cv. Desireé with yields close to 90 t ha-1 and Santos et al. (2010) for cv. Capiro and cv. Suprema with yields close to 50 t ha-1. In soils Andic Eutrudepts of Facatativa, the harvest indexes (HI) for Supreme were lower than 55% and lower than those obtained in low fertility soils with HI >75%, which corroborates a low sink-source capacity in soils of high fertility associated with high availability of N and K.
Accumulation of N, P, and K among cultivars
The demand for N, P, and K in the cultivars was directly related to the accumulation of FWT and DWT. A sequence of accumulation of mineral nutrients K> N> P in both cultivars was observed, similar to that reported by Westermann (2005); Coraspe-León et al. (2009); Subramanian et al. (2011) and Fallas and Bertsch (2014) in ssp. tuberosum, and by Grandett and Lora (1979), Villamil (2005); Gomez and Torres (2012) and Lefévre et al. (2012) in andigena potato.
The significant relationship between nutrient extraction and FWT allowed estimating the harvest extraction index (HEI) in kg of nutrient extracted per ton of harvestable organ (Fig. 2), and is important to determine nutrient requirements in fertilizer recommendations according to performance targets locals as reported Ciampitti and García (2008) and Fallas and Bertsch (2014). HEI of N and P showed significant differences among cultivars with higher yields at 50 t ha-1 (P<0.05) and HEI higher for Supreme than for Capiro in a positive linear model (Fig. 2A and B). For Supreme HEI was 1.92 kg N t-1, 0.38 kg P t-1; 5.3 kg K t-1, while for Capiro HEI was 2.27 kg N t-1, 0.47 kg P t-1; 5.3 kg K t-1, when compared with results reported by Sierra et al. (2002) and Ciampitti and Garcia (2008), when in potato ssp. tuberosum HEI was 3.5 kg N t-1, 0.7 kg P t-1; 5.4 kg K t-1 with greater differences for N and P and equal for K.
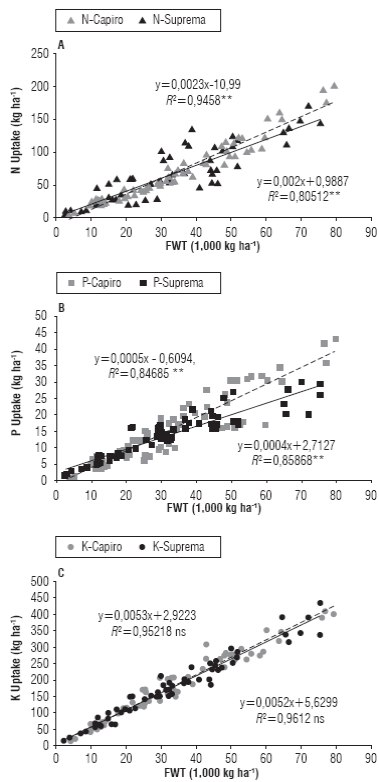
FIGURE 2 Relationship between the uptake in tubers of N (A), P (B) and K (C) (kg ha-1) and yield (FWT) in cv. Capiro and cv. Suprema in soils of Cundiboyacense plateau. ** The value of coefficients b were significant at the 0.05 probability level. ns, not significant the value of coefficients b.
Capiro required 32% more of N and 48% more of P for the yields superior than 50 t ha-1 more than Supreme with significant differences in beta coefficient according to a positive lineal model in both cultivars (Fig. 2A and B). These variations in N and P uptake in Capiro can be explained by a genotype of low stature and tuberization more synchronized with better adaptation to fertile soils and highest genetic potential; in addition this was also associated with accumulation of organic and mineral forms that determine the nutritional characteristics of the tubers. This confirms the differences in accumulation of NPK between potato genotypes reported by White et al. (2009), Karam et al. (2009) and Wekesa et al. (2014). It coincides with the differences in accumulation of these nutrients in cultivars ssp. tuberosum made by Tekalign and Hammes (2005) and White et al. (2009) in identical environments and andigena cultivars in the Bogota Plateau made by Grandett and Lora (1979).
Significant differences (P<0.05) in extracting P in favor of cultivar Capiro regarding Supreme (Fig. 1B and 2B) can be explained by the most concentrated tuberization, the better translocation of assimilates and increased demand of sinks than that in Supreme. This features in Capiro facilitate uniform ripening with a defined harvest, which requires a greater demand for ATP and may be associated with organic and inorganic forms of P. These forms of P could serve as a reserve and will explain responses in the dormancy period of the tubers that lasts up to 4 months in this cultivar and is longer than that in Supreme that presented an earlier period of up to two months.
The differences in P suggest a higher concentration of phytate complex for Capiro than that in Suprema, which should be evaluated in future studies to test this hypothesis. In relation to P in the tubers Subramanian et al. (2011) mentioned that this element was metabolized from organic bonds orthophosphate-ester in starch that accumulate early in tuber formation and from mineral forms of complexes of phytic acid (myo-inositol hexakisphosphate) towards maturation of tuber and sprouting and is necessary for proper formation of seeding tubers. The latter complex may reach between 0.1 and 0.27% DWT with differences between cultivars ssp. tuberosum according to Phillippy et al. (2004).
HEI for K showed no significant differences between the cultivars and this was the element that had the highest correlation (r 2 >0.95) with both yield and DWT compared to N and P (Figure 2c ). According to the results for K, HEI presented independent effects between cultivar, locality and fertilization, suggesting that the K extraction in these cultivars of ssp. andigena was mainly depended on the genetic potential of the cultivars. The later corroborate the direct effect of K in transport, storage, and conversion of carbohydrates in ssp. andigena, therefore, the importance of studied cultivars for the process of industrial transformation whit high dry matter in tuber that was corroborated in this study.
The K compared to N and P is the nutrient that had the highest correlation in Capiro (r2 0.95) and Supreme (r2 0.99) respect to DWT. The relationship between K and tuber growth matched the one reported by Coraspe-León et al. (2009), Karam et al. (2009), Sarkar et al. (2010), Kavvadias et al. (2012) for potato ssp. tuberosum and Villamil (2005) and Pérez (2015) for potato ssp. andigena. The accumulation of K was directly related to accumulation of dry matter (% DWT) and starch in the tubers (Perrenoud, 1993; Westerman et al., 1994; Reis Jr and Monnerat, 2000) of 19-28% DWT for ssp. andigena (Jiménez et al., 2009). DWT % evaluated in this study (20-27%) was similar to those reported by Jiménez et al. (2009) and Ñústez (2011).
Conclusions
It is necessary a differential management of mineral nutrition in cultivars according to their phenology and soil type and considering the differences in nutritional extractions with higher yields to 40 t ha-1 in both cultivar. It is suggested the management Supreme fertilization in soils of low fertility applying 30% of total extraction NPK before stage II, beginning of tuberization (75 dap) in a uptake ratio N:P:K of 10-2-12 N as related to Capiro. Capiro requires about 15% of the total extraction NPK in the uptake ratio N: P: K 10 2-18; which shows a higher uptake ratio of K in early stages, so its best response was in soils Humic Dystrudepts of Chocontá and Andic Eutrudepts of Facatativá where saturation K soil was >7%. For tuber filling stages, before stage III (maximum tuberization and start filling), 70% (Supreme) and 85% (Capiro) of the total NPK required by the tuber with an 8-1-20 uptake ratio for Supreme and 12-2-25 uptake ratio for Capiro should be applied. These recommendations can improve the physiological efficiency of these nutrients by taking into account the timing be-tween doses, ratio, and age. Actually, it is fractionated in two applications before 50 dap and is not applied according to the nutrient extraction.

