











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
In 2016 in Colombia, an estimated production of2,623,700 t ha-1 was achieved on 126,100 ha, with Cundinamarca and Boyaca provinces generating 76% of the potato production (Riascos, 2016). Diacol Capiro and Pastusa Suprema are considered cultivars with high economic importance and represent 80% of the area cultivated for fresh consumption and industrial processing in the country (Ñústez, 2011).
In the Andean region of Colombia, yields of 50-60 t ha"1 have been reported for Capiro (Gómez and Torres, 2012) and 40-50 t ha-1 for Suprema (Pérez, 2015). However, these yields vary considerably by location or soil type, with an increase in the potential yield in the last decade resulting from technological improvements in certified seed management, irrigation, mechanization, and balanced fertilization.
In potato cultivars, a high application of nitrogen and potassium fertilizers facilitates significant yields (Quinchoa et al., 2010; Saravia et al., 2016), but there is low absorption efficiency of mineral nutrients by the plants, probably because of the shallow root systems (Poljak et al., 2011). This implies a low nutrient recovery of up to 50% for N (Vos, 2009) and 70% for K (Gómez and Torres, 2012) as a result of the low influx of potassium used by the roots (Rengel and Damon, 2008). On the other hand, other factors include the availability of water and nutrients in the soil (Saravia et al., 2016) as well as the plant genetics in terms of source-sink efficiency, which is variable for cultivars (Trehan and Singh, 2013) and directly affects the efficiency of nutrient transport to tubers (Poljak et al., 2011; Giletto and Echeverria, 2015; Fernandes and Soratto, 2016). Nutrient Use Efficency (NUE) in potato tubers has recently been researched, mainly for N by Poljak (2011), Saravia et al. (2016), and Marouani and Harbeoui (2016), and for K by Trehan and Singh (2013), and Wang and Wu (2015) in Group Chilotanum, while studies on the efficient use of nutrients in Group Andigenum are less common (Zebarth et al., 2012).
Potatoes of the Group Andigenum cultivated in the Andean region respond favorably to fertilization, mainly in soils with a low nutrient supply (Quinchoa et al., 2010). Nutrient uptake rates of about 0.2-0.3 kg m2 are seen (Quinchoa et al., 2010; Gómez and Torres, 2012), which can reduce the efficient use of fertilizers and negatively impact the profitability and environmental sustainability of potato production in the country. The efficiency of nutrient use for Capiro and Suprema cultivars has not been quantified. The present research evaluated the production potential, accumulation of dry matter, harvest index (HI) and NUEt for N and K in tubers in response to balanced fertilization in three soils with contrasting fertility; this study aimed to establish an optimal management of these mineral nutrients according to the cultivar and soil type.
Materials and methods
Location and soils
The experiments were conducted between 2013 and 2014 in three potato producing locations with contrasting soils on the Cundiboyacense plains in the Andean region of Colombia as shown in Table 1.
TABLE 1 Environmental and soil fertility characteristics at the study sites (from Gómez et al. (2017), with permission from Agronomia Colombiana).
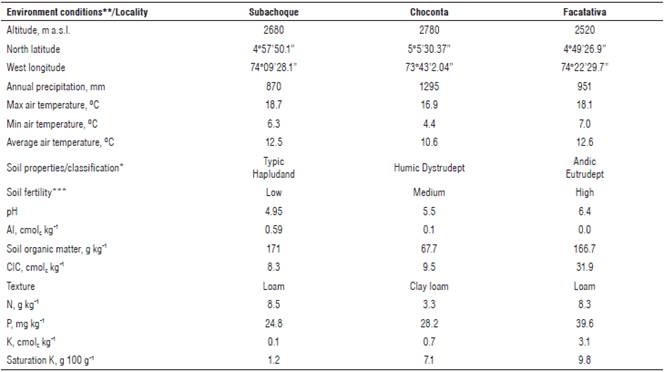
In each location, a randomized complete block design was established using a factorial arrangement in a divided subplots-mixed model with three replicates (one plant per replicate), where the main plot corresponded to the cultivars (Capiro and Suprema) and the subplots corresponded to two levels of fertilization (F0 and F1). The fertilizer doses F1 were not the same in the three locations (Tab. 2) due to the differences in initial soil fertility among the study sites (Tab. 1); therefore, the design was of incomplete nature due to the fertilizer dose factor. The amount of mineral nutrient applied, the sources of fertilizers, and the dose fractionation for the F1 level are shown in Table 2.
TABLE 2 Doses of mineral nutrients applied with fertilizers in the study sites (from Gómez et al. (2017), with permission from Agronomía Colombiana).
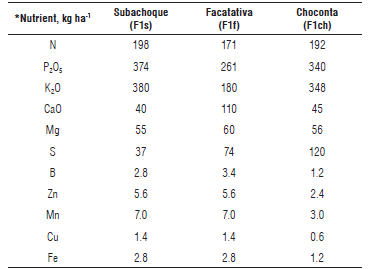
*Recommended fertilization rates derived from the soil-plant balance method (Castro and Gómez, 2013) and fertilizer dose fractionation according to historical references in the areas, where high yields have been obtained (>50 t ha-1): N, 60% at sowing and 40% at 45-50 days after sowing (das); P, 70% at sowing and 30% at 45-50 das; K, 30% at sowing and 70% at 45-50 das. Granulated fertilizer sources were: N-P, DAP; K, KCl (0-0-60), Potassium sulfate (0-0-50); Ca, Calcium Nitrate (25% CaO); Mg, Kieserite; Nutricomplet, complex source of micronutrients B, Zn, Cu, Mn and Fe based on sulfates.
Twelve combinations of factors were evaluated using three factors: two cultivars (Capiro, Suprema), three locations (Subachoque, Facatativa, and Choconta) and two levels of fertilization, F0 (unfertilized plots, initial soil fertility conditions) and F1 (fertilized plots). The factorial design with an intra-subject factor over time was associated with four phenological stages of tuber growth, and was adapted from Valbuena et al. (2010): Stage II, 70-75 days after sowing (das) (start of tuberization); Stage III, 90-100 das (flowering, maximum tuberization and start of tuber filling); Stage IV, 120-125 das (end of flowering, filling of tuber); Stage V, 150-160 das (maximum tuber filling and ripening). The sowing was done in experimental units of 50 m2 (135 plants/plot), with distances of 1 m between rows and 0.37 m between the plants and a density of 27,000 plants ha-1. The agronomic practices of irrigation, weed and phytosanitary management were carried out according to the needs of the locations, as a result the effects of external factors were minimized. For Subachoque and Choconta, dolomite type pre-sowing amendments were incorporated in the soils at rates of 1.5 and 1.0 t ha-1, respectively.
Plant sampling and analysis
At the four stages of tuber growth, four plants per experimental unit were evaluated. Destructive analysis of the leaves, stems and tubers was carried out. All parts of the plants were rinsed with deionized water. For each sampling, fresh leaves, aerial stems and tubers were weighed separately; a sample of 200 g (fresh weight) of each organ was placed in a paper bag and dried in the oven at 70°C for 72 h. The dry matter (DW) of each sample was then weighed, and the DW accumulation in each organ was evaluated per plant and per stage. All of the organs of the dried plants were ground using a 40 stainless steel mesh for subsequent chemical analysis. The harvest index (HI) was evaluated at phenological stage V: HI = (DWt/DWs)*100 according to Giletto and Echeverria (2015), where DWt is the tuber dry weight and DWs is the shoot dry weight. With dry samples, the nutrient concentrations in the tubers were determined according to the methodology of IGAC (2006). The amounts of N and K extracted by the tubers were calculated by multiplying the concentration of the nutrients by the DW accumulated by the tubers at each growth stage.
Efficiency use and recovery of mineral nutrients by tubers
The NUE indices were estimated by considering the treatments without fertilizer application (F0) with respect to the balanced fertilization in each location (F1). The N and K recovery efficiency by the tubers from fertilization, or acquisition efficiency (RFt), was calculated using the equation RFt = (Et1-Et0/amount of mineral nutrient supplied in the fertilizer)*100 (Tab. 2), where Et1 is the nutrient extraction by tubers in fertilized soils (kg ha-1) and Et0 is the nutrient extraction by tubers in unfertilized soils (kg ha-1). This equation was adapted from Fernandes and Soratto (2016). The efficiency use of N and K by the tubers (NUEt) was estimated as the accumulated dry matter in the tubers/nutrient accumulation in the tubers, as reported for potatoes by Poljak et al. (2011) and Rengel and Damon (2008) for N and K. In addition, we evaluated the efficiency of tuber production obtained per unit of accumulated nutrient (EPt) with the equation adapted from Prochnow et al. (2009): EPt = (FWT1-FWT0) / (Et1-Et0), where FWT1 is the tuber yield in the fertilized treatment and FWT0 is the tuber yield in the control treatment.
Statistical analysis
Multivariate analysis of variance was used, assessing differences in factor interaction with a confidence level of P<0.01. The Pearson correlation matrix for the qualitative variables was analyzed and the efficiency indices were adjusted. The statistical program SAS version 2014 was used.
Results and discussion
Yield, harvest index, and dry weight of tubers
The DWt and FWt presented significant differences (P≤0.01) in response to the phenology*fertilization interaction, with a higher yield obtained for Suprema in fertilized treatments (F1) in Subachoque (70.5 t ha-1) and Choconta (73.7 t ha-1), in soils with medium to low fertility (Fig. 1) and a higher DWt accumulation, between 17.1 and 19.98 t ha-1, respectively (Fig. 2). For Capiro, the highest yield was registered in fertilized soils with the highest K availability (Tab. 1): Humic Dystrudepts and Andic Eutrudepts in Choconta (67.3 t ha-1) and Facatativa (73.1 t ha-1), respectively (Fig. 1). which was directly related to the accumulation of DWt.
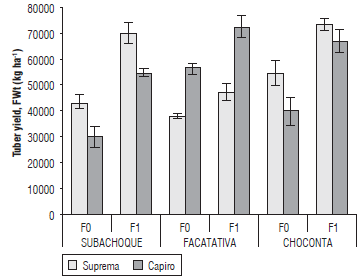
FIGURE 1 Yield (FWt) of the cvs. Capiro and Suprema at phenological stage V (maximum tuber filling and maturation) in the absence of fertilization F0, with respect to balanced fertilization by location: F1s, Typic Hapludands (Subachoque); F1ch, Humic Dystrudepts (Choconta), and F1f, Andic Eutrudepts (Facatativa). p<0.01 for fertilization {location*cultivar). Error bars indicate standard errors.
The yields of 70 t ha-1 obtained in both cultivars proved the high genetic potential of these Andean potatoes under optimal environmental and fertilization conditions; these yields were dependent on the soil-plant conditions, where a greater number of tubers and a better translocation of assimilates were promoted, exceeding the yields of 45 t ha-1 observed by Quinchoa et al. (2010), Ñústez (2011), and Pérez (2015) in the same cultivars.
For the fertilized Humic Dystrupets soils (Choconta) (Fer1ch), an increase in yield of 36.4% and 69.9% was observed for Suprema and Capiro, respectively, with respect to the initial soil conditions. These values were lower than the ones found in Typic Hapludands in Subachoque, but with higher increases in FWt, 60.9% (Suprema) and 87.1% (Capiro). This shows the importance of fertilization for both cultivars in low fertility soils (Tab. 1) and agrees with the report by Quinchoa et al. (2010), who found positive responses to fertilization of up to 95% YFt in Capiro in low fertility andisoles in the Antioquia province (Colombia). The lowest response to fertilization was found in Andic Eutrudepts (Facatativa) with 20.1% for Suprema and a marginal and non-significant increase with respect to the control treatment (Fig. 1); in addition, there was a 30.3% yield increase in Capiro, which corroborates a higher nutrient availability in this soil, as compared to the other evaluated soils.
For the two cultivars in the three locations, positive and significant responses in yield were observed, with a better response to fertilization in Capiro than in Suprema since Capiro is better adapted to soils with higher fertility. Capiro had the best response to the higher edaphic K supply, a better base ratio (Ca/K, Ca+Mg/K) (Tab. 1) and a balanced fertilization (Fig. 1). The differential response of the two cultivars, which was greater in Humic Dystrudepts (Choconta), could be explained by the better balance in the edaphic supply of K (0.68 cmol kg-1), with lower P fixation, higher average contents of soil organic matter and absence of interchangeable Al+3 (Tab. 1). This was observed in the Typic Hapludands (Subachoque), which had P fixing soils associated with low mineralization and Al+3, where lower levels of available N might be present. Additionally, deficient K levels were observed below the critical levels (0.1 cmol kg-1) as reported by Castro and Gómez (2013) for similar soils.
The cvs. Suprema and Capiro showed a greater conversion of assimilates in the fertilized Humic Dystrudept soils (Choconta), with 27.03% and 28.2% DWt at harvest, respectively, a higher value than the one reported by Nústez (2011) in the same cultivars (24% DWt). This greater contribution of DWt% favors their industrial use. In contrast, in the high fertility Andic Eutrudepts soils (Facatativa), lower yields and a lower accumulation of assimilates were obtained in the tubers, with 22.01°% DWt and 19.8% DWt in Capiro and Suprema, respectively. The best DWt in Suprema was related to the fertilized soils with lower fertility in Subachoque and Choconta (Figs. 2 and 3), with a total DWt of634 g plant-1 and 740 g plant-1, respectively. The DWt was higher than those reported by Nústez et al. (2009), 450 g plant-1, for this cultivar in low fertility soils in the Colombian potato producing zone of Zipaquira (2,580 m a.s.l.).
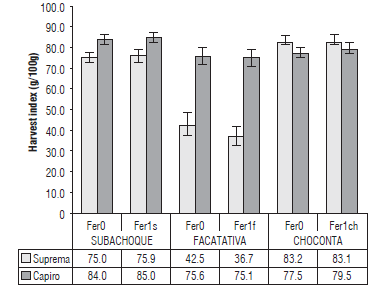
FIGURE 2 Harvest index (HI) in the cvs. Capiro and Suprema in the absence of fertilization Fer0, with respect to balanced fertilization, by location: F1s, Typic Hapludands (Subachoque); F1ch, Humic Dystrudepts (Choconta), and F1f, Andic Eutrudepts (Facatativa). p<0.0l for location*cultivar. Error bars indicate standard errors.
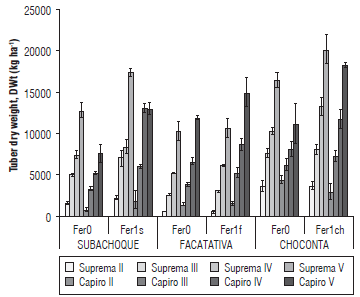
FIGURE 3 Tuber dry weight (DWt) in the cvs. Capiro and Suprema at four phenological stages (II, start of tuberization, III, maximum tuberization-start of filling, iV, filling of tuber, V maximum filling and maturation) in the absence of fertilization F0, with respect to balanced fertilization by location for the soils: Typic Hapludands (Subachoque), F1s; Humic Dystrudepts (Choconta), F1ch and Andic Eutrudepts (Facatativa), Fer1f; p<0.01 for fertilization (location*cultivar). Error bars indicate standard errors.
Capiro, with low to high fertility soils, presented a HI between 75% and 85%, as compared to Suprema, which presented a HI between 75 and 83% in low fertility soils in the locations of Subachoque and Choconta, respectively. Suprema limited its assimilate partition in the high fertility soils of Facatativa with a lower than 45% HI because of its more indeterminate growth habit and a later growth cycle with less accumulation of DWt (21.2%); similar results were obtained by Giletto and Echeverria (2015) for late-cycle and indeterminate-type cultivars, such as Markies Russet.
The lower HI in Suprema was related to the lower adaptation of this genotype because it was cultivated in Facatativa at marginal altitudes close 2,500 m a.s.l. and with average air temperatures higher than 13°C, as compared to the environmental parameters defined by Nüstez (2011). Additionally, there was an availability of soil nutrients (Tab. 1), which may have generated excess N that could have inhibited tuberization and tuber growth, confirming the report by Ruza et al. (2013). On the other hand, Capiro was more adapted to high levels of N and K because of its better ability to partition assimilates to the tuber, as characterized by its genotype, with a determinate habit and better aerial shoots/tubers ratio. This makes it a more efficient crop; coinciding with research conducted by Kleinkopf et al. (1981) and Trehan and Singh (2013) for Group Chilotanum.
The HI in Capiro and Suprema did not show significant differences in response to the fertilization, but did present differences in the cultivar x location interaction (Fig. 2).
The non-significance of HI in response to fertilization is consistent with results obtained by Zelalem et al. (2009) and Burga et al. (2014), who reported that increasing doses of N and K, respectively, had no influence on this parameter. This can be explained by adaptation mechanisms and characteristics of the genotype and also coincides with reports by Rengel and Damon (2008) and Gileto and Echevarria (2015).
As for the accumulation of DWt in Suprema at 125 das (Stage IV) and in Capiro at 100 das (Stage III), an earlier filling of tubers was observed with the Typic hapludands (Subachoque) soil as compared to the other locations. This was probably because these cultivars were grown in soils with lower fertility and a low contribution of K (0.1 cmolc kg-1), as compared to the Choconta and Facatativa soils (Fig. 3, Tab. 1), where tuber filling in both cultivars was concentrated towards the end of the cycle. This could possibly be due to the better availability of nutrients, mainly K, during all of the phenological stages, which presented a gradual and linear extraction of these elements (Fig. 3). The greater DWt accumulation at stages IV and V in the evaluated cultivars could be associated with phenological stages of higher photosynthetic demands, and coincided with stages of high translocation and assimilation partition, verifying the report by Valbuena et al. (2010) for these cultivars.
Efficiency in the nutrient use by tubers
Suprema in the Andic Eutrudepts with high fertility (Facatativa) presented a negative response in the recovery of N (-10.8 RFNt) and K (-9.6 RFKt) for the balanced fertilizer application (Fig. 4) and a negative physiological response of N (-267.9 EPt N) and K (-250 EPt k) for tuber production when compared to positive NUE indices for Capiro (Fig. 5), indicating significant differences between location and cultivar. The highest RFt of N and K for the applied fertilizer was observed for Capiro in Facatativa in high fertility soils in the basin zone, with an RFNt of 70.2% and an RFKt of 79.8%; these indexes were higher than those found in Choconta, with an RFNt of 44.8% and an RFKt of 42.3%, and in Subachoque (38.1°% RFNt and 46.9% RFKt) probably because these soils present greater losses of these nutrients as a result of runoff since they are located in mountainous terrain. Choconta and Subachoque also presented a high yield potential for Capiro and a lower than 60% RFNt, as reported by Vos (2009) for nitrogen applications between 150-200 kg ha-1, and had similar indices for K acquisition, as reported by Gómez and Torres (2012) in the cv. Capiro.
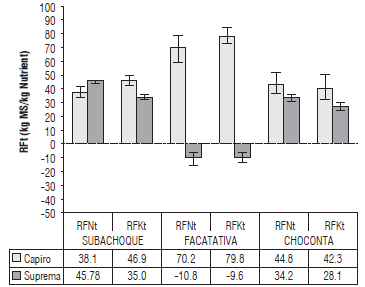
FIGURE 4 Efficiency of recovery of N and K in tubers (RFt), kg nutrient extracted per 100 kg of nutrient applied in balanced fertilization, for the cv. Capiro and cv. Suprema in contrasting soils in the Andean region of Colombia. P<0.01 in N and K for location*cultivar. Error bars indicate standard errors.
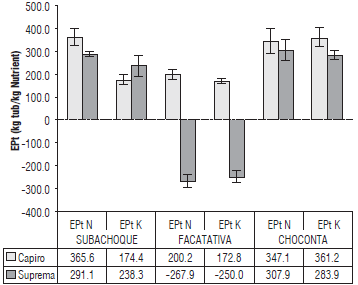
FIGURE 5 EPt of N and K (kg of tuber harvested per kg of nutrient extracted) in Capiro and Suprema in contrasting soils in the Andean region of Colombia. P<0.01 in N and K for location*cultivar. Error bars indicate standard errors.
Suprema presented a low recovery efficiency (<45%) of N (RFNt) and K (RFKt), with lower rates than Capiro for K in the low fertility soils (Fig. 4). This is a characteristic of high yield and indeterminate cultivars, as reported by Kleinkopf et al. (1981) for the cvs. Russet Burbank and Centennial Russet. In addition, the low K efficiency was probably due to a low supply of non-exchangeable K to the roots, with low K extraction in these soils associated with a low root/ shoot ratio. Similar results were reported by Trehan and Singh (2013) for the cultivars Group Chilotanum Kufri Jyoti and Kufri Badshah. Therefore, it is necessary for soils with low K availability to increase the diffusion and mass flow of this element by means of an adequate K contribution and fractionation in the mineral and/or organic fertilizers starting at sowing. This helps counteract the effect of antagonistic elements, such as Al+3, maintaining a balance in the Ca+2 and Mg+2 ratio, and, thus, improving the rhizosphere environment and root growth.
In the Humic Dystrudepts soils with lower fertility in Choconta, where the best yields were obtained, a better physiological efficiency in the production of tubers by nutrient extraction (EPt) was observed, with significant differences in the interaction between the location and the cultivar (Fig. 5). Under these conditions, Capiro and Suprema produced an EPtN of 347 and 308 kg FWt kg-1 N, respectively, and an EPtK of 361 and 283 kg FWt kg-1 K. These results were higher for N and similar for K when compared to those reported by Trehan and Singh (2013), with an EPtN between 250 and 318 kg FWt kg-1 N and an EPtK between 256-360 kg FWt kg-1 K for the efficient Group Chilotanum cultivar Kufri and the hybrid JX 576.
This is associated with the higher response to fertilization in FWt and DWt in Capiro and Suprema in this location. It also coincides with the greater use efficiency of N and K in the tubers (Fig. 6), which can be explained by the better partitioning and conversion of N, K and assimilates to the tubers with a better sink strength of the tubers during filling for both cultivars under the edaphic-environmental conditions of this location. In addition, the greater efficiency in the cv. Capiro in contrasting soils coincides with the better adaptation of this cultivar to a wider range of altitudes (1,800-3,200 m) and soils, similar to the optimal environmental conditions reported by Ñústez (2011).
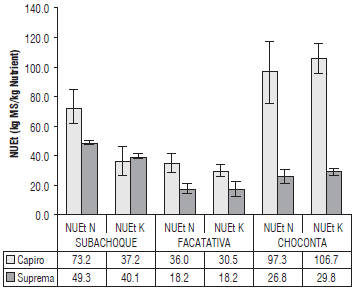
FIGURE 6 Efficient use of N and K in tubers with balanced fertilization, NUEt (kg of dry matter of the tuber per kg of nutrient extracted) in the cvs. Capiro and Suprema in Typic Hapludands, Subachoque, Andic Eutrudepts, Facatativa, and Humic Dystrudepts, Choconta in the Andean region of Colombia. Error bars indicate standard errors.
Capiro responded better to the fertile soils, with HI>75% (Fig. 2). Lower soil loss factors were observed as a result of the location in lacustrine basin areas and the high native K and N contents (Tab. 1). Again, this confirms that the best adaptation for Capiro is in flat areas and in the high fertility soils of the Bogota plateau. This is seen in a better efficiency in the translocation of nutrients, which are assimilated to the tuber, and a high removal of N and K from the plant (Fig. 4), which needs to be replenished with fertilization plans. The better adaptation of this cultivar can be explained by possible differential absorption mechanisms with a greater flow of K and N to the root and tuber. In addition, the presence of specialized channels that also favor assimilate translocation, together with a lower aerial shoots/tuber ratio, represent mechanisms that have been explained for K by Trehan and Singh (2013) and Wang and Wu (2015), and for N by Vos (2009), who verified the differences between the genotypes.
Under the conditions of high K availability (3.14 cmolc kg-1) in the Andic Eutrudepts (Facatativa), Suprema responded negatively to the fertilization of this nutrient with an EPtK of -250 and presented the lowest EptK, 172.8 for Capiro (Fig. 5). Additionally, the tuber production response of Suprema was marginal for the high potassium saturation soils (>9%) in the Andic Eutrudepts (Facatativa), as shown in Figure 1.
These data suggest a possible luxury K uptake for Suprema and a marginal uptake for Capiro, phenomena that have been explained for the Group Chilotanum by Kang et al. (2014). On the other hand, Karam et al. (2009) found less efficiency in the use of K with K2O levels higher than 289 kg ha-1 for the cvs. Derby and Umatilla Russet. In addition, Burga et al. (2014) verified that, in tetraploid cultivars, high levels of K affected the development of tubers, to the detriment of yield, where excesses of this element might limit the transport of other assimilates or hormones.
On the other hand, the Facatativa soils had the lowest physiological efficiency of N (NUEt of 18 kg kg-1 for Capiro and 36 kg kg-1 for Suprema) and K (NUEt of 36 kg kg-1 Suprema and 30 kg kg-1 Capiro) with contributions of 171 kg ha-1 of N and 180 kg ha-1 of K, respectively, with high N and K availability (Tab. 2). This agrees with the findings of Zebarth et al. (2012) and Saravia et al. (2016), who found low NUE values in potatoes, 40 and 10 kg kg-1, respectively; increasing the availability of N in crops with doses between 200 and 300 kg ha-1 of N can help with positive plant responses.
The highest physiological efficiency in the use of nutrients in the tubers was obtained in Capiro, with an NUEt of 97.3 kg kg-1 for N and 106.7 kg kg-1 for K in the fertilized soils with lower fertility in Choconta. This corroborates the better efficiency of Capiro, a characteristic of this genotype that has already been mentioned, and verifies that the best NUE was significantly different from that in Suprema. Capiro appears to be a genotype of a determinate type, in agreement with results of Kleinkopf et al. (1981) for cultivars of a similar type. These indices, found in the soils with lower fertility, were higher than those reported by Poljak et al. (2011), with a NUEt of 71 to 76 kg kg1 for N in the Group Chilotanum with an N fertilizer contribution between 150-200 kg ha-1. This was probably due to the positive interaction of the nutrients in the balanced fertilization of macro and micronutrients in unsaturated soils, with an improvement in the efficient use of nutrients in tropical soils, which confirms the observations of Pro-chow et al. (2009).
The low physiological efficiency of N and K in the tubers for the cv. Suprema in the soils with excesses of N might suggest a luxury consumption of these elements that could be due to a lower growth of tubers resulting from the low transport of assimilates to organs associated with low HI (Fig. 2). With a higher average temperature than in the other locations, this could result in a high aerial shoots/ tubers ratio, which limits the flow of carbon and nutrients to tubers. These results are similar to those reported by Fandika et al. (2012) and Saravia et al. (2016) for N and Wang and Wu (2015) for K. These authors suggested agronomic and genetic strategies to decrease the air shoot/ tuber ratio and improve the translocation of assimilates to the tubers. According to Roumeliotis (2012), excess N and high temperatures limit tuber formation and growth. Fandika et al. (2012) also reported a less efficient use of N with lower partitioning of assimilates to the tubers and a low number of physiological sources at the start of tuberization under high N doses.
An imbalance resulting from excess available N in a soil during tuber filling at the start of tuberization could also cause a reversal of tubers into stolons, affecting the productive potential, a phenomenon that should be evaluated in these cultivars in future research. Similar results were proposed by Güller (2009) for the potato Group Chilotanum cv. Ana, where it was found that, at doses higher than 200 kg ha-1, a smaller number of tubers were present at the tuberization stage, with a higher generation of source structures. In addition, Zelalem et al. (2009) reported that doses higher than 140 kg N ha-1 increased the number of stems and delayed flowering, which, according to Roumeliotis (2012), affected the synthesis and transport of the FT-like protein tuberigen, which favors tuberization in this species.
The lower physiological indices found in the use of K obtained for Suprema in high fertility soils (Figs. 4-6) coincided with a rapid vegetative growth, later tuberization and less growth of the source organ. This agrees with the lower accumulation of K and dry matter and the presence of smaller tubers despite the presence of the same number of tubers as in Capiro. This can be explained by the excess K, which can generate an imbalance by allowing nitrate accumulation with a lower assimilation in the aerial part and a decrease in the transport of carbohydrates and proteins towards the tubers. Similar effects from excess K were found by Kang et al. (2014).
Conclusions
Capiro is more efficient in the use of N and K than Suprema regardless of the soil type and location, although Suprema presented the best responses and physiological indices in the soils with lower fertility at the higher altitude and with a lower ambient temperature in response to the balanced fertilization. This suggests the use of integrated fertilization management aimed at improving the N and K availability in the rhizosphere (acidity and nutrient balance) and a specific fertilization in both cultivars, considering the environmental supply per site and the soil supply given by the soil pedogenesis.


