











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
High temperatures cause considerable changes in plant morphology, cellular dynamism and hormonal relationships, which modify growth patterns (Argentel et al., 2017), creating heat stress when plants do not overcome the minimum tolerance index (Mahalingam, 2015). A single heat shock during growth may cause irreversible damage to plant physiology and yield (Valluru et al., 2017; Garatuza et al., 2018). Many species, such as wheat, need cold hours (also called heat units) to complete critical phenophases (Ganem et al., 2014); for example, at least 300 cold hours are needed for good tillering. The tillering capacity is directly correlated with yield; however, both depend on temperature (Argentel et al., 2017). The tillering phenophase is hormonally regulated by the relationship between abscisic (ABA) and gibberellic (AG) acids, with temperature variations being the abiotic factor that contributes most to hormonal imbalances (Xie et al., 2016). Currently, in some high producing regions, such as Yaqui Valley, Sonora, Mexico, where more than 39% of the domestic wheat production occurs, significant changes in temperature patterns are viewed as evidence of climate change (Lares et al., 2016). This phenomenon, according to parametrized models, will limit the tillering capacity and, consequently, modify the genetic-productive expression potential of wheat (Argentel et al., 2017). For this reason, the present study aimed to evaluate the effect of temperature variability on the tillering capacity and grain yield in eight representative sites in Yaqui Valley during two crop cycles, comparing them to that obtained with an experimental warming of +2°C in the ambient canopy temperature (Garatuza et al., 2018) using the CIRNO C2008 wheat variety.
Material and methods
Experiment area location
The experiment was carried out during the growing seasons of 2016-2017 and 2017-2018 (December-May) under field conditions using a calculus surface of one ha in seven representative sites in Yaqui Valley, Sonora, Mexico, where agro-meteorological stations belonging to the INIFAP (Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias) are located (Fig. 1). These stations record the data of agroclimatic variables, mainly the temperature of the region.
FIGURE 1 Distribution of experimental sites and their respective meteorological stations in Yaqui Valley.
To simulate a site with the maximum temperature (+2°C) predicted for this region, the canopy temperature was increased +2°C with respect to the ambient canopy temperature, at one site in order to evaluate the effect of global warming on the tillering capacity, yield components, and hormonal concentrations during the tillering phenophase. The experimental warming was carried out at the Experimental Technology Transfer Center (CETT-910) of the Instituto Tecnologico de Sonora (ITSON) located in Yaqui Valley at: 27°22'0.4'' N and 109°54'50.6'' W (UTM: 607393.24 m E; 3027508.34 m N), which is a representative place for wheat crops in Yaqui Valley.
Temperature control
The temperature database was supplied by seven climatological stations located in the experiment sites. The digital memory of the meteorological station records readings every 10 min and provides integrated data by hour and day, averaging the temperature and calculating the thermal sum (heat hours, cold hours) with the maximum and minimum threshold temperature of 10°C and 0°C, respectively, following the methodology of thermal sums of Confalone and Navarro (1999). These data are available at http://www.siafeson.com/remas/index.php.
For the maximum predicted temperature value for the region (warming), a T-FACE system was used (Kimball, 2015), which consisted of raising the crop canopy temperature by 2°C using six thermal radiators per plot (FTE-1000 model, 1000W, 240 V, 245 mm long x 60 mm wide, built by Mor Electric Company Heating Association Inc. Comstock Park, MI, USA) on eight equilateral triangular structures, with sides of 5.2 m. Two radiators were installed on each side of the triangular structures, forming a hexagon, which raised the temperature in a 3 m diameter circle for each plot. To control the temperature, infrared temperature sensors (IRTS Apogee Instruments Inc., Logan, UT, USA) were pointed to each plot at an inclination degree of 45° from the soil horizontal surface, covering a circle with a r = 1.5 m at the center of each plot (Garatuza et al., 2018). The IRTS were registered in a datalogger (CR1000 Campbell Sci, Inc. Logan, UT, USA), which sent a voltage signal to an interface (MAI-05V, Avatar Instruments).
Variety, seeding and crop management
In all sites, the CIRNO C2008 wheat variety was used. This variety is classified as crystalline or hard wheat. It originated from selection in segregating populations of SOOTY-9 / RASCON-37 // CAMAYO crossbreed, carried out at the International Center of Maize and Wheat Improvement (CIMMYT). This variety was released for cropping in 2008 and is widely used in Mexico, particularly in the northwest. The grain yield has reached 5.6 t ha-1 and 6.3 t ha-1 with two and three irrigations, respectively, at an irrigation rate of 14 cm when this variety was released in Sonora. This variety, after eight years of being released for crops in southern Sonora, has shown a high yield genetic stability (Argentel et al., 2018).
The seeding was done with a sowing machine (SUB-24) on 8-14 December 2016 and 7-14 December 2016 on vertisol soils (Bockheim et al., 2014) with a seeding density of 170 kg ha-1. The background fertilization, in both experiment crop cycles, was done with a standard of 250 kg ha-1 of Urea + 100 kg ha-1 of monoammonium phosphate fertilizer (MAP, 11-52-00). Three irrigation treatments were applied during the crop cycle. Nitrogen fertilizer was also applied at a dose of 50 kg ha-1 of Urea during the first and second irrigation treatments (growth and tillering phenophases). All irrigation treatments were applied with an average water depth of 14 cm and an irrigation interval of 25 d.
During both crop cycles, the presence of the foliage aphid (Schizaphis graminum) was observed, and the Muralla Max® pesticide (a.i. Imidacloprid + Betaciflutrin) was applied at a rate of 0.20 L ha-1 in the tillering phenophase. Also, a slight presence of broadleaf weeds was observed, which were controlled manually before the irrigation treatments.
Evaluated variables
The tillering capacity was evaluated according to the number of tillers around a principal plant. The final tiller count was done at 45 d after germination.
ABA and GA concentration at the site of maximum temperature (Warming treatment)
For the ABA and GA determination at 15 and 30 d after germination, the methodology of Ortiz et al. (2001) was used. Leaf tissue (10 g fresh weight) was collected and immediately frozen in liquid nitrogen. The frozen leaf tissue was freeze-dried for 48 h, ground and extracted in distilled deionized water with an extraction ratio of 1:40 (dry weight: mL water) at 4°C. The ABA and GA concentrations of the extract were determined using high performance liquid chromatography (Wang et al., 2016).
Grain yield
The grain yield (t ha-1) was determined at each experiment site based on a square meter using 16 replicates.
Statistical analysis
For the tillering capacity and grain yield variables, the mean and standard deviation of each treatment (experiment sites) were determined. The differences between the sites were detected with analysis of variance based on a randomized effect linear model (Fisher, 1937). Additionally, a Tukey post-hoc test for P<0.05 and P<0.01 was applied (Tukey, 1960). To analyze the temperature effect on the grain yield, simple regression was used. The hormonal concentration was compared using a parametric theoretical distribution for t-student quantitative continuous variables (Gosset, 1917) using 16 samples per treatment. For all data analyses, the professional statistical software STATISTICA 11.2 (StatSoft, 2008) was used.
Results and discusion
Wide temperature variability between the studied sites was observed, with a 1°C difference in 2017-18, as compared to the previous cycle. This is an important result that demonstrated a temperature increase in Yaqui Valley that evidences climate change. The site effect explained 95% of the temperature variation according to the randomized effect linear model used for the analysis of variance, while only 2% of the total variability was explained by crop cycle effect. There was not significant interaction between the sites and crop cycles (Fig. 2).
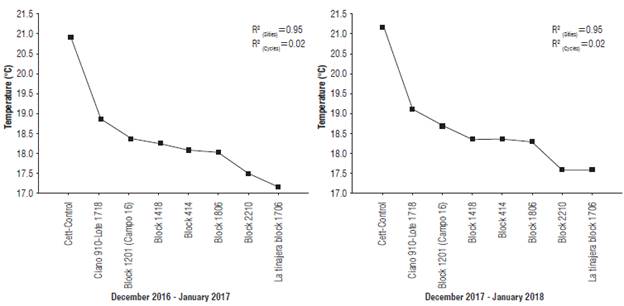
FIGURE 2 Temperature variability at eight experiment sites in Yaqui Valley during the crop cycles December 2016 - January 2017 and December 2017 - January 2018 (tillering phenophase of wheat). ñ2: coefficient of determination without adjustment for the sites and crop cycles. Squares represent mean ± standard deviation.
Some reports state that significant variation and increases (about 2°C) have been predicted for several latitudes for the next 50 years, including this area (the semiarid region of Mexico) (Garatuza-Payan et al., 2018). However, in the last crop cycle, the total variability was 1°C.
The tillering capacity experienced significant variation between the sites in both crop cycles, with a reduction in all sites as a result of the temperature variation (Fig. 3).
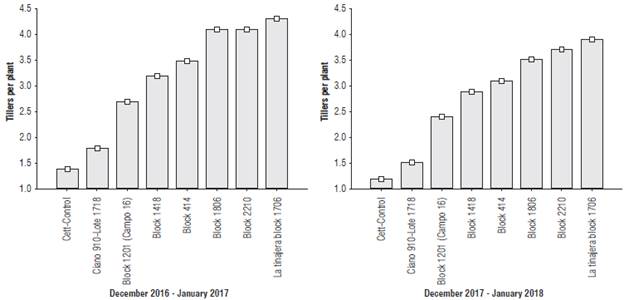
FIGURE 3 Tillering capacity in the evaluated sites in Yaqui Valley during the 2016-17 and 2017-18 crop cycles: A. December 2016 to January 2017, and B. December 2017 to January 2018. R 2 \ coefficient of determination without adjustment for the sites = 0.96, and crop cycles = 0.04. CV: coefficient of variation for sites = 8.32, and for crop cycles = 1.46. Squares represent mean ± standard deviation.
The hormonal relationship was significantly affected by the warming effect, which shows that temperature increases favor GA synthesis and inhibit ABA synthesis; this hormonal response limited the tillering capacity (Fig. 4).
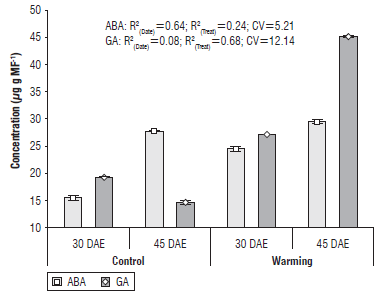
FIGURE 4 ABA and GA content at 30 and 45 d after emergence (tillering phenophases) evaluated in the higher temperature site (crop canopy +2°C) and control site in Yaqui Valley (mean of both 2016-2017 and 2017-2018 crop cycles). R2: coefficient of determination without adjustment for the phenophases and treatments; CV: coefficient of variation.
Some studies have explained that tillering cessation and number of total tillers is regulated by many biochemical, physiological, genetic and environmental factors (Xu et al., 2016). Phytotechnical factors, such as plant density (through an increase in seed density during sowing), can also affect the total tiller number (Pinto et al., 2017).
The tillering capacity in wheat determines plant canopy size (Sattar et al., 2015), photosynthetic area and, more importantly, the number of spikes and full grains per spike (Garatuza et al., 2018).
During the tillering phenophases, there were highly significant differences in the amount of cold hours between the evaluated sites and crop cycles (P = 0.015); CETT-910 (experimental warming) had the least cold hours.
Nevertheless, a reduction greater than 30% was observed in cycle 2017-18 with respect to 2016-17. The site effect contributed to 78% of the total variability in the cold hours; however, the crop cycles contributed only 18%. A significant interaction between the sites and crop cycles effects was observed (P = 0.002), which contributed 4% to the total variability (Fig. 5). These results, according to Espinosa et al. (2018), demonstrated that the tillering capacity was reduced by the reduction of cold hours in both evaluated crop cycles.
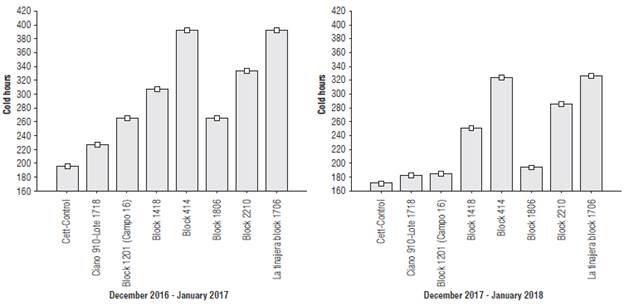
FIGURE 5 Cold hours during the December 2016 - January 2017 and December 2017 - January 2018 crop cycles. R2: coefficient of determination without adjustment for the sites = 0.78, for cycles = 0.18 and their interaction = 0.04; CV: coefficient of variation = 29.72; Squares represent mean ± standard deviation.
Historically, it has been shown that between 300 and 850 cold hours per season provide a yield near the genetic-productive potential of the varieties, with the greatest contribution to grain yield seen during the first stages, until the tillering phenophase (Zou et al., 2017). Currently, the number of cold hours or heat units is the basis of research in various regions around the world under the context of climate change (Kaur and Kaur, 2017). These studies focus mainly on gene identification and introgression for adaptability to cold hour reductions and on the achievement of hormonal stability during initial growth until the tillering phenophase (Pinto et al., 2017).
Grain yield
The temperature variation showed a negative and significative correlation with grain yield in both experiment crop cycles (Fig. 6). The observed general yield decrease was fit at 92% with the linear model used for the statistical process in the crop cycle 2016-17 and at 51% in the second crop cycle.
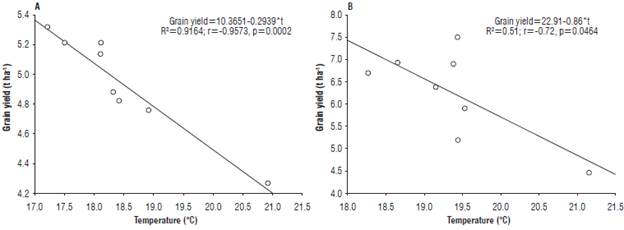
FIGURE 6 A. Effect of temperature on the grain yield during the 2016-2017, and B. 2017-2018 crop cycles. R2: coefficient of determination without adjustment; r: regression coefficient; p: probability.
Warming stresses may cause a grain yield reduction through a reduction of spike number per m2 (Shirdelmoghanloo et al., 2016) and an increase of non-viable pollen production, which makes grain formation difficult (Siebers et al., 2017). Currently, increased seed densities are used to overcome the temperature effect on tillering capacity reductions in a productive manner in agronomic practices (Lazzaro et al., 2018), which greatly increase seeding costs (Chandra et al., 2017).
Mean temperature values of 18.4°C and 18.66°C were observed during crop phenology in the 2016-17 and 2017-18 crop cycles, respectively. The regression equations were used to determine the theoretical temperature values that affected yield at 30%, between the temperature and grain yield. The first crop cycle had a theoretical temperature value of 23.7°C, whereas the second cycle had a value of 22.9°C, demonstrating a greater sensitivity of the wheat to temperature increases in the crop cycle 2017-18.
Conclusions
A temperature variability of 1°C was observed between the 2016-17 and 2017-18 crop cycles.
The tiller number was significantly reduced from four to one tiller in both evaluated crop cycles.
The warming caused a significant decrease in the ABA content and an increase in the GA, which corroborated its contribution to the wheat tillering capacity reduction in Yaqui Valley.
Although the 2017-18 crop cycle had a higher grain yield, the temperature increase had a negative and significant effect on the grain yield at all evaluated sites and crop cycles in Yaqui Valley, Mexico.


