











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
The common bean (Phaseolus vulgaris L) is a widely cultivated legume that originated in the New World, which has been domesticated in Meso and South America (Chacon et al., 2005). Dry beans remain the primary source of dietary protein carbohydrates, minerals and fiber in most developing countries, making them the third most important food legume crop worldwide (Singh and Muñoz, 1999).
The small-scale agriculture of the common bean is constrained by the multiplicity of biotic and abiotic factors that affect crop productivity and quality during the growing season. In addition, one of the most important biotic factors is predation of dry bean seeds by post-harvest pests (bruchids), which largely affect the overall food grain production and consumption. It is estimated that 13% out of a total loss of 37% in global agricultural production is caused by pests and diseases (Obeng-Oferi, 2007).
A cost effective strategy is required in order to provide an efficient and sustainable control of bean infestation by bruchids. A combination of genetic resistance and biological and cultural control methods presents a sustainable way to deal with this problem. In efforts to develop bruchid-resistant beans, researchers have identified and introgressed different genetic resistance factors from wild relatives into common bean. The principal resistance factors in mature bean seeds are arcelins (Osborn et al., 1988a; Osborn et al., 1988b; Zaugg et al., 2013).
Arcelins (Arcs) belong to the lectin-like family described in eight allelic variants in various accessions of the wild common bean, P. vulgaris, which exhibit different levels of resistance to bruchid insect pests. They are seed storage proteins associated with lectin or the arcelin-phytohae-magglutinin-alpha amylase inhibitor (APA) complex locus (Zaugg et al., 2013).
Although homologous to lectins, arcelins have a different intrinsic specificity for complex sugars, creating a mechanism of toxicity to bruchids, as opposed to the monosaccharide-binding affinity of true lectins (Fabre et al., 1998; Blair et al., 2010). The toxic properties of arcelins may be related to their ability to recognize and interact with glycoproteins and other constituents of the digestive tract membrane, as well as to their capacity to directly bind to the intestinal cells of insects (Minney et al., 1990; Fabre et al., 1998; Paes et al., 2000).
One or two dimensional sodium dodecyl sulfate gel electrophoresis (SDS-PAGE) techniques have been used for seed protein characterization to distinguish Andean bean genotypes from those of Mesoamerica according to the electrophoretic mobility of phaseolins (Gepts et al., 1986; Gepts and Bliss, 1986; Kami et al., 1995).
Additionally, in spite of the genetic studies that have been carried out and the approaches that have been used to study the P. vulgaris proteome (Natarajan et al., 2013; Parreira et al., 2016), the unique identification of arcelins in the different varieties is complicated.
The present study provides the first inmuno-proteomic approach used for the characterization of arcelins from the common bean, P. vulgaris, as a preliminary germplasm characterization.
Materials and methods
Seed material
Mature seeds from eight common bean (P. vulgaris) accessions from the germplasm bank of INIA-CENIAP (Maracay-Venezuela) (10°17T4" N, 67°36'02'' W, and 480 m a.s.l.) were used. Information about the germplasm used in the present study (passport data) was published by Gamero et al. (2010). The seeds were selected from different states of Venezuela, showing dissimilar commercial values, consumer preferences and seed color. Commercial seeds corresponded to the states of Victoria, Tacarigua, Montal-ban, and Tenerife, and study cultivars NAG-7S, DOR-500, MGM-08-02-026 and MGM-10-02-108 were also selected.
Protein extraction and samples preparation
Dry beans were milled, and 100 mg of flour were suspended in 20 mM Borate buffer pH 9 with 30 mL-1 of a protease inhibitor mix (Amersham Biosciences, UK) and stirred for 1 h at 37°C. The samples were then centrifuged for 15 min (14,000 g) at 4°C, and the supernatant was used for the assays.
Protein determination
The protein concentration was determined with the Bradford method (1976) using the Bio-Rad assay reagent (Bio-Rad, USA) and bovine serum albumin as the standard.
Polyacrylamide gel electrophoresis
The proteins were separated with SDS-PAGE according to the method of Laemmli (1970). Briefly, 20 ug of proteins were dissociated with heating at 90°C for 5 min in the presence of a denaturing buffer (20 mM Tris-HCl pH 8.6 containing 1% SDS, 8.3% glycerol and 0.5% 2-mercaptoethanol) and separated with electrophoresis in 12% polyacrylamide gels. The proteins were stained with Coomassie Brillant Blue and destained in 7% acetic acid.
Two-dimensional electrophoresis (2-DE)
2-DE was performed as described by O'Farrell (1975); 1-D isoelectric focusing (IEF) was conducted in a ZOOM® IPGRunner™ (Invitrogen, USA), whilst the SDS-PAGE was carried out using a Mini-protean II (BioRad, USA). Each sample was run in triplicate. The protein samples were diluted in a rehydration solution (De Streak™, Amersham Biosciences, UK) with 1% IPG buffer pH 3-10 to a final volume of 155 µl (ZOOM® IPGRunner™ Cassettes, Invitrogen, USA) in a 7 cm IPG strip (Amersham Bio-Sciences, UK), pH 3-10 overnight; the proteins were then focused at a maximum voltage of 2,000 V. After IEF, the proteins were reduced and alkylated by soaking the IPG strips in the equilibration buffer (6M urea, 2% SDS, 30% glycerol, 50 mM Tris-HCl, pH 8.8) containing 1°% DTT for 15 min at room temperature and then equilibrated in 4% iodoacetamine (IAA) for 15 min. The equilibrated IPG strips were sealed on top of the SDS-PAGE gel (12% polyacrylamide) using 0.5% agarose. The 2-D SDS-PAGE was carried out using a Tris-glycine-SDS buffer system at 200 V until the dye front reached the bottom edge of the gel. The protein spots were visualized with staining using PlusOne™ silver staining kit (Amersham Bio-Sciences, UK).
Immunoassays
The proteins from the 2-D SDS-PAGE were transferred at 100 V for 1 h to a nitrocellulose membrane for Western blot analysis. The membrane was Ponceau stained for 1.5 min to visualize the transferred protein and air dried. The membranes were re-wetted in a blocking solution composed of 2% non-fat dry milk in tris-buffered saline (TBS) with 0.5% Tween-20 (TTBS) and finally incubated for 1 h. The membrane was then probed for lectin-like proteins during 1 h of incubation with rabbit anti-lectin polyclonal antibodies (Sparvolli and Bollini, 1998) diluted at 1:6000 in a blocking mixture at 1:5 in TTBS. The immune serum was pre-absorbed overnight with 100 ng of bean flour from arcelin-null P. vulgaris in 1 mL of TBS to eliminate any unspecific cross-reactivity (Lioi et al., 2003). The bound antibodies were detected with horseradish peroxidase conjugated anti-rabbit IgG antibodies (1:2000 in TTBS) using the ECL™ Western Blotting Detection Reagents (GE Healthcare Life Sciences, UK), following the manufacturer's instructions. The image of the chemiluminescent signals was recorded on an X-ray film and developed according to standard procedures in a darkroom.
Image analyses
2-DE protein patterns were obtained as digitalized images using a high-resolution system (ProXCISION, PerkinElmer, Inc., USA). Spot detection, quantitation, and analysis were performed using Phoretix 2D (Nonlinear Dynamics, UK).
Protein identification
Spots of interest were excised, picked and placed in a 96-well plate using the protein picking workstation ProXCISION (PerkinElmer, Inc., USA). The MultiPROBE II PLUS HT EX (PerkinElmer, Inc., USA) Robotic Liquid Handling System was used for enzymatic digestion. Briefly, the method involves the following steps: (i) reduction with 10 mM DTT; (ii) alkylation with 55 mM IAA; (iii) overnight tryptic digestion (6 ng µL-1 enzyme); (v) sample loading onto MALDI target using 1 µl of the tryptic digests mixed at 1:1 with a matrix solution (a saturated solution of α-cyano-4-hydroxyxinnamic acid in 0.1% TFA-acetonitrile 50:50).
A MALDI prOTOF™ 2000 (PerkinElmer, Inc., USA) was used for peptide analysis. Calibration was performed using a commercial mixture of peptides, PepMix (Perkin Elmer, Inc., USA), of known molecular masses. TOFworks™ 1.0 (PerkinElmer, Inc., USA) was used for the data processing and for the peak list generation. The most intense peaks of the obtained spectra were identified with MASCOT PMF searching (http://www.matrixscience.com) using the Swiss-Prot and NCBI databases.
Results and discussion
Different methods have been used for the identification and characterization of seed storage proteins in beans, including protein chemistry for purification of storage proteins for further structural and functional studies, proteomics protocols, and immunoassay techniques (Osborn et al., 1988a; Osborn et al., 1988b; Fabre et al., 1998; Sparvolli and Bollini, 1998; Paes et al., 2000; Lioi et al., 2003; Zaugg et al., 2013).
Moreover, in a previous study (Bernal et al., 2006), an approximation of the comparative immunoassay technique with antibodies against the lectin family was used; however, the technique was nonspecific for the identification of proteins with mass spectrometry. The present study proposes a combination of proteomics and immunoassays techniques to successfully display the entire Arcelin catalogue of P. vulgaris for comparative purposes.
From the lectin family, arcelins were described as proteins associated with the inhibition of some bruchid species development. Although initially found in a limited number of wild common bean accessions from Mexico, approximately 10% of wild common bean accessions from Mesoamerica express Arcs. Until recently, Arcs were thought to be restricted to wild accessions of common bean, but arcelin-like sequences were obtained from other Phaseolus species (Osborn et al., 1988a; Osborn et al., 1988b).
Proteins of 30 P. vulgaris commercial accessions from a germplasm bank were first analyzed with SDS-PAGE to select the varieties that presented greater differences in the regions corresponding to the proteins of interest, in this case arcelins; these differences were visualized with image analysis programs (Fig. 1). Figure 1 shows the first selection of seeds with important differences in the 25-40 kDa range, where the majority of arcelins resolved (dotted boxes on the image, Fig.1).
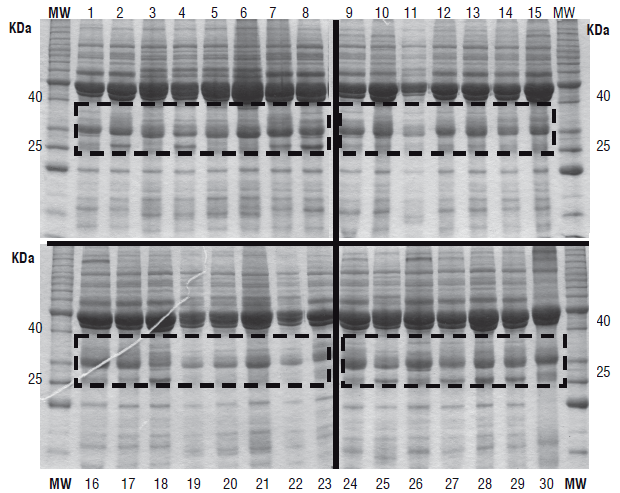
After this analysis, only 8 accessions (Fig.1: 1, 2, 4, 8, 15, 18, 22 and 25) were selected and further analyzed with two-DE. The comparative analysis of the proteome images using Phoretix 2D revealed an average of 104 spots on the different proteomes from P. vulgaris (Fig. 2 B-I). These experiments were performed in triplicate for each accession.
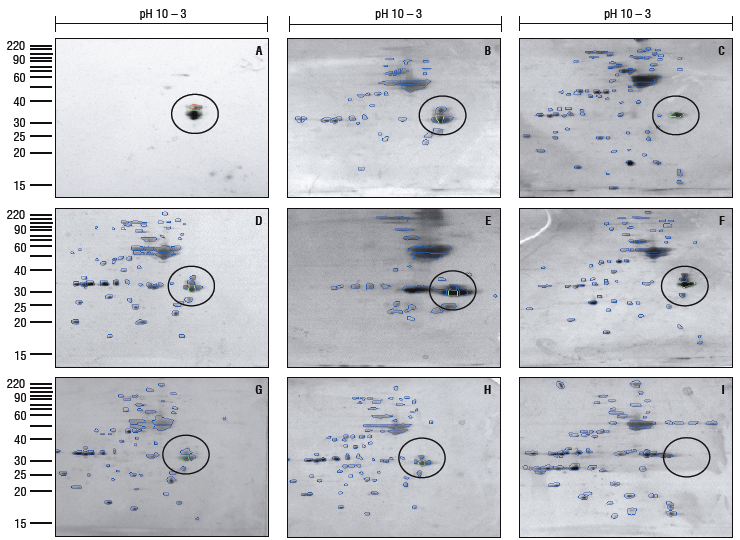
FIGURE 2 (A) 2-DE immunoblot of extracted proteins from a commercial variety of R vulgaris. (B-l) 2-DE of extracted proteins from eight different accessions. Immuno-detection of the arcelin region with a specific antibody (A). Circles highlight the Two-DE region corresponding to arcelins in the different proteomes according to the immunoblot and the Phoretix 2D profile analysis.
The immuno-detection of the arcelins was obtained from the 2-DE gels of extracted proteins from a commercial variety by using a specific immune serum against this family of proteins. Figure 2A shows the results of the immunoblot, with only a few spots detected in the range of 33-35 kDa and 5.5-5.7 pH, which correspond to the characteristics of arcelins. The immunoblot image was used as a reference to identify the same group of proteins in each proteome with Phoretix 2D (Fig. 2 B-I). The protein spot patterns were different in each P. vulgaris proteome, suggesting the presence of different arcelins in each case, although it was not possible to distinguish between subgroups of arcelins with this approach. We observed the presence of two, three, four and five spots related to the range of molecular weight and inmuno-detection of arcelins (Fig. 2).
Some authors have stated that the more promising arcelin variants that confer insect resistance are arcelin-1, arcelin-5 and arcelin-8 (Osborn et al., 1988a; Zaugg et al., 2013); however, there is not much information on the presence of other variants and the role of these proteins in bruchid toxicity is not clear. Moreover, genetic studies suggest that resistance to storage pests in fact depends on a combination of proteins from the APA locus (arcelin, phytohemagglutinin and a-amylase inhibitor), in which case the participation of the arcelin family can play a fundamental role (Blair et al., 2010).
More profound mass spectrometry analysis is needed to elucidate which arcelin variants are expressed in these common bean accessions. It is worth noting that assayed commercial varieties have shown spots related to arcelins (Fig. 2). It is interesting to observe this high frequency of arcelins in the Venezuelan commercial varieties of P. vulgaris.
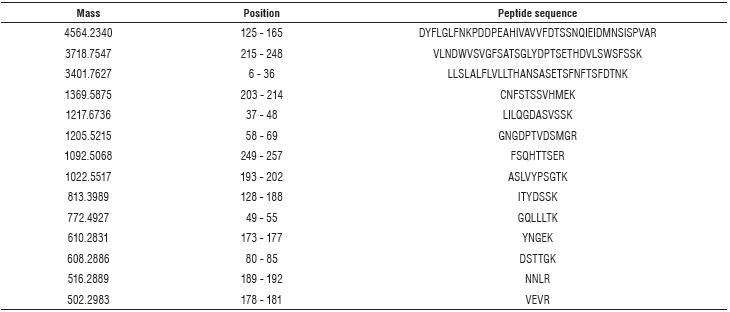
As shown in Table 1, it was possible to identify a group of peptides related to arcelins (98% identity) with mass spectrometry.
TABLE 1 Peptide sequences and mass-specs for arcelin identification. Theoretical pl: 6, 16 / Mw (average mass) 29375.87 / Mw (monoisotopic mass): 29357.62.
Despite the tools described in this paper, it was not possible to determine the type of arcelins present in each variety. These results further confirmed the arcelin nature of the immune-detected spots.
The large-scale study of the P. vulgaris proteome has been described in previous studies (Natarajan et al., 2013; Parreira et al., 2016); however, it does not present a useful tool for the study of protein families of interest, such as lectins. The inmuno-proteomic approach described in this study is a reliable alternative technique to easily localize/ identify a subset of proteins from complex bidimensional gels. This is the first immuno-proteomic approach used for the characterization of arcelins from P. vulgaris.


