











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Physalis peruviana L. is a plant of Andean origin cultivated in South America, from Venezuela to Chile. Nowadays, it is grown in other parts of the world, such as California, South Africa, India, New Zealand, Australia and Great Britain (Ramadan and Morsel, 2003; El-Tohamy et al., 2012). P. peruviana L. fruits are round, bright orange and have a pleasant taste related to their tropical origin (Fischer et al., 2007). This species is very important due to its high nutritional value and potential health benefits, derived from the high content of ascorbic acid, vitamins and antioxidants (Puente et al., 2011; Briones-Labarca et al., 2013). The high prices paid for cape gooseberries in local and international markets have made this plant attractive to producers (Fischer et al., 2007; Ramírez et al., 2013).
One of the main limiting factors of agricultural production, especially in the tropics and subtropics, is the phosphorus (P) deficiency (Sánchez, 1976; Ramaekers et al., 2010). This element has an important role in various processes, such as nucleic acid synthesis, energy production, photosynthesis, respiration, synthesis and stability of the membrane, enzyme activation and signaling (Vance et al., 2003; Hawkesford et al, 2012; Manschadi et al, 2014). It has been reported that P deficiency reduces the production of biomass in leaves, affecting plant growth (Kirschbaum and Tompkins, 1990; Plénet et al, 2000; De Groot et al., 2001; De Groot et al., 2003; Chaudhary et al., 2008; Maathuis, 2009; Reich et al., 2009). A change in the root/shoot ratio due to an increase in the density of root hairs, an adaptive response of plants to P deficiency, has also been reported (Mollier and Pellerin, 1999; Hermans et al., 2006; Yao et al, 2007; Zhang et al., 2013).
The decrease in CO2 assimilation due to P deficiency is mainly associated to a decline in the regeneration of Rubisco (Jacob and Lawlor, 1992; Rao and Terry, 1995; Campbell and Sage, 2006; Singh et al., 2013) and stomatal closure caused by the accumulation of CO2 in intercellular spaces (Thomas et al., 2006). This effect has also been observed in corn (Tewari et al., 2004) and rice (Guo et al., 2012). Phosphorus deficiency decreases the chlorophyll content, thus affecting photosynthesis. The imbalance in photosynthesis phases generated by P deficiency can cause an increase of reactive oxygen species (ROS), which alter plant functions by damaging lipids, proteins, enzymes, nucleic acids and photosynthetic pigments. Moreover, an increase in free radical production has been reported in bean (Phaseolus vulgaris L.) roots in plants with phosphate deficiency (Malusa et al., 2002). ROS damage cell membranes, causing changes in permeability and resulting in electrolyte leakage (EL) and alterations in cellular metabolism (Blokhina et al., 2003).
In order to reduce oxidative damage, plants have developed different response mechanisms, especially the production of antioxidants and protective molecules (Cruz de Carvalho, 2008). P deficiency in maize (Tewari et al., 2004) and rice (Guo et al., 2012) has been observed to cause a significant increase in the activity of antioxidant enzymes, such as catalase (CAT), peroxidase (POD) and superoxide dismutase (SOD), and an increase in the content of proline (Pro). In tomato plants, which are tolerant to low P availability, increased peroxidase activity and anthocyanin accumulation have been observed (Khavari-Nejad et al., 2009). Also, an increase in the proline content of roots and stems (Sarker and Karmoker, 2011) has been found in lentil plants (Lens culinaris Medik).
P. peruviana L. is grown from 1500 m to 3000 m a.s.l. in the Andes, often in volcanic soil with an acidic pH and low phosphorus availability due to the high binding capacity of phosphate anions in the soil solution (Fischer et al., 2007; Ramirez et al., 2013). These P limitations affect both the yield and quality of cape gooseberry plants (Garzón-Acosta et al., 2014). However, there have not been any studies that describe the effects of phosphorus deficiency on the physiology of the plant at different stages of development. The purpose of this research was to determine the response mechanisms of cape gooseberry (P. peruviana L.) seedlings in the early stages of development under conditions of phosphorus deficiency through the analysis of morphological, physiological and biochemical responses.
Materials and methods
Plant material and experimental design
Cape gooseberry (P. peruviana L.) seeds ecotype Colombia were germinated in peat moss in a growth chamber with a photosynthetic photon flux density (PPFD) of 350 μπω! m-2 s-1, 70% relative humidity, with 12 h of light at 22°C and 12 h of darkness at 10°C. Twenty days after germination, the seedlings were transplanted to 50-cell trays with an inert substrate (2:1:1 mixture of river sand, quartz sand and fine granite). A 0.8 strength Hoagland solution and the designated P doses were applied. The seedlings grew in a greenhouse with a PPFD range between 450-900 μmol m-2 s-1 PAR (photosynthetic active radiation), 70% relative humidity, 12 h of light at 25°C and 12 h of darkness at 8°C. A completely randomized block design with five treatments, four replications and an experimental unit of eight plants was used. The treatments had five levels of P2O5 kg-1 in the substrate: 0 mg P2O5 kg-1 (P0), 6 mg P2O5 kg-1 (P6), 12 mg P2O5 kg-1 (P12), 25 mg P2O5 kg-1 (P25) and 50 mg P2O5 kg-1 (P50). The variables were measured at 15 d of treatment (dt), 30 dt, 60 dt and 90 dt.
Growth parameters
The plants were individually divided into roots, stems and leaves and the number of leaves (NL) and the leaf area (LA) were measured using a portable area meter (CI-202, CID Inc, USA). Then, the plant material was dried at 70°C until a constant weight was achieved. The root/shoot ratio was determined using these data and the root length density (RLD) was determined according to Newman and Ritz (1986).
Gas exchange
The photosynthetic rate (A), stomatal conductance (gs) and transpiration (E) were registered at 90 dt using a photosynthesis measurement system (LCpro-SD, Portable BioScientific, UK). The measurements were taken from the fourth apical leaf on six plants per treatment, between 9:00 AM and 11:00 AM, with a CO2 concentration of 360 μl L-1 and a PPDF of 900 μmol m-2 s-1.
Chlorophyll content and leaf temperature
The pigments were extracted according to Cubas et al. (2008). The apical expanded leaves from six plants per treatment where homogenized in 80% acetone. The absorbance was determined at an optical density (OD) of 663 nm and 647 nm. The leaf temperature (LT) was determined with an infrared thermometer (HD550, Extech®, USA) on the fourth apical leaf.
Membrane permeability
The membranes permeability was measured by electrolyte leakage (EL) according to Rodríguez et al. (2005). Eight 2.5 mm diameter leaf discs were placed in Falcon tubes with 2 ml of deionized water at 24°C. The electric conductivity (EC) was determined with an electrical conductivity meter (HI 9835 HANA, USA) at 24 h. The EC values are expressed as a percentage with respect to the highest value using the following equation:
PE = (EC1*EC2-1)*100 (1)
where: PE = % of lost electrolytes, EC1 = electric conductively at 24 h, and EC2 = electric conductivity after heating to 80°C. The measurements were carried out on the fourth apical leaf.
Antioxidant enzymes, proline and protein content
A 0.2 g powdered leaf sample was treated with polyvinylpyrrolidone 40 (22% w/w) and with 1.5 ml cold 80% (v/v) acetone and centrifuged (8000xg for 30 min, 4°C) to eliminate pigments, according to Lichtenthaler (1987). Afterwards, samples were extracted in 3 ml of 110 mM sodium phosphate buffer (pH 7.2) containing 3.82% (w/v) polyvinyl pyrrolidone 40. The homogenate was centrifuged (6000 x g, 1 h, 4°C) and the supernatant was used for the subsequent enzyme assays. The CAT activity was determined by the method of reduction of potassium permanganate (KMnO4) with hydrogen peroxide in sulfuric acid (Ulrich, 1974). The POD activity was estimated by the absorbance change at 436 nm caused by the oxidation of guaiacol according to Kireyko et al. (2006). The proline level was determined according to Bates et al. (1973) based on the prolineninhydrin reaction. For the proline (Pro) determination, a 1:1:1 solution of proline, ninhydrin and glacial acetic was incubated at 97°C for 1 h. The reaction was stopped by placing the vials in an ice bath; the chromophore was extracted with toluene and its absorbance was measured at 520 nm. The soluble proteins were measured based on the method of Bradford (1976), using bovine serum albumin as a reference.
Results and discussion
Growth parameters
The seedling dry weight decreased by reducing the concentration of P in the nutrient solution (Fig. 1A). This decrease was significant between P0 and P6 and the other treatments. The highest values for the dry weight were observed in P50 at 30 dt (0.60 g), 60 dt (4.63 g) and 90 dt (18.13 g) and the lowest values were observed in P0 at 30 dt (0.07 g) and 60 dt (0.08 g); the P0 plants died before 90 dt. Although there were differences between P12, P25 and P50 at 90 dt, the values were closer to the values of 30 dt and 60 dt. The assimilate partitioning in P0 and P6 was higher towards the root, compared to the other treatments at 30 dt and 60 dt (P0: 0.038 g, 0.047 g and P6: 0.043 g, 0.064 g, respectively). At 90 dt, there were differences in the partition of assimilates to the root between P12 and P25 as compared to P50, where fewer assimilates were partitioned to roots (Fig. 1B).
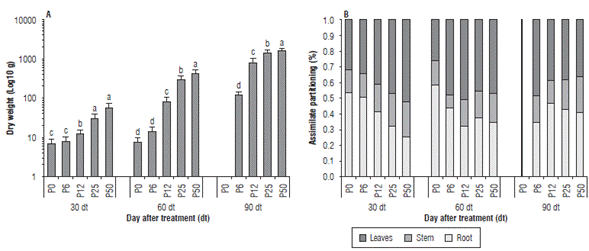
FIGURE 1 Effect of five phosphorous levels in the soil: 0 (P0), 6 (P6), 12 (P12), 25 (P25) and 50 (P50) mg of P2O5 kg1 on the seedling dry weight, logarithmically transformed data (A) and assimilate partitioning (B) of the cape gooseberry seedlings (Physalis peruviana L.) at 30, 60 and 90 d of treatment (dt). The data shown are the averages of sixteen replicates, with the standard deviations indicated by the vertical bars. Means denoted by the same letter do not significantly differ at P<0.05 according to the Tukey test.
The root/shoot ratio was higher at 30 dt and 60 dt for P0 (1.1; 1.5; nd) and P6 (1.0; 0.8; 0.7) compared to the others treatments. P12 and P25 did not show differences in the root/shoot ratio between each other, but were higher than P50 (Tab. 1). The NL, LA and RLD progressively decreased with a reduction in the concentration of phosphorous in the nutrient solution (Tab. 1). At 90 dt, no differences were observed in the NL and RLD between P25 and P50, but the LA was lower in P25 (168.3 cm2) than in P50 (196 cm2). Phosphorus is an essential element for the growth and development of plants; however, there are differences in requirements due to the efficient use some plants make of this element (Kirschbaum and Tompkins, 1990; Nielsen et al., 2001; Chaudhary et al., 2008; Maathuis, 2009; Reich et al., 2009). In the cape gooseberry seedlings, there was a decrease in the dry weight, number of leaves and leaf area as a result of a decrease in the dose of phosphorus; similar data have been reported for different species (Radin and Eidenbock, 1984; Guo et al., 2012). The seedlings in the P0 treatment, without phosphorus, achieved little growth, possibly by using seed reserves, and did not survive beyond 60 dt. In addition to the reduction of total biomass in plants, one of the indicators of phosphorus deficiency is an increase in the partitioning of photoassimilates to roots (Mollier and Pellerin, 1999; Hermans et al., 2006; Yao et al., 2007; Zhang et al., 2013). It was observed that all of the treatments, except P50, had a higher proportion of biomass in the roots and a higher root/shoot ratio. This response has been described for many plants suffering P deficiency as a strategy to get through the depleted zone in the substrate and obtain more P (Mollier and Pellerin, 1999; Hermans et al., 2006; Yao et al, 2007). These data suggest that the cape gooseberry seedlings in P6, P12 and P25 had a P deficiency. Likewise, the decrease in the difference in the dry weight of the seedlings at 90 dt between P12, P25 and P50 suggests that the seedlings changed their metabolism to increase the efficiency of P use, as has been reported in many plants (Nakamura, 2013).
TABLE 1 Effect of five phosphorous levels in the soil: 0 (P0), 6 (P6), 12 (P12), 25 (P25) and 50 (P50) mg of P2O5 kg1 on the Number of Leaves per plant (NL), Leaf area (LA), Root Length Density (RLD) and Root Shoot ratio (Root/Shoot) in cape gooseberry (Physalis peruviana L.) seedlings at 30, 60 and 90 d after treatment (dt). The data shown are the averages of sixteen replicates. Pi, Phosphorous levels; nd, data not available.
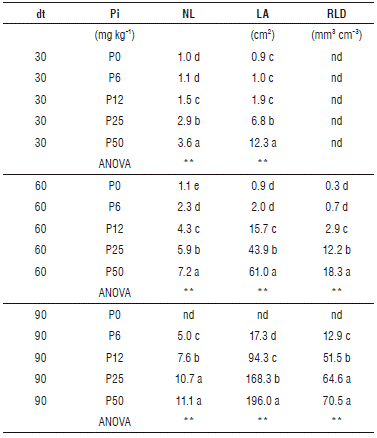
Means denoted by the same letter do not significantly differ at P<0.05 according to the Tukey test. Comparisons among treatments were analyzed by one-way ANOVA. Statistical significance for P<0.05 (*), P<0.01 (**).
Gas exchange and leaf temperature
The P25 and P50 treatments had higher A values (10 μmol m-2s-1; 10.54 μmol m-2s-1) as compared to P12 and P6 (2.76 μmol m-2s-1; 1.67 μmol m-2s-1) (Fig. 2A). E behaved similarly to A, with differences between P6 (0.26 mmol m-2s-1) and P50 (2.81 mmol m-2s-1) (Fig. 2B). P25 had the highest gs value (0.202 mmol m-2s-1) and P6 and P12 had lower values (0.025 mmol m-2s-1; 0.100 mmol m-2s-1) (Fig. 2C).
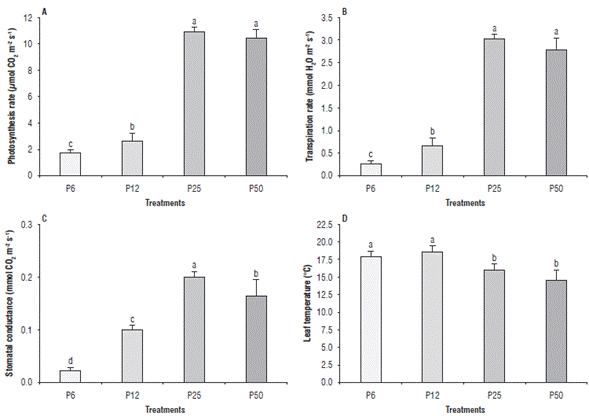
FIGURE 2 Effect of four phosphorous levels in the soil: 6 (P6), 12 (P12), 25 (P25) and 50 (P50) mg of P2O5 kg1 on the gas exchange. A, photosynthesis rate (A), B, transpiration rate (E), C, stomatal conductance (gs) and D, leaf temperature (LT) in cape gooseberry (Physalis peruviana L.) seedlings at 90 d of treatment (dt). The data shown are the averages of sixteen replicates, with the standard deviations indicated by the vertical bars. Means denoted by the same letter do not significantly differ at P<0.05 according to the Tukey test.
The leaf temperature was higher in P0 (18.1°C) and P12 (16.1°C) compared to P25 (16.1°C) and P50 (14.8°C) (Fig. 2D). The temperature increased between 3.3°C and 3.9°C between the treatments with deficient P (P0 and P6) and the treatment with sufficient P (P50). Plant biomass production depends directly on net photosynthesis, which is related to the plant ability to intercept light and the efficiency of photosynthetic metabolism (Singh et al., 2013). Large decreases in A and gs were observed in P6 and P12, as compared to P50, possibly due to the decreased synthesis of ATP in the photosynthesis phase, which causes a limitation in the regeneration of the CO2 acceptor ribulose-1,5-bisphosphate in the Calvin cycle. This slows carboxylation and increases the intercellular concentration of CO2, which causes stomatal closure (Singh et al., 2013). Similar results have been reported in other species at P deficit conditions, such as Arabidopsis, sorghum, corn, beans and cotton (Radin and Eidenbock, 1984; Barrett and Gifford, 1995; Abel et al., 2002; Tewari et al., 2004; Yao et al., 2007; Chaudhary et al., 2008; Singh et al., 2013). Similarly, the decrease in gs caused by an imbalance in the two phases of photosynthesis produced a reduction in E and an increase in LT due to a decrease in energy dissipation. Although the seedlings in P25 had a higher root/shoot ratio than P50 and a decrease in dry weight accumulation, which indicates a deficiency of P, they did not show a decrease in A. This suggests that these plants exhibited a slight deficiency that affected cell division, but not the function of the photosynthetic apparatus.
Chlorophyll content
The chlorophyll content (Chl) decreased in P6 (1343 μg g-1 DW) and P12 (1330 μg g-1 DW) as compared to P25 (1615 μg g-1 DW) and P50 (1568 μg g-1 DW). The reduction in Chl in P6 compared to P50 was 14.3% (Fig. 3). A decrease in chlorophyll content due to P deficiency has been reported in plants such as maize (Tewari et al., 2004). In the case of Solanaceae, such as tomatoes, a severe reduction has been reported in the chlorophyll content with combined N and P deficiency (Khavari-Nejad et al., 2009). The reduction in the chlorophyll content can also be related to the degradation of these molecules by ROS (Misson et al., 2005). The decrease in the A may also have been related to the decrease in the chlorophyll content observed in P6 and P12.
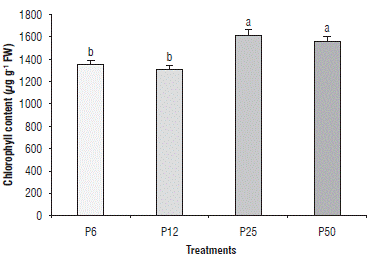
FIGURE 3 Effect of four phosphorous levels in the soil: 6 (P6), 12 (P12), 25 (P25) and 50 (P50) mg of P2O5 kg-1 on the chlorophyll content in cape gooseberry (Physalis peruviana L.) seedlings at 90 d of treatment (dt). The data shown are the averages of sixteen replicates, with the standard deviations indicated by the vertical bars. Means denoted by the same letter do not significantly differ at P<0.05 according to the Tukey test.
Membrane permeability
The P0, P6 and P12 treatments had higher percentages of EL (Fig. 4). The EL for P6 at 30 dt, 60 dt and 90 dt was 44%, 40% and 37%, respectively, while for P50 it was 19%, 30% and 19%, respectively (Fig. 4). The P25 and P50 treatments showed no significant differences between each other at 60 and 90 dt. Due to the imbalance in the two phases of photosynthesis caused by ATP deficiency, there is an increase in ROS production, which caused damage to macromolecules and structures as membranes (Misson et al., 2005). Here, there was an increase in EL in P0, P6 and P12, suggesting that P deficiency causes damage to membranes, affecting the permeability degree, probably due to lipid peroxidation induced by ROS.
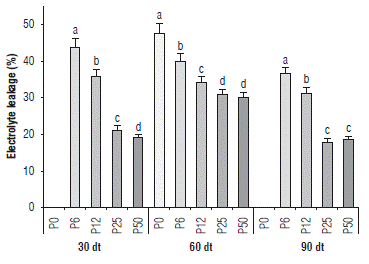
FIGURE 4 Effect of five phosphorous levels in the soil: 0 (P0), 6 (P6), 12 (P12), 25 (P25) and 50 (P50) mg of P2O5 kg-1 on electrolyte leakage in cape gooseberry (Physalis peruviana L.) seedlings at 30, 60 and 90 d of treatment (dt). The data shown are the averages of sixteen replicates, with the standard deviations indicated by the vertical bars. Means denoted by the same letter do not significantly differ at P<0.05 according to the Tukey test.
Antioxidant enzymes, proline and protein content
The activity of the antioxidant enzyme CAT was determined in the leaves at 15 dt, 30 dt, 60 dt and 90 dt (Tab. 2). The CAT activity was higher for all of the treatments at 15 dt, when the activity in P0 (791 UA g-1 FW) was approximately 3.3 times higher than P50 (238 UA g-1 FW). The CAT activity gradually decreased over time in all of the treatments and, at 90 dt, presented lower values. Throughout the experiment, P50 had the lowest values of CAT activity (106.2 UA g-1 FW to 238.3 UA g-1 FW) (Tab. 2). The POD activity was low in all of the treatments, but had higher values in P6 (1.4 mg POD g-1 FW) and P12 (1.38 g-1 FW) at 15 dt. The POD activity gradually reduced until 90 dt, when the lowest value was observed for all treatments.
TABLE 2 Effect of five phosphorous levels in the soil: 0 (P0), 6 (P6), 12 (P12), 25 (P25) and 50 (P50) mg of P2O5 kg-1 on the content of Catalase (CAT), Peroxidase (POD), Proline (Pro) and total protein in cape gooseberry seedlings (Physalis peruviana L.) at 15 dt, 30 dt, 60 dt and 90 dt. Pi, Phosphorous level; FW, fresh weight; nd: data not available.
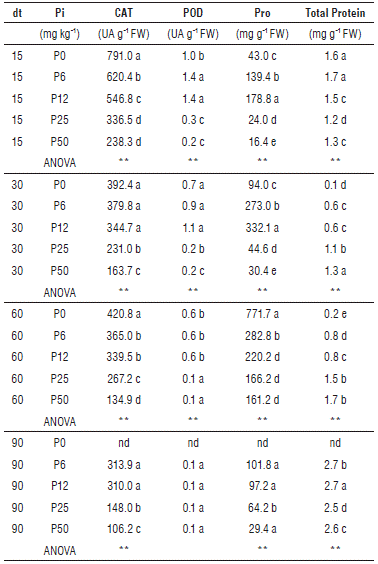
Means denoted by the same letter do not significantly differ at P<0.05 according to the Tukey test. Comparisons among treatments were analyzed by one-way ANOVA. Statistical significance at P<0.05 (*), P<0.01 (**).
The contents of Pro throughout the experiment were higher in P0, P6 and P12 (Tab. 2). At 60 dt, P0 showed the highest value for the Pro content (771.7 mg g-1 FW), just before the plants died. At 90 dt, the Pro content in all of the treatments decreased between 101.8 mg g-1 FW (P6) and 29.4 mg g-1 FW (P50). The protein content was higher in all of the treatments at 90 dt, when the Pro content was lower (Tab. 2), with values between 2.5 mg g-1 FW (P25) and 2.7 mg g-1 FW (P6, P12). Higher antioxidant enzyme activity, such as peroxidase (POD) and superoxide dismutase (SOD), has been reported in rice seedlings (Guo et al., 2012) and also in maize, with increase in other enzymes as CAT and ascorbate peroxidase (APX) under P deficiency (Tewari et al., 2004). Additionally, in beans, P deficiency increases CAT and POD activities (Juszczuk et al., 2001). In cape gooseberry seedlings, there was an increased CAT activity for the P0, P6, P12 and P25 treatments at 15 dt (Tab. 2). The increase was greater for the treatments with lower doses of P, showing that the expression of this enzyme is an early response in cape gooseberry seedlings to a deficiency of this element and may be related to the observed decrease in EL. Similarly, POD enzyme activity was low, but was higher in treatments with lower doses of P. It has also been reported that P deficiency can increase the Pro content in roots and stems in lentil plants (Sarker et al., 2011) and in leaves and roots in beans that are deficient in P (Juszczuk et al., 2001). The observed Pro increase in P0, P6 and P12, which was higher at 30 dt and 60 dt, could be another response of cape gooseberry seedlings to reduce ROS caused by an imbalance in photosynthesis (Hare et al., 1998). It has been reported that low molecular weight metabolites, such as Pro, are efficient at detoxifying the OH radical, and increased Pro synthesis has been reported in response to different abiotic stresses or by combining phosphorus deficiency simultaneously with other stresses, such as drought stress (Al-Karaki et al., 1996) or salinity stress (Zribi et al., 2015). Pro accumulation has also been reported in response to stress nutrients such as Al and Ca in beans (Yang and Chen, 2001), or Al and P (Ismail, 2005; Guo et al., 2012) and Fe (Arias-Baldrich et al., 2015), but not under conditions of phosphorus deficiency alone. The observed Pro accumulation may have been a stress response to P deficiency to protect macromolecules and structures from damage caused by ROS, due to its function as a compatible osmolyte (Schobert and Tschesche, 1978). The increase in the defense and protective mechanisms in cape gooseberry seedlings was probably due to metabolic adjustment, which aims at reducing the effect of stress under moderate P deficiency, as noted here. The development of mechanisms such as increased antioxidant capacity and the synthesis of osmolytes has an energy cost that plants probably cannot assume with a severe phosphorus deficiency, as in P0 treatment, because the limitation imposed by deficiency is too high. Furthermore, the metabolic adjustment induced by P deficiency includes other mechanisms, such as the use of PPi-dependent enzymes in the glycolysis pathway and the replacement of membrane phospholipids with sulfolipids; plants have developed these mechanisms since P is a limiting element on the planet (Nakamura, 2013). These stress defense mechanisms, described here for the first time for P. peruviana L., may explain the acclimatization capability observed in cape gooseberry seedlings under moderate P deficiency.


