











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
Light wavelengths between 300 and 900 nm can affect the growth and development of plants. However, the quality of light is not the only factor that can influence the growth processes of plants because other properties, such as light intensity and duration as well as climatic factors, are equally involved. The research using a single wavelength range is very useful in identifying the spectrum most suitable for the agricultural application (Casierra-Posada and Peña-Olmos, 2015).
The study of the response of plants to light quality has aroused the interest of researchers in order to improve production (Casierra-Posada et al., 2015). Among the bands of the visible spectrum, the green and infrared ranges are of particular interest because they are poorly absorbed or not absorbed; nonetheless, they induce important photomorphogenic responses (Wang and Folta, 2013). Although it is presumed that the amount of light being absorbed in the green range is very low and, therefore, not important for photosynthesis, the response spectrum used for photosynthesis shows that green light is in an effective spectrum band for increasing plant photosynthesis (Kim et al., 2004). On the other hand, CAM-type species can growth in contrasting environments in terms of light intensity. However, it is still unknown how these species can meet energy and carbon requirements when exposed to shadow conditions (Ceusters et al., 2011).
Sisal is a CAM-type plant grown in Colombia to produce natural fibers (Casierra-Posada and Gonzalez, 2009); therefore, because of its physiology, this species is of great interest, especially for the study of its adaptation to contrasting conditions of light intensity and quality. Based on previous approaches, this study was developed to evaluate the effect of shading and filtered sunlight through green polypropylene films on the growth of sisal plants (Furcraea hexapetala Jacq.).
Materials and methods
This experiment was carried out under greenhouse conditions in Tunja, Colombia, at the facilities of the Universidad Pedagógica y Tecnológica de Colombia (UPTC) located at an altitude of 2,690 m a.s.l. at 5°33'10.62" N and 73°21'23.97" W. During the study, an average temperature of 19.0°C, 72.6% relative air humidity and 606.0 ± 362.9 цггю1 m-2 s-1 illumination were maintained in the greenhouse. The illumination was natural, with a photoperiod of ± 12 h of light / 12 h of darkness, corresponding to tropical conditions.The plants were obtained from(Furcraea hexapetala Jacq., plant known as Fique or Cabuya) bulbs, which were placed under hydroponic conditions in 4 L containers filled with the following solution: (mg L-1): Nitrogen, 200; Phosphorus, 100; Potassium, 50; Calcium, 2; Magnesium, 10; Sulfur, 15; Iron, 0.2; Manganese, 1; Copper, 3; Zinc, 6; Boron, 4; Molybdenum, 0.1 and Cobalt, 0.05. The pH of the solution was adjusted to 6.2. The solution in each container was aerated with an aquarium pump to provide oxygen to the solution.
One month after planting, when leaf emergence was observed, the plants were divided into four groups consisting of ten plants each. Three groups of plants were exposed to sunlight filtered through green polypropylene films with a thickness of 15 μηι, placed 1 m above the plants and mounted on wooden frames that covered all sides of each group of plants, except underneath. The frame of the first group was covered with a polypropylene filter, the second with two and the last with three. The remaining group of plants was left exposed to the environmental conditions inside the greenhouse without any cover and was used as a control treatment. As a consequence of placing the polypropylene filters on the frames, the illumination in each group of plants was reduced by 55.4, 85.8 and 90.1%, respectively from 606.0 ± 362.9 μmol m-2 s-1 (Tab. 1).
TABLE 1 Environmental conditions in the sisal (Furcraea hexapetala Jacq.) plants exposed to shading with green-colored covers.
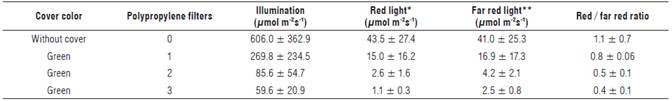
The illumination measurements were taken under the polypropylene filters, 10 cm over each plant.
* Red light was measured at 660 nm, 40 nm FWHM (Full width at half maximum) (± 20 nm).
** Far red light was measured at 730 nm, 30 nm FWHM (Full width at half maximum) (± 15 nm).
The trial lasted 256 d counted from the moment of sowing, at the end of which the plants were collected for destructive tests and final measurements. With the information collected at the end of the trial, the absolute growth rate (AGR), the relative growth rate (RGR), the root to shoot ratio (based on dry weight), the leaf area ratio (LAR), the specific leaf area (SLA), and the leaf weight ratio (LWR) were determined according to Hunt (1990). The total dry weight per plant was recorded by drying the plants in an oven at 80°C until reaching a constant dry weight. The dry matter partitioning was taken as the percentage of dry weight of leaves, stem and roots, separately, in each plant, depending on the total dry weight in the plant. Twice a week, the amount of water missing in each container was recorded and taken as the water uptake. The water use efficiency (WUE) was calculated as the dry mass quantity produced by the whole plant as a function of the amount of water consumed, according to Sinclair et al. (1984). The chlorophyll concentration index (CCI) was determined on all leaves of each plant and the average value per plant was taken using a CCM 200 Plus meter (Opti-Sciences, Hudson, USA). The leaf area was determined using a Li-cor 3000A analyzer (LI-COR Biosciences, USA). The illumination and red/far-red ratio were measured with a Field Scout Quantum Meter and Field Scout Red/Far Red Meter (Spectrum, Plainfield IL, USA), respectively. Additionally, the light transmitted through one, two and three polypropylene filters in the wavelength range of 300-880 nm was determined using a Shimadzu UV-1603 spectrophotometer (Tokyo, Japan).
The experiment was arranged in a totally-randomized one-factor design. Each treatment consisted of 10 plants. The measurements were taken individually on each plant. The collected data were subjected to an analysis of variance (P<0.05) and Tukey's significance test using the statistical software IBM-SPSS Statistics version 20.0.0 (IBM Corporation, New York, USA).
Results and discussion
Quality and decrease of the incident light
The transmittance of light through the green filters presented the behaviour shown in Figure 1. The maximum transmittance occurred at 533 nm; however, the transmittance was reduced as more polypropylene filters were used. Therefore, it was shown that the plants were exposed to a spectrum enriched in the green range, but also that the incident radiation on the plants was enriched in light in the far red range, as the plants were grown under the influence of more polypropylene filters, as shown in Tab. 1 and Figure 1. In this respect, light in the green range (510-550 nm) was considered to be an inactive portion of the solar radiation since it was thought to be reflected by the leaf blades and not used as an energy source. However, there has been increasing interest in the study of the effect of this spectrum range on the behaviour and life cycle of plants (Efimova et al., 2017) and plant responses to green light exposure, which may be dependent or independent of the action of light receptors in the blue range, involving cryptochrome (CRY) activity. CRYs act, functionally and physically, together with phytochromes (PHYs) in the regulation of physiological processes in plants, without excluding other photoreceptors in the perception of light in the green range nor the interaction of several photoreceptors (Dhingra et al., 2006).
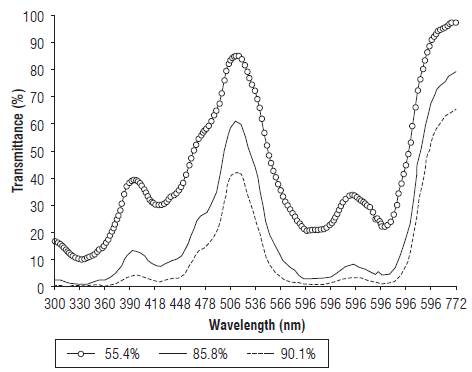
FIGURE 1 Transmittance of light filtered through one, two, and three green polypropylene films, which caused a light decrease of 55.4, 85.8 and 90.1%, respectively.
On the other hand, light in the far red range has a strong effect on plants. For example, the results reported by Turk et al. (2003) who found that light in the red, blue, and white ranges suppresses CYP72B1 transcript accumulation in Arabidopsis, but increases with the exposure of plants to light in the far red range, given that genetic and physiological effects of hypocotyl response to exogenous blue light and varying intensities of white and monochromatic light revealed that CYP72B1 modulates photomorphogenesis, mainly with exposure to the far red band and, to a lesser extent, with exposure to blue and red light. Therefore, it is possible to infer that the responses of the sisal plants in the present study were the result of the joint action of light in the green and far red bands and the shading.
Kim et al. (2004), based on the approaches of several studies, stated that light in the green band is not important for photosynthesis since it has a low absorption coefficient in the absorption ranges of chlorophyll. However, although the amount of light absorbed in the green range is very small, unabsorbed wavelengths are repeatedly reflected from one chloroplast to another in the complex network of photosynthetic cells. With each reflection, a percentage of these wavelengths is absorbed, until finally half or more end up being absorbed by most leaves and used in photosynthesis, since it has been found that the response spectrum used for photosynthesis shows that green light is in an effective spectral band to boost photosynthesis in plants. This process involves some auxiliary pigments and the transfer of energy to the reaction centers. In leaves, the absorption of light by carotenoids shifts from the blue portion of the spectrum towards the green, also used in photosynthesis, since most carotenoids (both β-carotene and xanthophylls) present in the tilakoids efficiently transfer their excitation energy to the same reaction centers as that of chlorophyll, also contributing to photosynthesis.
Chlorophyll content
The chlorophyll content index (CCI) values were significantly reduced with exposure to shading, with statistically significant differences only between the control and the set of shaded plants with the green filters (Tab. 2). This reduction in the plants exposed to shading was in the range of 52.2-55.0% in relation to the control plants. In this respect, the shading caused alterations in the plant pigment content, given that Casierra-Posada et al. (2012b) found that, in Calendula officinalis plants, the value of the a/b chlorophyll ratio was higher in leaves of the shaded plants, while the value of carotene/chlorophyll ratio was higher in plants that grew with full exposure, implying that shading increased the content of chlorophyll, relative to that of the carotenes. Additionally, in Beta vulgaris plants, Casierra-Posada et al. (2015) found an increase in the value of the chlorophyll a/b ratio and a decrease in the carotene/chlorophyll ratio, in plants placed under a green cover, in relation to plants placed under transparent cover. However, the a/b chlorophyll ratio showed a similar behaviour in the plants exposed to the green cover as in the red and blue color coverings. For the carotene/chlorophyll ratio, there was no statistical difference between the values found under the green cover and those found under the blue and yellow covers.
Additionally, Materová et al. (2017), in Hordeum vulgare plants exposed to green monochromatic light, found an increased amount of geranylgeranyled chlorophyll, as compared to plants exposed to white light, as a consequence of an altered activity of the enzyme geranylgeranyl reductase, responsible for the reduction of phytyl chain double bonds in chlorophyll synthesis, indicating that chlorophyll synthesis is severely affected as a result of exposure to green light. Therefore, it can be inferred that the values found for CCI in the present study were due to the combined action of shading and green color of the cover since both have a marked influence on the content of photosynthetic pigments.
Growth variables
We found a significant difference in the absolute growth rate (AGR) values between the control treatment and the group of shaded plants with the green cover, regardless of the level of shading. The AGR was reduced by between 61.3 and 83.0% in the plants placed under the green covers relative to the control plants (Tab. 2). Casierra-Posada et al. (2012a) reported an 85.6% decrease in the AGR value in Fragaria sp plants exposed to green polypropylene covers, as compared to plants grown under transparent covers. On the other hand, a reduction of35.1, 44.0 and 58.4%, relative to the control plants, was observed in the relative growth rate (RGR) values in plants exposed to 55.4, 85.8 and 90.1% shading using green covers, respectively, with statistically significant differences between all treatments (Fig. 2). In this regard, Casierra-Posada et al. (2014b) found no difference in the RGR value in Beta vulgaris plants exposed to green and transparent polypropylene covers, whereas Casierra-Posada et al. (2012a) found a 47.3% decrease in the RGR value in Fragaria sp plants placed under green covers, as compared to plants developed under transparent covers. For RGR and AGR values, these authors suggested that, when the quality and intensity of light on plants is evaluated, the degree of illumination is more determinant than the spectral composition and the response of plants in terms of growth since acclimatization to the environmental supply depends on the amount of light rather than its quality. Additionally, Casierra-Posada et al. (2014b) stated that certain spectral wavelength bands that are not useful for plant metabolism can significantly influence the morphology, composition and adaptive strategy of a plant, optimizing light capture when the light quantity or quality is unfavourable.
TABLE 2 Variables evaluated to determine the growth of the sisal plants (Furcraea hexapetala Jacq.) exposed to shading using green polypropylene filters.
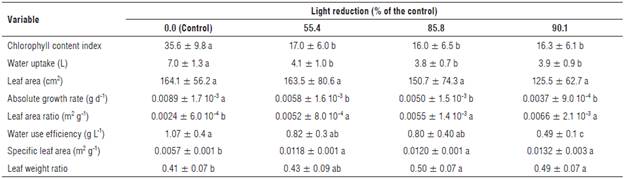
Mean values are shown followed by the standard deviation. For each variable, different letters indicate statistical significance according to Tukey's honest significance test (P≤0.05).
Casierra-Posada, Portilla-Fuentes and Molano-Díaz: Combined effect of green-colored covers and shading on the growth of sisal (Furcraeahexapetala) plants
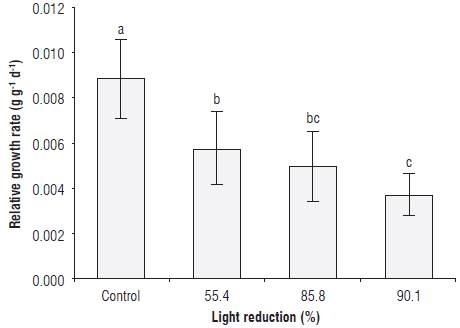
FIGURE 2 Relative growth rate in sisal plants (Furcraea hexapetala Jacq.) exposed to shading with green polypropylene filters. The bars in each column indicate the standard deviation. Different letters indicate statistical significance according to Tukey's honest significance test (P≤0.05).
As for the specific leaf area (SLA) values, an increase in the shaded plants with the green covers was evidenced in relation to the control plants, with significant differences. The group of treatments with plants that were exposed to shade using green covers, without taking into consideration the level of light reduction, showed an increase of 107.5132.4% in the SLA value as compared to the control plants (Tab. 2). In agreement with these results, Casierra-Posada et al. (2012a) reported an increase in the value of this variable of 97.5% in Fragaria sp plants maintained under green polypropylene covers, as compared to plants that grew under a transparent cover. On the other hand, Kim et al. (2004) found a SLA value of 49.4 m2 kg1 in Lactuca sativa plants exposed to green fluorescent lamps, whereas in plants placed under cool-white fluorescent lamps, the value of this variable was reduced to 33.9 m2 kg-1. In this regard, these authors mentioned that a larger leaf area per unit of dry leaf mass obtained in plants placed under green light is indicative of greater photosynthetic surface area per unit of leaf tissue inversion, which is associated with the behaviour of plants exposed to shading. Therefore, they inferred that, in the plants exposed to light in the green band, a great amount of resources was assigned with the purpose of obtaining a greater leaf area, whereas the capacity of assimilation of carbon in relation to the leaf area was reduced, as compared to other treatments. In addition, changes in the leaf structure of plants exposed to different spectral environments could also have a significant impact on the chloroplast density in leaves, as well as the amount of light captured by the photosynthetic pigments.
The values found for the leaf area ratio (LAR) showed a statistically significant difference between the control treatment plants and the set of plants that grew exposed to the green filters. In the set of plants that grew under the green filters, regardless of the intensity of light reduction, the LAR value increased between 116.6 and 174.9% in relation to the control plants (Tab. 2). In agreement with this result, Casierra-Posada et al. (2012a) reported a 320.6% increase in the LAR value in Fragaria sp plants exposed to a green polypropylene cover, as compared to plants exposed to a transparent cover. These authors mentioned this result as an adaptation of the plants to the shade induced by the green cover; therefore, plants exposed to conditions of shading and green light allocate more resources for the formation of a greater assimilation surface to overcome the stress induced by the joint action of the shading and the color of the cover.
The leaf weight ratio (LWR) values increased 4.2, 20.3 and 19.8% in the plants exposed to 55.4, 85.8 and 90.1°% shading with green covers, respectively, in relation to the control plants, with statistically significant differences between all treatments (Tab. 2). Casierrra-Posada et al., (2012a) found an increase of 113.3% in the value of this variable in Fragaria sp plants exposed to a green polypropylene cover in relation to plants placed under a transparent cover. In this regard, these authors mentioned that many plant species show an increase in the leaf weight ratio as an adaptive response to conditions of low light availability. In the present study, the LWR value increased proportionally to the intensity of the shading, as compared to the control plants; therefore, in these plants, the biomass was allocated more to the assimilating surface formation, so the LWR value in plants placed under a green cover indicated the importance of light quality in addition to the amount of light in the parameters related to growth.
On the other hand, it was found that the light in the far red and green bands accentuated the effects induced by the shading, rather than the green light or the far red light separately. In this respect, this effect has been observed both in the morphology and in the gene expression of plants exposed to these factors (Wang and Folta, 2013). In this sense, the effects found in the present study in terms of growth and other evaluated variables were the result of the combined action of shading and light filtering through the green filters, which, according to Tab. 1, induced a value in the far red/red ratio that was inversely proportional to the reduction of the light, since the combination of the shade, the green light and the infrared light induced morphogenic responses in the plants that helped them to adapt better to the abiotic stress caused by an environment of this nature.
Water uptake and water use efficiency
The water uptake was reduced by 40.9-44.9% in the plants grown under the green covers, in comparison with the control plants, with statistically significant differences between the control plants and the set of plants exposed to the covers (Tab. 2). In relation to this variable, Casierra-Posada et al. (2014b) did not find statistical difference in the water uptake of Beta vulgaris plants placed under green and transparent polypropylene covers even though the leaf area in these plants was higher under the green cover than under the transparent one. In contrast to the results found in the present study for this result, the leaf area showed no statistical difference in any of the treatments, despite which, the water uptake was higher in the control plants in relation to all the plants under the green covers, suggesting that stomatal activity would be involved in this response, as indicated by Wang et al. (2009), who found that Cucumis sativus plants exposed to green, yellow and red light had a lower stomatal index than those exposed to white light, suggesting that stomatal density and stomatal index depend on the quality of light and purple and blue light also regulate most of the genes encoding the key enzymes in the Calvin cycle, while the green, yellow and red light regulates them. Additionally, Schoch et al. (1980) confirmed that the stomatal index is a function of the radiation received and its variations during the days preceding the differentiation of leaf stomata.
The values recorded for the water use efficiency were by 23.0, 25.3 and 53.7% in the plants exposed to the green covers with 55.4, 85.8 and 90.1% light reduction, respectively, as compared to the control plants, with statistically significant differences (Tab. 2). In contrast to this result, Casierra-Posada et al. (2014a) did not find statistical differences in the WUE value in Capsicum annuum plants, which grew without cover and those that were under the influence of a green polypropylene cover. Therefore, according to Hatfield et al. (2001), who stated that the WUE is a measure of the efficiency of the stomata to maximize the photosynthesis, reducing the loss of water by transpiration. Then it can be inferred that, in the present study, both the shading and the color of the cover induced a very low yield in relation to the WUE in the sisal plants. In this regard, Gil-Marín et al. (2006) found that water stress, plastic mulch and partial shading did not affect stomatal resistance or leaf transpiration in CAM Aloe vera plants. Stomatal resistance increased exponentially with the increase in air temperature and vapor pressure deficit and decreased with higher relative humidity. Additionally, these authors also indicated that the transpiration rate of the Aloe plants was remarkably low. The highest perspiration occurred at night, when the stomata were open, and, during the day, the transpiration was negligible or absent because the stomata were partially closed when the air temperature reached its maximum level. It could be thought that the lack of cover in the control plants of the present study was responsible for the increase in water uptake in the control plants, since the others were under cover, despite the fact that the plants of the present study and those studied by Casierra-Posada et al., (2014a) were exposed to similar environmental conditions although the results were very different. In addition, Fucraea hexapetala (syn. Furcraea macrophylla) is a CAM plant (Crassulacean acid metabolism), according to Casierra-Posada and Gonzalez (2009), and, therefore, has high energy requirements; however, CAM species can thrive in habitats experiencing contrasting light intensities, ranging from shaded habitats to full sun exposure. In addition, it is unknown how CAM species can meet energy and carbon requirements when they are suddenly exposed to shaded conditions in order to ensure survival (Ceusters et al., 2011). For this purpose, CAM plants must make use of their plasticity to adapt to the environmental offer of shading, which implies a low luminous intensity and low red/far red ratio. Therefore, based on the drastic reduction in the values of all the variables involved in growth, it can be inferred that the Furcraea hexapetala plants presented a low plasticity that allowed them to adapt to and survive the shading conditions evaluated in the present study.
Allometric distribution and production of dry material
The total dry weight per plant was reduced by between 80.8 and 94.0%, as compared to the control plants, in the group of plants exposed to shading with green filters, without taking into consideration the level of light reduction, with significant differences between the control treatment and the shaded group (Fig. 3). In this regard, Casierra-Posada et al. (2015) did not find statistical differences in Beta vulgaris plants exposed to polypropylene covers that were transparent, green, red and blue. These authors mentioned that, although the blue filter with the highest reduction (62.3%) did not show statistical differences with the response of the plants that were exposed to red, green or transparent color filters, phytochrome is involved in the response of plants to the production of dry mass since the yellow cover induced the greater production of dry mass in the plants.
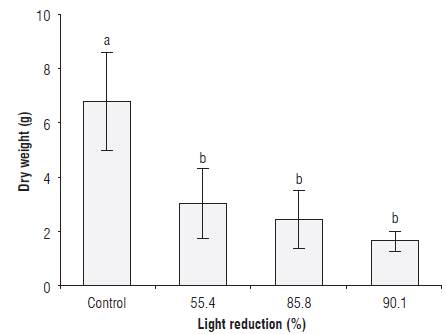
FIGURE 3 Total dry weight of sisal plants (Furcraea hexapetala Jacq.) exposed to shading with green polypropylene filters. The bars in each column indicate the standard deviation. Different letters indicate statistical significance according to Tukey's honest significance test (P≤0.05).
A variable related to the production of dry matter in plants is the electron transport rate (ETR). Casierra-Posada and Ávila-León (2015) found that, in Calendula officinalis plants exposed to shading, the ETR value was reduced by 17.3%, as compared to plants placed in full illumination. These authors mentioned that the magnitude of the electron transport rate in low light conditions depends on the incident radiation and the size of the antenna. Therefore, given that, in the present study, the CCI was reduced and that the genus Furcraea sp is light demanding because it is a CAM plant, the amount of dry matter per plant was severely compromised as a result of shading.
We found an increase of 24.2, 28.2 and 73.5% in the values of the root to shoot ratio in the plants placed under 55.4, 85.8 and 90.1% light reduction with the green filters, respectively, as compared to the control plants, with statistically significant differences (figure 4). In Fragaria sp plants, Casierra-Posada et al. (2012a) found that the value of the root to shoot ratio in plants placed under a green polypropylene cover was 86.7% above the value found in plants under a transparent cover, while Casierra-Posada et al. (2014b) found no difference in the value of this variable in Beta vulgaris plants exposed to green and transparent polypropylene covers. In agreement with the findings of the present study, Casierra-Posada et al. (2014b) found that, in Fragaria sp plants, a green polypropylene cover presented a greater accumulation of dry mass in the roots, in relation to yellow, blue, red, transparent covers and a control without cover. These authors commented that both the variations of the root to shoot ratio and the changes in the dry mass allocation in the different organs of the plants were the result of the interaction of the quality of the light and the shading provided by the covers because there are anatomical, morphological and physiological parameters that induce greater or lesser plasticity, given variations in the quality of light incident on the plants. In addition, according to Hirose (1987), the optimal partition theory assumes that plants respond to variations in the availability of resources through the preferential partition of resources between different organs in order to optimize the capture of light and maximize the growth rate.
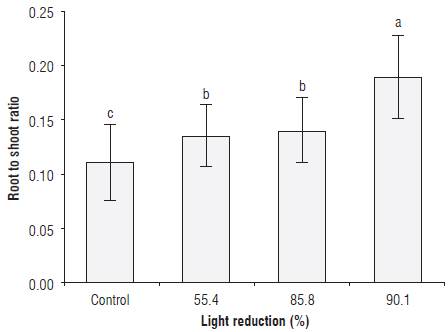
FIGURE 4 Root to shoot ratio in sisal plants (Furcraea hexapetala Jacq.) exposed to shading with green polypropylene filters. The bars in each column indicate the standard deviation. Different letters indicate statistical significance according to Tukey's honest significance test (P≤0.05).
For dry matter partitioning, statistically significant differences were observed in the amount of dry mass assigned to the roots and to the stem, but not to the leaves. In the roots, a statistically significant difference was observed between the control treatment plants and the set of plants placed under the green covers, independently reducing the light through the green covers. In this case, the amount of dry mass assigned to the roots of the plants in the treatment group under shading increased 2.9-6.0%, as compared to the value recorded in the control treatment plants (Fig. 5). The light reduction levels of 55.4, 85.8 and 90.1% reduced the amount of dry mass accumulated in the stems, as compared to the stems of the control treatment plants (Fig. 5), by 4.6, 10.9 and 13.4%, respectively. Although no statistical difference was found in the amount of dry matter assigned to the leaves, an increase of this variable can be observed in Figure 5 in the plants placed under the green covers, in relation to the control plants. The leaf area values for all treatments were in the range of 164.1-125.5 cm2. In agreement with these findings, Casierra-Posada et al. (2014b) found that Fragaria sp plants under a green polypropylene cover presented a greater accumulation of dry mass in the roots, in relation to yellow, blue, red, and transparent covers and a control without cover. These authors commented that variations in the root to shoot ratio and changes in the dry mass allocation in the different organs of plants are the result of the interaction of the quality of the light and the shading provided by covers; therefore, it can be inferred that, in the present study, the color of the cover, the shading and the low far red/red ratio value induced photomorphogenic modifications in the Furcraea hexapetala plants with the purpose of adapting to the environmental supply.
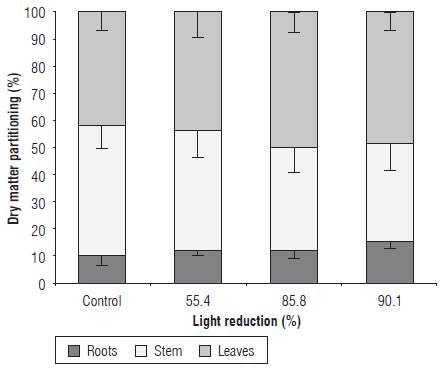
FIGURE 5 Dry matter partitioning in sisal plants (Furcraea hexapetala Jacq.) exposed to shading with green polypropylene filters. The bars in each column indicate the standard deviation
Finally, Kim et al. (2004) pointed out that the response of different species of plants or cultivars that grow exposed to different light environments is too complicated to interpret using a quantitative parameter of light quality. Additionally, according to Casierra-Posada and Peña-Olmos (2015), the detection of influences of the adjacent spectrum segments on plants is difficult because the spectrum is continuous, with no apparent margins between adjacent colors. When analyzing the results, it can be inferred that the Furcraea hexapetala plants presented little tolerance to the conditions of low illumination and low red/far red ratio induced by the green polypropylene covers; in addition, the difficulty of the studied plants having a CAM-type metabolism was seen, which were not only exposed to shading, but also to the placement of superimposed, green polypropylene layers that reduced the red/far red ratio. This environmental supply constituted a factor of severe abiotic stress for the plants, which did not have enough plasticity to tolerate it and, therefore, the growth variables were severely affected.
Conclusions
With the present results, it can be concluded that sisal plants (Furcraea hexapetala) have a low phenotypic plasticity to adapt to the conditions of an environment with diminished illumination and an enriched green color band. This environment induced profound changes in the values of the allometric variables and, therefore, in the architecture of the plant bodies, such as the root to shoot ratio and the leaf area ratio, whose values were severely increased. Similarly, the values of the variables involved in plant growth, such as chlorophyll content index, water use efficiency and relative growth rate, among others, were strongly diminished.

