











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
The Cucurbitaceae family is composed of approximately 118 genera and 825 species. One of its most important genus is Cucurbita, which is composed of 20 to27 species according to Esquinas and Gulick (1983), 15 species according to Lira (1995), and 12 to 13 species according to Nee (1990).
Five species of the genus Cucurbitaare were domesticated in the New World and have been grown for centuries (Merrick, 1990; Smartt and Simmonds, 1995; Valdés, 2014): C. argyrosperma Huber, C. ficifolia Bouché, C. maxima Duchesne, C. pepo L., and C. moschata (Duchesne ex lam) Duchesne ex Poir.
Nee (1990) and Lira (1995) suggested that the species C. argyrosperma is constituted by two subspecies: argyrosperma and sororia. The subspecies argyrosperma is formed by four varieties: argyrosperma, callicarpa, stenosperma, and palmieri. The first three varieties include all cultivated types, while the fourth corresponds to related wild populations. The subspecies sororia L.H. Bailey comprises wild populations distributed extensively from Mexico to Nicaragua. Whitaker (1981) and Lira (1995) suggested that the subspecies sororia is the wild ancestor of the domesticated species based on both its reproductive compatibility and its morphological similarity. Its scarce diffusion worldwide is probably attributable to its poor pulp quality as compared to C. moschata or C. pepo. Different parts of the plant (flowers, young stems, tender or mature fruits) are consumed as vegetables (Lira, 1995), and its seeds are consumed whole, roasted, toasted, or ground, being the main ingredient in an array of sauces (Valdés et al., 2010). Due to their high oil (39%) and protein (44%) contents, seeds are the most important product of the fruit (Ortiz et al., 2009; Nawirskaet al., 2013; Valdés, 2014).
In Colombia, a native or creole butternut squash variety, C. moschata, produces, on average, 400 kgha1 of seed, and an average yield of 200 Lha-1ether extract. This variety may be consumed as food or destined to medicinal and agroindustrial uses (Ortiz et al., 2009). Its oil is characterized by a high unsaturated fatty acids content (55.28%), in particular linoleic acid (55.11%). Oilseed cake contains approximately 51.11% protein and 4604.66 kcal kg-1 energy (Ortiz et al., 2009).
In Greece, the oil obtained from C. pepo is well-known as an alternative source of biodiesel (Schinas et al., 2009). The seed of this species has a high oil content and is rich in polyunsaturated fatty acids, such as linoleic (43-56%) and oleic (24-38%), beta and gamma tocopherols (vitamin E), and carotenoids such as luteolin and beta-carotene (López et al., 2009).
The seed oil of C. maxima has antioxidant properties (Nawirska-Olszańska et al., 2013) and it is well known for its multiple health benefits such as the prevention of prostate growth, reduction of hypertension, mitigation of hypercholesterolemia and arthritis, reduction of gastric levels, and prevention of cancer.
Most of the studies conducted on Cucurbita seed have focused on oil content, fatty acid profile and intrinsic seed properties. To date no studies have been reported on the heterosis for ether extract production in seeds of Cucurbita argyrosperma subsp. sororia.
This study aimed to evaluate the average, variety, and specific heterotic effects related to the character of ether extract production per plant and its components in C. argyrosperma subsp. sororia.
Materials and methods
Study area
The Fieldwork was carried out at the Experimental Centre of the Universidad Nacional de Colombia (UNAL, as its Spanish acronym), located in the municipality of Candelaria, Valle del Cauca, Colombia (2°6' N and 63°3' W) (Espitia et al., 2006). The experimental Centre, located at 972 m above sea level, presents an average annual temperature of 26°C, average annual precipitation of 1100 mm, and 76% relative humidity. Based on rainfall and sunshine rates, the climate is classified as sub-humid (Valdés et al., 2010). The macromolecular evaluation was carried out at the Seeds Laboratory of UNAL-Palmira Campus.
Genetic material
Twenty-one genotypes of C. argyrosperma subsp. sororia, obtained from diallel crosses between six selected parental lines (introductions 256, 140, 260, and 132 from Central America and 107 and 68 from Mexico) and their corresponding 15 direct F1 crosses, were evaluated (Fig. 1, 2, 3, 4, 5, and 6). A randomized complete block design was used with four replicates and five plants per replicate, and the three central plants of each plot were evaluated. A planting distance of 3 m between and within the furrow was used. The diallel cross was evaluated in two consecutive semesters: 2012B and 2013A.



FIGURE 3 Introduction 132 of Cucurbita argyrosperma subsp. sororia. A, whole fruit; B, fruit cut open at the equatorial diameter.

FIGURE 4 Introduction 260 of Cucurbita argyrosperma subsp. sororia. A, whole fruit; B, fruit cut open at the equatorial diameter.

FIGURE 5 Introduction 107 of Cucurbita argyrosperma subsp. sororia. A, whole fruit; B, fruit cut open at the equatorial diameter.

Evaluated features
Ether extract production per plant (EEPP): the number of fruits per plant (NFP) multiplied by seed weight per fruit (SWF) and percentage ether extract (EE) in seed, expressed in g.
Ether extract (EE) in seed: ether extract content (%) in seed.
Seed weight per fruit (SWF): average weight (g) of three fruits randomly taken from each genotype and conditioned at 12% moisture.
100-seed weight (100-SW): weight (g) of 100 seeds randomly taken from three fruits of each genotype and conditioned at 12% moisture.
Number of fruits per plant (NFP): average number of fruits per plant based on three plants of each genotype.
Heterosis analysis
The fixed model (selected parental lines) (Griffing, 1956) and method 2 (parental lines and direct hybrids) of the methodology proposed by Gardner and Eberhart (1966) was used. The statistical genetic model is the following:
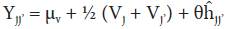
Where:
Yjj.: expected average value of a parental variety (j = j') or hybrid (j ≠j'). If j = j', then θ = 0 and if j ≠ j, then θ = 1
μv: mean of n parental varieties.
Vj: variety effect of the j-th parental variety.
ĥjj’: effect of heterosis that results when variety j is crossed with variety j’
Total heterosis (ĥjj’) break down as follows: ĥjj’ = + (hj + hj’)+ Sjj’
Where:
ĥ: mean heterosis of all hybrids.
hj and hj’: variety heterosis produced by the j-th variety.
Sjj’: specific heterosis for the cross between varieties j and j’; this is a deviation from the expected mean based on the effects ĥ + hj.
The analysis ofvariance proposed by Gardner and Eberhart (1966) included the main sources of variation: replicates, genotypes (parental lines and hybrids), error, and total. The source of variation "genotypes" was broken down into effects of varieties (Vj) (parental lines), and heterosis (hjj'). On the other hand, total heterosis effect was broken down into average heterosis (h), variety heterosis (hj), and specific heterosis (Sjj').
Average heterosis (ĥ) is the difference between the average of all hybrids and the average of all parental lines.
Variety heterosis (hj) is the difference between the heterosis given by parental lines or variety j in its corresponding hybrids and the average heterosis of all the parental lines.
Specific heterosis (Sjj) between parental lines or varieties j and j' measures the deviation between the observed performance of a specific hybrid and its expected performance, based on variety effect (Vj), average heterosis (ĥ), and variety heterosis (hj) (Espitiaet al., 2006).
Software developed by SAS Institute Inc. (2000), SAS-9.1 (Windows version), and Microsoft Excel 2010 were used to process results.
Results and discussion
Analysis of averages
In both semesters, prevailing environmental conditions affected the performance of parental lines and hybrids. In semester 2012B, evaluated features presented higher averages due to the semester precipitation and temperature and how they influenced the phenotypic expression. In 2013A, the excess precipitation negatively affected the evaluated features.
Tab. 1 shows the parental lines and hybrids averages regarding EEPP and its components on C. argyrosperma subsp. sororia. The average of both parental lines and hybrids in 2012B was higher than that of 2013A to all the evaluated features, except to the character EE for which parental lines presented a lower average in 2012B.
TABLE 1 Averages of parental lines and hybrids for the character of ether extract production per plant (EEPP) and different components in Cucurbita argyrosperma subsp. sororia for two semesters (2012B and 2013A).
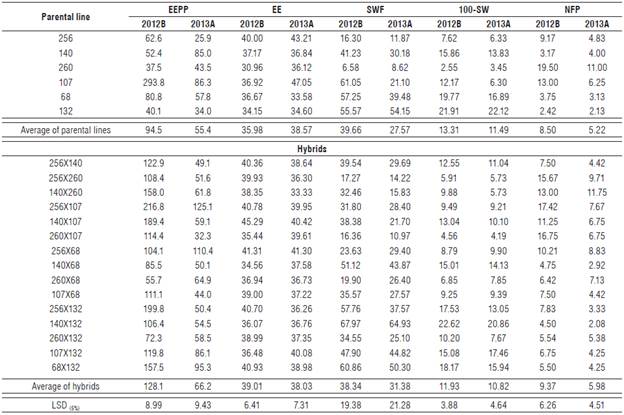
For the feature EEPP, the average of hybrids was superior to that of parental lines in both semesters, indicating high genetic diversity. However, no hybrid exceeded the best parental line 107, which presented 293.8 g/plant in 2012B and 86.3 g/plant in 2013A. Only hybrid 256x107 yielded a similar value: 216.8 g/plant in 2012B and 125.1 g/plant in 2013A.
For the components of the feature EEPP (EE, SWF, 100-SW, and NFP), the performance of hybrids was similar to that of parental lines in both semesters evaluated, indicating low genetic diversity for these components.
Table 2 presents the mean squares of the analysis of variance for the character EEPP and its components in semesters 2012B and 2013A.
TABLE 2 Square means of variance analysis of ether extract production per plant (EEPP) and its components according to the methodology of Gardner and Eberhart (1966) in a diallel cross, evaluated in two semesters (2012B and 2013A) in Cucurbita argyrosperma subsp. sororia.
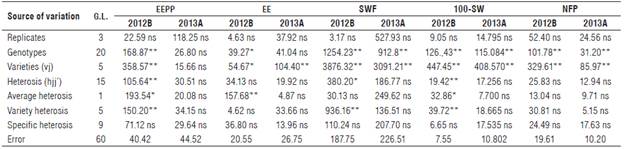
* Significant effect (Pr>F)<0.05; ** Highly significant effect (Pr>F)<0.01; ns: not significant.
For the character EEPP, the average squares associated with varieties (six parental lines) and heterosis (15 hybrids) for semester 2012B were highly significant, confirming the existence of genetic diversity between parental lines and between hybrids for this feature. There were no significant differences in semester 2013A, possibly because its genetic expression was affected by unfavorable environmental conditions. These data agree with the environmental effects found in other experiments on the agroindustrial traits of C. moschata Duch. (Valdes et al., 2010; Valdes et al., 2013).
The average heterosis of the character EEPP and two of its components, EE and 100-SW, was significant in semester 2012B, indicating that genetic variation exists between parental lines to explore heterosis. For the remaining components (SWF, 100-SW, and NFP), average heterosis was non-significant, indicating absence of genetic diversity between parental lines to explore heterosis.
Variety heterosis was highly significant for the character EEPP and two of its components, SWF and 100-SW, only in semester 2012B, confirming the presence of genetic diversity between the parental lines and these features. Variety heterosis was non-significant for components EE and NFP in both semesters, showing little genetic divergence between parental lines.
Specific heterosis was non-significant to the character EEPP and its components in both semesters, indicating that parental lines did not present favorable gene complements in the different hybrids. Similar results were observed on the species Cucurbita moschata (Valdés et al., 2014).
Estimating heterosis effects on C. argyrosperma subsp. sororia
Effect of average heterosis
Table 3 shows the effect of average heterosis on the character EEPP and its components. Average heterosis was best expressed in 2012B, where there were significant differences for EEPP and the component EE. In 2013A, only the component SWF presented significant differences. The significance of these characters indicated that, overall, the parental lines used are suitable to explore the effect of heterosis on hybrids regarding the aforementioned features. Regarding the other characters (100-SW and NFP), the parental lines used are not suitable to tapping heterosis.
TABLE 3 Effect of average heterosis of the feature ether extract production per plant (EEPP) and its components during two semesters (2012B and 2013A) to the species Cucurbita argyrosperma subsp. sororia.
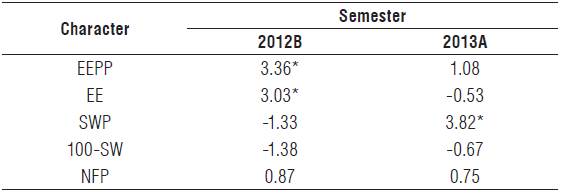
* Significant effect (Pr>F)<0.05
Effect of variety heterosis
Table 4 shows the effects of variety heterosis for all evaluated characters.
TABLE 4 Effects of variety heterosis of the character ether extract production per plant and its components during two semesters (2012B and 2013A) for Cucurbita argyrosperma subsp. sororia.
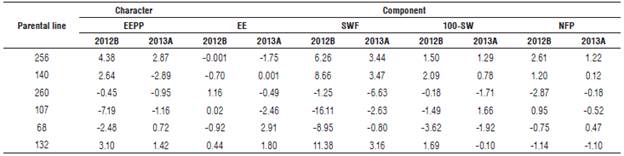
Variety effects varied between semesters. However, parental line 256 presented positive effects in both semesters to the character EEPP and its components SWF, 100-SW, and NFP, indicating that it is a stable parental line and can be used in recurrent selection programs to multiply these characters.
Parental line 132 presented positive effects for both semesters to the character EEPP and its components EE and SWF, indicating that this parental line also has important additive genetic effects that can be used in selection programs to multiply these characteristics.
If the objective of a breeding program is to increase the percentage of ether extract on seed and, at the same time, ether extract production per plant, parental lines 256 and 132 should be considered for hybridization and selection programs.
Effect of specific heterosis
Specific heterosis helps identify hybrids with better characteristics than those of their parental lines. The significance of this heterosis indicates presence of dominance effects, probably attributable to favorable gene complementation in the crosses in which they form part.
Table 5 shows the effects of specific heterosis, which varied between semesters. For the character EEPP, the effects of hybrids 256x132 and 140x260 were significant for semester 2012B, indicating that these hybrids were superior to the average of their parental lines and the overall average. In semester 2013A, no significant values were obtained.
TABLE 5 Effects of specific heterosis of the character ether extract production per plant and its components during two semesters (2012B and 2013A) for Cucurbita argyrosperma subsp. sororia.
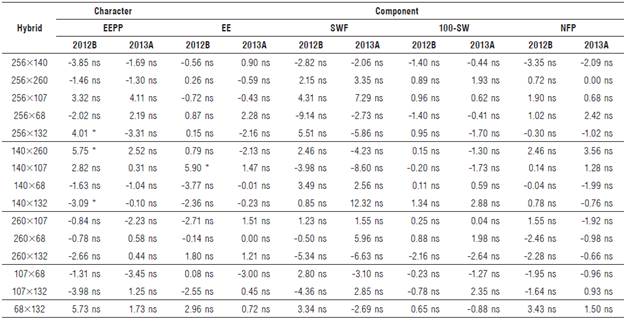
Hybrid 140x107 was outstanding in terms of percentage of EE in semester 2012B, presenting positive, significant values. In semester 2013A, although there were hybrids presenting positive values, these were non-significant, suggesting a reduced variability among parental lines and possibly a marked environmental effect even on hybrids.
It is therefore recommended to conduct studies on the response of parental lines and hybrids in additional contrasting environments.
The effects of specific heterosis to components SWF, 100-SW, and NFP presented positive but non-significant values, indicating that no genetic complementation exists between their parental lines, even though their geographic origin differs.
Conclusions
The group of parental lines used allowed the effect of average heterosis to be explored to the character EEPP and its EE components.
Parental lines 256 and 132 proved best to tap heterotic effects, based on the effect of average and variety heterosis. Furthermore, of all the parental lines evaluated, their stability was the highest to begin a crossbreeding program to select superior-performing segregants to the character EEPP.
The average heterosis of hybrids 256x132 and 140x260 was superior to that of their parental lines and the overall average to the character EEPP in 2012B. Hybrid 140x107 was outstanding regarding the EE component, also during 2012B.
In the case of semester 2013A, although several hybrids presented positive values, these were non-significant.

