











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Cotton farming is one of the main activities of the Brazilian agribusiness. The increasing use of more productive varieties worldwide, with a higher nutrient demand, requires a better knowledge on the nutritional relationships in cotton (Rochester and Constable, 2015). In this context, a better knowledge of the nutritional dynamics is important for the establishment of an efficient fertilization program.
Gossypium hirsutum L. - species of greatest use for cotton production in Brazil - is responsive to the application of micronutrients, especially in low natural fertility areas. Among the micronutrients, boron (B) and zinc (Zn) are the most limiting elements to achieve better yields (Araújo et al., 2012; Araújo et al., 2013). B is directly related to the metabolism of the ribonucleic acid (RNA), different functions of the plasmatic membrane and constitutes the structure of the cell wall and pectic substances associated with it, especially the middle lamella (Wimmer and Eichert, 2013). Zn is required in many active sites of different proteins, such as carbonic anhydrase and superoxide dismutase. The deficiency of this nutrient reduces the photosynthetic rate through the modification of the activity of enzymes involved in the process of carbon fixation (Broadley et al., 2007; Assunção et al., 2013).
B-Zn interaction influences different metabolic processes of the plant, interfering with the mineral composition through the stimulation or inhibition of the absorption of other nutrients (Baxter, 2009). These relationships are highly variable and may occur inside the cells or in the rhizosphere (Morgan and Connolly, 2013). The B-Zn interactions in cotton are correlated (Araújo et al., 2012) and the reduction in B absorption as a function of Zn is common. Besides cotton (Araújo et al., 2012), this result has already been observed for lemon (Rajaie et al., 2009), olive tree (Jasrotia et al., 2014) and wheat (Nasim et al., 2015). Thus, the effect of the B-Zn interaction on the absorption of these nutrients is evident. However, there is still limited information on the effect of B-Zn interaction in the absorption dynamics of other micronutrients, such as Cu, Fe and Mn.
Many nutritional relationships have been observed between micronutrients (Baxter, 2009; Milner et al., 2013; Aibara and Miwa, 2014). These nutritional interactions have been cataloged as complex, with initial modification at a molecular level, resulting in a variation of the absorption and mineral composition of the plants. Thus, the observed nutritional interaction depends on the availability of nutrients and on the analyzed plant species (Araújo et al., 2012; Araújo et al., 2013). Therefore, the need for studies that address the classic nutritional interaction between B and Zn, evaluating the absorption of Cu, Fe and Mn is evident. Understanding the micronutrient interaction would enhance the use of fertilizers, reducing production costs. The hypothesis of this study is that the combinations of B and Zn in nutrient solution modify the total content and the efficiency of absorption, use and transport of Cu, Fe and Mn in cotton. Given the above, the main objective of this research was to evaluate the effect of B and Zn concentrations on the absorption of Cu, Fe and Mn in cotton cultivated in nutrient solution.
Materials and methods
The experiment was conducted in an agricultural greenhouse of the Plant Production Sector of the State University of Mato Grosso do Sul (UEMS), in Aquidauana-MS, at the geographic coordinates 20°28' S and 55°48' W with an altitude of 174 m a.s.l. The experimental design was completely randomized with three replicates, in a 4x5 factorial scheme, corresponding to four concentrations of B (0, 20, 40 and 80 μM L-1), applied in the form of boric acid, and five concentrations of Zn (0, 1, 2, 4 and 8 μM L-1), applied in the form of zinc sulfate.
The experimental units consisted of plastic pots with 3 L capacity, filled with washed and sterilized quartz sand.
Cotton seeds, cv. FiberMax 910, were placed to germinate on trays with moistened sand. Five days after emergence, when cotyledon leaves developed, three seedlings were transplanted to each experimental unit, where they received a complete and diluted (1/5) nutrient solution (Epstein and Bloom, 2006). At 28 days after emergence, thinning was performed leaving only one plant in each experimental unit. The nutrient solution application started according to each treatment, irrigating three times a day with deionized water. In the treatment solutions lacking a specific nutrient, the concentrations were identical to those of the complete solution, except for the absent nutrient.
The nutrient solution showed the following composition: 6.0 mL of 1 mol L-1 KNO3; 4.0 mL of 1 mol L-1 Ca(NO3)4-H2O; 2.0 mL of 1 mol L-1 NH4H2PO4; 1.0 mL of 1 mol L-1 MgSO4-7H2O; 1.0 mL of 0.2 mol L-1 Fe-EDTA; 1.0 mL of 0.05 mol L-1 KCl; 1.0 mL of 0.02 mol L-1 H3BO3; 1.0 mL of 0.002 mol L-1 MnSO4-H2O; 1.0 mL of 0.002 mol L-1 ZnSO4-7H2O; 1.0 mL of 0.0005 mol L-1 CuSO4-5H2O; 1.0 mL of 0.0005 mol L-1 H2MoO4 (85% MoO3).
At 115 days after emergence, the plants were collected and divided into roots, shoots (stem and leaves) and fruits (boll). All the collected plant material was rinsed in a detergent solution at 3 mL L-1, running water, 0.1 mol L-1 HCl solution and deionized water, respectively. The samples were placed in paper bags and then dried in a forced-air oven at 65°C for 72 h. After drying, the plant material was ground in a Wiley-type mill (Thomas Scientific, USA). The samples were subjected to nitric-perchloric digestion for the determination of Cu, Fe and Mn contents in the different plant parts (Malavolta et al., 1997).
The determination of Cu, Fe and Mn was used to calculate the absorption efficiency (mg g-1) = [total content of nutrient in the plant (mg)/root dry matter (g)] (Swiader et al., 1994); transport efficiency (%) = [(content of nutrient in the shoots (mg)/total content of nutrient in the plant (mg)) x 100] (Li et al., 1991); and use efficiency (g2 mg) = [(total dry matter produced (g))2/total content of nutrients in the plant (mg)] (Siddiqi and Glass, 1981).
The results were statistically assessed with an analysis of response surface, using the statistical package SAS and adopting a 0.05 significance level. Initially, an analysis of variance was performed and, according to the significance of the F test, the study of polynomial regression was conducted (for cases with significant interaction), through the RSREG procedure.
Results and discussion
The interaction between the B and Zn treatments influenced the Fe content in the boll, total Fe in the roots and in the boll, total Mn in the shoots and transport efficiency of Fe and Mn (Tab. 1). There was a significant response (P<0.05) of B concentrations for Cu content in the boll, total Cu in the roots and in the boll, Fe content and total Fe in the roots, Mn content in the boll, Cu use efficiency and the efficiencies of absorption of Fe and Mn (Tab. 1).
TABLE 1 Summary of the analysis of variance: coefficient of variation and P-value for the effect of B, Zn and the B-Zn interaction for content, total and efficiency of absorption, transport and use of Cu, Fe and Mn.
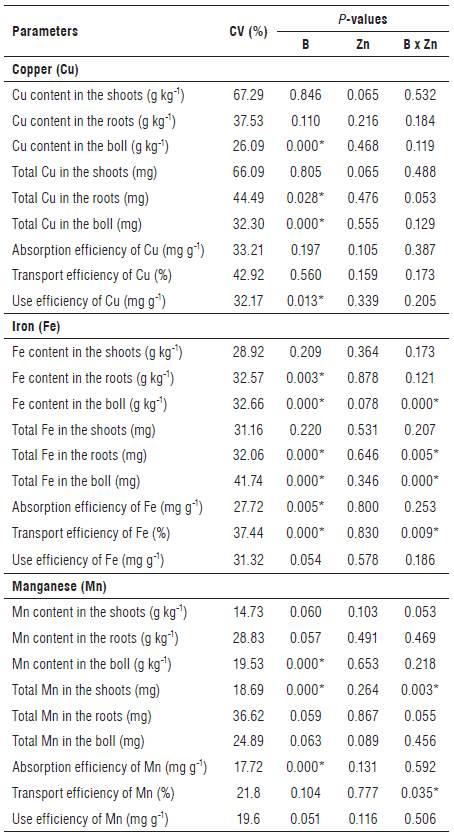
B: Boron; Zn: Zinc; CV: Coefficient of variation. *Significant by F test at P<0.05.
Zn doses in the solution did not affect individually any of the studied variables. Various studies indicate antagonistic relationship between Zn and cationic micronutrients, such as Cu, Fe and Mn. This interaction occurs due to the competition of the cations for the absorption sites (Broadley et al., 2007; Baxter, 2009; Assunção et al., 2013). This study did not find any antagonistic relationship between Zn and other cationic micronutrients (Fe, Mn and Cu), disagreeing with the results reported by Rajaie et al. (2009), Aref (2011) and Lima Neto and Natale (2014) in other plant species. Hence, only B doses and the combined doses of B and Zn influenced the absorption of Cu, Fe and Mn in cotton (Tab. 1).
Cu content and total Cu in the cotton fruit were influenced by the B concentrations in the solution. The increase in B concentrations in the solution until 40 μM L-1 promoted a reduction in the Cu content and total Cu in cotton bolls (Figs. 1A, 1B). B doses higher than 40 μM L-1 increased Cu content and total Cu in the bolls. Aref (2011) observed an increment in the concentration of Cu in maize leaves with the increase in B concentrations in the planting substrate. Ahmed et al. (2011) observed a positive effect of B doses on Cu content and total Cu in different parts of cotton plants. However, the total Cu in cotton roots linearly decreases with the increment in B concentrations in the solution (Fig. 1C), agreeing with the results found by Esringü et al. (2002) who observed a rgjeduction of Cu concentration in strawberry roots with the increase in the supply of B reporting an antagonistic relationship between the nutrients.
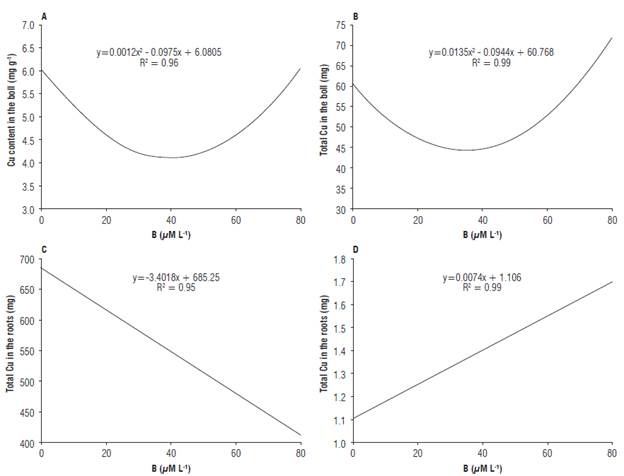
FIGURE 1 Content of Cu in the boll (A), total Cu in the boll (B), content of Cu in the roots (C) and Cu absorption efficiency (D) of cotton in response to different B concentrations in the nutrient solution.
Cu use efficiency for the production of dry matter was positively influenced by the supply of B in the solution, so that the increase in B concentrations promoted linear increment in the Cu use efficiency of cotton plants (Fig. 1D). B is related to various biochemical processes in the plants; thus, its concentrations in the cultivation substrate may directly or indirectly affect the use of Cu by the plant (Malavolta, 2006; Wimmer and Eichert, 2013).
B concentrations in the solution promoted a significant increment in Fe content and total Fe in cotton roots (Figs. 2A, 2B). Similar results were obtained by Esringü et al. (2012) in strawberry, in which the Fe content in the roots increased with the concentration of 10 μM L-1 of B, decreasing at the concentration of 20 μM L-1 of B. These results suggest that B has certain affinity for Fe and that there may be a synergetic relationship between the nutrients. Rajaie et al. (2009) also observed significant increase in the concentration of Fe with the increment in B levels in Citrus aurantifolia.
Fe content and total Fe in the cotton fruit were significantly affected by the interaction between B and Zn in the solution. The increase in B concentrations in the solution promoted the reduction in Fe contents and total Fe in the fruit with the increment in Zn concentrations (Figs. 2C, 2D). Ahmed et al. (2011) reported that the Fe content, in different parts of the cotton crop, significantly increased with the increment in the applied levels of B. The increase in Fe concentration in the fruit with the increasing levels of B has also been reported in guava (Salvador et al., 2003) and in Citrus aurantifolia (Rajaie et al., 2009), disagreeing with the results obtained in the present study. There is a lack of available information about the influence of B on the absorption of other micronutrients, and the results vary according to the analyzed plant species.
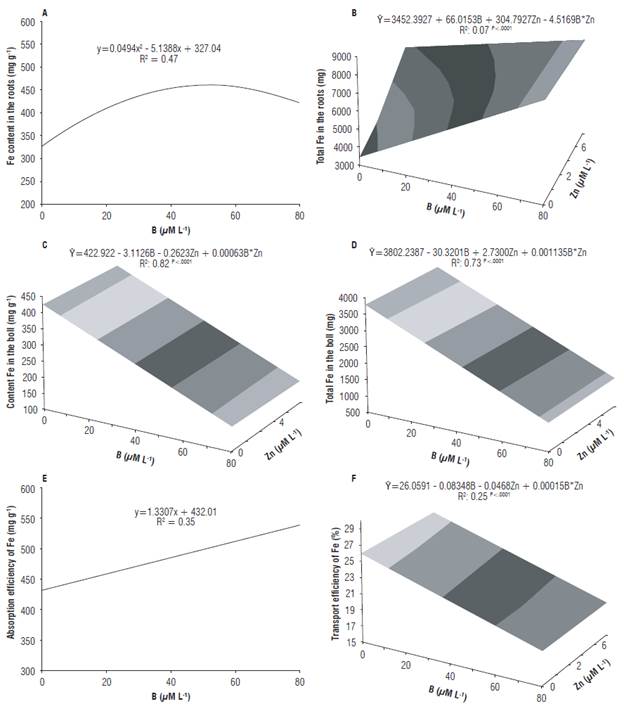
FIGURE 2 Content of Fe in the roots (A), total Fe in the roots (B), content of Fe in the boll (C), total Fe in the boll (D) and efficiency of absorption (E) and transport (F) of Fe of cotton in response to different B and Zn concentrations in the nutrient solution.
Fe absorption efficiency positively responded to the B supply in the solution, so that the increase in B concentrations promoted increment in the Fe absorption efficiency of cotton (Fig. 3E). Fe transport efficiency was affected by the interaction between B and Zn in the solution, demonstrating an antagonistic relationship between B and Zn at the highest concentrations of B, and a synergetic relationship at the highest concentrations of Zn (Fig. 3F). It is possible to observe an increment of 25% in the transport of Fe by the roots with the increase in Zn concentrations at 0 μΜ L-1 of B.
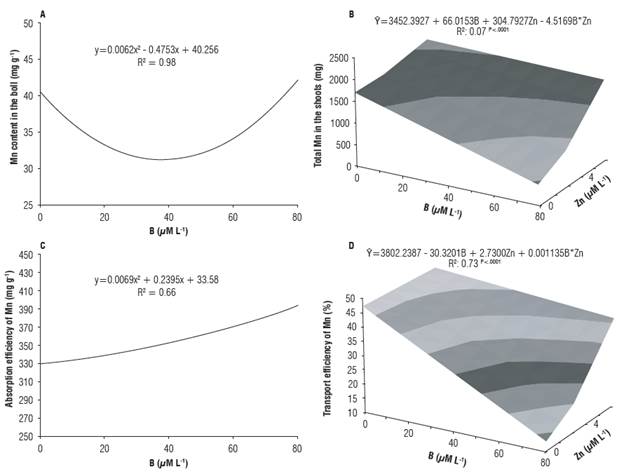
FIGURE 3 Content different B and Zn of Mn in the boll (A), total Mn in the shoots (B) and efficiency of absorption (C) and transport (D) of Mn of cotton in response to concentrations in the nutrient solution.
The Mn content in the fruit decreased with the increment in B concentrations, until 40 μΜ L-1 of B, with an increase at the highest concentrations of the nutrient (80 μΜ L-1), the same behavior observed for Cu content and total Cu in the cotton fruit (Fig. 3A). Aref (2011), working with maize plants, observed that the Mn content was affected by B fertilization, with a reduction in leaf Mn content under these conditions. On the other hand, Esringü et al. (2012) observed an increment in leaf Mn content with the increase in B concentrations, until 10 μM L-1, with reduction at the highest B concentration (30 μM L-1). In contrast, the data of the present study indicate that Mn positively responded to B only at the highest concentration of the nutrient.
The total Mn in the shoots and Mn transport efficiency were negatively influenced by the increase in B concentration at 0 μM L-1 of Zn and positively influenced at 8 μM L-1 of Zn (Figs. 3B, 3D). It is possible to observe an increment of 49% in the transport of Mn by the roots with the increase in Zn concentrations at 0 μM L-1 of B. This reduction in Mn content and total Mn in cotton roots and shoots in response to the increase in B concentrations may be due to the effect of dilution or the antagonistic relationship between B and Mn. The source of B can influence, as a regulator or inhibitor, the accumulation and use of other nutrients (Aref, 2011). Mn absorption efficiency linearly increased with the increment in B concentrations (Fig. 3C). B is involved in physiological processes that control the absorption and transport of Cu, Fe, Mn and Zn (Malavolta, 2006; Wimmer and Eichert, 2013). Dursun et al. (2010) also observed increase in Cu, Fe and Mn contents in leaves of tomato and pepper as a function of B doses.
Conclusions
Cu content and total Cu in the fruit, total Cu in the roots, Cu use efficiency, Fe content in the roots, Fe absorption efficiency, Mn content in the fruit and Mn absorption efficiency of cotton are influenced by the concentrations of B in the solution.
The B-Zn interaction affected total Fe in the roots, Fe content and total Fe in the fruit, Fe transport efficiency, total Mn in the shoots and Mn transport efficiency; in addition, Zn acts differently according to the B supply and vice versa.

