











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Setaria sphacelata (Poaceae) is a perennial pasture grass native to tropical Africa (Hacker, 1991). It is a cross-pollinating, tetraploid species (2n=4x=36) (Hacker and Jones, 1969; Le Thierry d'Ennequin et al., 1998) of summer growth. It is cultivated in regions with rainfall exceeding 750 mm; but it also exhibits resistance to drought, flooding, and nutrient-deficient soils (Borrajo and Pizzio, 2006). In Argentina, S. sphacelata is cultivated in the northern regions, and the most agronomically important cultivars are Splenda, Kazungula, and Narok (Pensiero, 1999; Borrajo et al., 2006; Burghi et al., 2014).
Trichloris crinita (Lag.) Parodi (Poaceae) is a perennial grass species native to arid regions of the American continent (Kozub et al., 2017). It is a warm-season species with good forage quality due to its protein content and palatability (Nicora and Rúgolo de Agrasar, 1987). It is an autogamous tetraploid (2n=4x=40) (Fedorov, 1969), tolerant to drought and low-salinity (Aronson, 1989; Greco and Cavagnaro, 2003), which is used for protection against soil erosion (Dalmasso, 1994). In Argentina, T. crinita integrates the grasslands of arid and semi-arid regions (Cano, 1988). In 2012, the genus Trichloris was embedded in Leptochloa and the species T. crinita was renamed L. crinita (Snow and Peterson, 2012).
Both species are used as forage in Argentina and are strategic for the development of regional and national livestock breeding. The accessibility of molecular markers could complement the genetic improvement programs for these species. Although there are no microsatellites specifically designed for these grass species to date, a few studies have been reported using molecular markers. On the one hand, Li et al. (1998) applied random amplification of polymorphic DNA (RAPD) analysis to assess the intraspecific and interspecific variation in several species of genus Setaria, including S. sphacelata. Moreover, simple sequence repeat (SSR) markers were developed in S. italica and transferred to S. sphacelata (Gupta et al., 2012; Gupta et al., 2013; Kumari et al., 2013; Pandey et al., 2013). Cavagnaro et al. (2006) assessed the genetic diversity in T. crinita varieties using amplified fragment length polymorphism (AFLP) markers. More recently, SSR markers were developed in T. crinita using sequence data from related grass species (Kozub et al., 2018).
In order to expand the set of molecular markers available for genetic improvement programs of S. sphacelata and T. crinita, we conducted an SSR cross-amplification from related species of the Poaceae family. For this, we first optimized the DNA extraction for T. crinita.
Materials and methods
Plant materials
We studied three S. sphacelata cultivars in this research: Narok, Splenda, and Kazungula. We also included an experimentally improved population derived from Narok, called "Selección INTA" (EEA INTA Mercedes, Corrientes), in the analysis. Twenty individuals from each cultivar were analyzed in bulk.
We collected Trichloris crinita material from four sites in Argentina: one individual from La Pampa (36°35'34.4" S and 64°41'47.6" W), eight individuals from Catamarca (29°18'22" S and 65°08'40" W), seven individuals from Cordoba (three individuals from 29°57-59' S, 64°28-29' W, and 300 m a.s.l., and four individuals from 29°49-54' S, 64°27-28' W, and 310 m a.s.l.), and seven individuals from La Rioja (three individuals from 31°24-25' S, 66°46-47' W, and 490 m a.s.l. and four individuals from 31°30-32' S, 66°48-49' W, and 450 m a.s.l.) (Quiroga et al., 2010).
Plants were grown at 25°C in 20 cm pots filled with a 3:1 mixture of fertilized soil and vermiculite in a greenhouse at the Ewald A. Favret Institute of Genetics, National Institute of Agricultural Technology (IGEAF-INTA). Pots were watered approximately every 3 d. The photoperiod was 16 h light and 8 h dark.
DNA extraction
Different protocols were attempted for DNA extraction from S. sphacelata without successful results. Therefore, in order to obtain high-purity genomic DNA, we modified the extraction protocol described by Dellaporta et al. (1983). Fresh young leaves were ground in liquid nitrogen and 30-40 mg were resuspended in 800 μl of extraction buffer (50 mM Tris-HCl, 0.5 mM EDTA, 50 mM NaCl, 10% SDS, 10 mM β-mercaptoethanol, pH 8.0). Tubes were incubated at 65°C for 30 min. Then, 200 μl of potassium acetate (5 M) was added, and the tubes were incubated in ice for 20 min. The suspension was centrifuged at 12,000 rpm for 20 min at 4°C, and the supernatants were transferred into clean tubes. Next, 800 μl of chloroform:isoamyl alcohol 24:1 (v/v) was added, and the tubes were centrifuged again at 12,000 rpm for 10 min. Isopropanol (1 volume) was added to the supernatants for DNA precipitation, and the tubes were spun at 12,000 rpm for 30 min at 4°C. The supernatants were discarded and the DNA pellets resuspended in 250 μl of TE1X buffer (50 mM Tris-HCl, 10 mM EDTA, pH 8.0). Then, 2.5 μl of RNase (10 mg/ml) was added, and the tubes were incubated for 30 min at 37°C. Later, phenol (1 volume) was added, and the tubes were centrifuged at 12,000 rpm for 10 min at 4°C. The supernatants were washed with TE1X buffer and spun. One volume of chloroform:isoamyl alcohol 24:1 (v/v) was added to the supernatants, and the tubes were centrifuged again. The supernatants were transferred to new tubes, and the DNA was precipitated with isopropanol (1 volume). The tubes were incubated for 30 min at 4°C and then spun at 12,000 rpm for 40 min at 4°C. The supernatants were discarded, and the pellets were washed with 500 μl of 70% (v/v) ethanol. Finally, the tubes were centrifuged at 12,000 rpm for 2 min, and the DNA pellets were resuspended in HPLC water.
Genomic DNA from T. crinita was extracted using the Saghai-Maroof method, modified by Pérez de la Torre et al. (2008). The DNA extracted from both species was verified by 0.8% agarose gel electrophoresis.
SSR markers
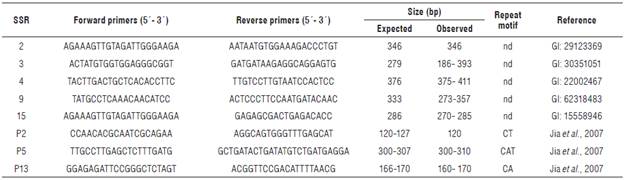
We selected a set of 29 SSR primers for cross-amplification in S. sphacelata: 20 pairs from S. italica (www.ncbi.nlm. nih.gov, Jia et al., 2007), two from Bromus tectorum (Ramakrishnan et al., 2004), one from Lolium perenne (Jones et al., 2001), three from Triticum aestivum (Roder et al., 1998), and three from Zea mays selected from http//www. agron.missouri.edu. We labeled all forward primers for S. sphacelata with FAM (6-carboxyfluorescein) and HEX (hexachloro-6-carboxyfluorescein) fluorescent dyes (Alpha DNA).
For cross-amplification in T. crinita, we selected a set of 260 SSR primers: six pairs from B. tectorum (Ramakrishnan et al., 2004), 50 from Cenchrus ciliaris (Jessup, 2005), seven from Eleusine coracana (Dida et al., 2007; Arya et al., 2009), 20 from Festuca arundinacea (Saha et al., 2003), 10 from L. perenne (Jones et al., 2001), 12 from Panicum maximum (Ebina et al., 2007; Chandra and Tiwari 2010), 20 from S. italica (Jia et al., 2007; www.ncbi.nlm.nih.gov), 66 from T. aestivum (Roder et al., 1998; http://wheat.pw.usda.gov), and 69 designed in Z. mays (http//www.agron.missouri.edu).
PCR amplification
We performed PCR reactions in a total volume of 20 which contained 75 ng of DNA, 1X PCR buffer (Invitrogen), 0.5 U of Taq DNA polymerase, 0.125 mM of each primer, 0.15 mM of dNTPs, and 2.5 mM of MgCl2. The amplification reactions were performed in a thermal cycler (Mas-tercycler Epgradient S, Eppendorf), under the following conditions: initial denaturation temperature of 94°C for 10 min, followed by 30 cycles of denaturation at 94°C for 1 min, primer annealing at 50°C for 30 s, and amplification at 72°C for 2 min, with a final extension at 72°C for 10 min. We applied each PCR four times.
In S. sphacelata, we analyzed the individuals from each cultivar in bulk. We detected SSR fragments by ABI PRISM 3130 Genetic Analyzer using Genemapper 3.4 software (Applied Biosystems). In T. crinita, we separated PCR products by 6% non-denaturing polyacrylamide gels and stained them with ethidium bromide. The fragment size for each product was determined by 100, 50, and 10 bp standard size markers (Invitrogen).
Data analysis
We estimated genetic distances among the materials using Dice and Jaccard coefficients for S. sphacelata and T. crinita, respectively. We conducted clustering between species using the Unweighted Pair-Group Method with the arithmetic Average (UPGMA). In the dendrogram, we used S. italica and C. ciliarisas external controls (out-groups) for S. sphacelata and T. crinita, respectively. We quantified the level of polymorphism for each SSR with the Polymorphism Index Content (PIC) (Botstein et al., 1980). We performed Principal Coordinates Analysis, along with the rest of the statistical analyses described in this section, using InfoGen software (Balzarini and Di Rienzo, 2016). We estimated genetic variability within and among species via an analysis of molecular variance (AMOVA) using Gen AIEx 6.2 software (Peakall and Smouse, 2006).
Results and discussion
Eight out of the 20 markers designed in S. italica were transferable to S. sphacelata. None of the primer pairs from the other species, B. tectorum (2), L. perenne (1), T. aestivum (3) and Z. mays (3), generated amplicons.
We obtained 21 different PCR products with an average of 2.62 bands per marker and with sizes ranging from 120 to 411 bp. The average transfer rate of SSR markers from S. italica to S. sphacelata was 40% (Tab. 1). Six of the eight transferred microsatellites could amplify polymorphic bands, which represents a 75% level of polymorphism. The polymorphism index content (PIC) varied from 0.27 to 0.36.
Transferability between S. italica and S. sphacelata was consistent with the results obtained by Barbará et al. (2007) for amplification within genera in monocots (40%). However, the polymorphism obtained in this study for the transferred markers to S. sphacelata (75%) was higher than the 26% observed by Jia et al. (2007) with microsatellites developed and evaluated in S. italica. It is also higher than the polymorphism obtained by Wang et al. (2005) between Poaceae species (67%) and between self-incompatible species (57%).
In order to assess the applicability of the transferred SSR markers, we performed a genetic diversity analysis in S. sphacelata cultivars (Fig. 1). The genetic distance of materials ranged from 0.18 to 0.33. The dataset computed a high cophenetic correlation between matrixes (0.892). "Selección INTA" was separated from Narok cultivar at a genetic distance of 0.18, which is expected considering that the former derives from the latter (Borrajo et al., 2009). The genetic distance between S. sphacelata and S. italica obtained in this study was similar to that reported by Kumari et al. (2013).
Previous research on S. sphacelata reports significant variability in agronomically relevant, morphological characteristics among populations, lines, and cultivars (Hacker and Cuany, 1997; Jank et al., 2007). In this context, the availability of molecular markers for the species could contribute to a production of more proficient genetic improvement programs. Furthermore, Hacker and Cuany (1997) studied variation in seed production between cultivars of S. sphacelata and found that Kazungula and Narok are the most contrasting ones. Similar results are reported by Hacker (1991), Hacker and Cuany (1997), Jank and Hacker (2004) and Jank et al. (2007), coinciding with the molecular classification obtained in this study (Fig. 1).
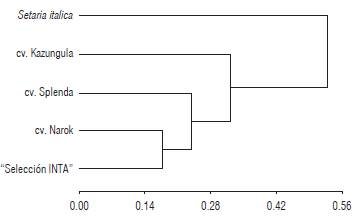
FIGURE 1 Genetic variability among Setaria sphacelata cultivars based on the 8 SSR markers transferred from S. italica. The clustering analysis was performed using the UPGMA and Dice coefficient. S. italica was used as an outgroup.
For T. crinita, 19 (out of the 260 evaluated markers) SSR were successfully transferred from five Poaceae species (a 7.3% transfer rate). Microsatellites from C. ciliaris and Z. mays showed the highest transfer rates of 18% and 10%, respectively (Tab. 2). These results are in full agreement with reports by Barbará et al. (2007) on transfer rates between genera of approximately 10% for eudicots and even lower for monocots.
TABLE 2 Summary of the SSR markers transferred to Trichloris crinita from five Poaceae species.
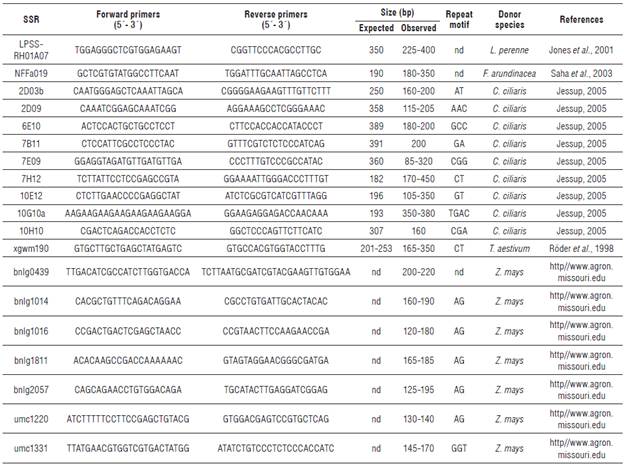
bp: base pairs; nd: not determined.
PIC varied from 0.11 to 0.36. Sixty-one different PCR products were obtained with an average of 3.21 bands per marker and sizes ranging from 85 to 450 bp. In total, 69% of the transferred SSR were polymorphic. These are high levels of polymorphism in comparison with the 34-46% obtained by Saha et al. (2006) with transferred SSR markers from F. arundinacea to Poaceae species. Our results are similar to those obtained by Wang et al. (2005) for Cynodon with SSR transferred from major cereal crops. Kozub et al. (2018) also transferred SSR markers to T. crinita. Whereas the level of polymorphism obtained in our work (69%) is higher than that obtained by the authors (37.5%), our transfer rate (7.3%) is lower than theirs (15.2%). This could be explained by the use of highly phylogenetically related species by Kozub et al. (2018).
We assessed genetic diversity among T. crinita individuals through cluster analysis. We grouped materials in four clusters, with genetic distances ranging from 0.13 to 0.37 and a cophenetic correlation coefficient of 0.905 (Fig. 2). Although the distribution of the individuals into the four clusters was not completely consistent with the collection sites, the SSR markers used for this study exhibited a potential for discriminating the individuals from Catamarca and Cordoba. Individuals from La Rioja were scattered between the groups. The sample collected in La Pampa (cluster A) was the first to separate, with a genetic distance of 0.37. Cluster B, formed by four individuals collected in Cordoba, separated at a genetic distance of 0.34. Finally, clusters C and D separated at 0.31. The first was composed of three individuals from Cordoba and two from La Rioja, and the latter consisted mainly of individuals from Catamarca.
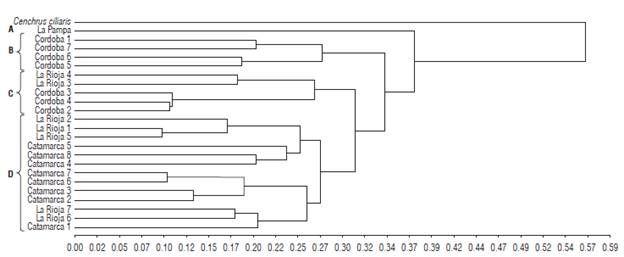
FIGURE 2 Dendrogram showing the genetic relationships among 23 Trichloris crinita individuals based on the transferred SSR markers. The clustering analysis was performed using the UPGMA and Jaccard coefficient. Letters indicate groups discriminated at 50% of the maximum estimated genetic distance. We used C. ciliaris as an outgroup.
Additionally, we performed a principal coordinates analysis (PCoA) to better visualize the variability among the individuals (Fig. 3). The distribution obtained with the PCoA is analogous to the classification obtained with the cluster analysis. Moreover, an AMOVA was computed, and we observed that 79% of the molecular variance belonged to differences within populations, whereas 21% belonged to differences among them.
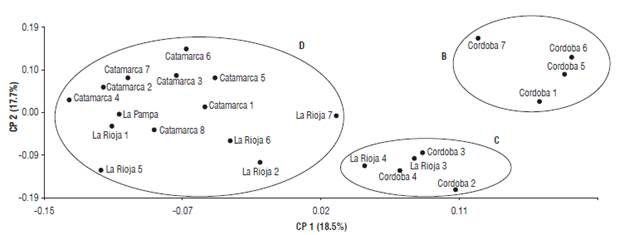
FIGURE 3 Principal coordinates analysis plot. Variability among 23 Trichloris crinita individuals based on the transferred SSR markers. Circles delimit the individuals grouped into clusters B, C, and D according to the clustering analysis showed in Figure 2.
Conclusions
The goal of this study was to generate a molecular tool for S. sphacelata and T. crinita that could be used to make more efficient genetic improvement programs in these species. The strategy was to exploit the available genetic resources of agronomically relevant crops to identify markers in grass species with limited genomic information.
We successfully transferred eight polymorphic SSR markers to S. sphacelata and 19 to T. crinita. These markers widen the available molecular resources for these forage crops, especially in T. crinita for which this study constituted the second report of transferred microsatellites in the species. As for S. sphacelata, these findings not only expanded the available genetic resources but also increased the number of methodological tools for the species with the optimization of the DNA extraction protocol. We expect the results obtained in this study will be valuable for the development of new molecular breeding programs and novel strategies to assess genetic diversity in the species.

