











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Corn is one of the most important crops in Colombia's agricultural sector. This crop represents 13% of the agricultural area in the country (CIMMYT & CIAT, 2019). More specifically, the Huila region recorded technical corn production of 23,706 t covering 5,689 ha in 2018, while traditional corn showed a production of 9,957 t with an area of 6,496 ha (Agronet, 2019).
The corn leafhopper Dalbulus maidis (DeLong) is a hemipteran insect of the family Cicadellidae known as the most important vector of corn pathogens such as Candidatus phytoplasma asteris (maize bushy stunt phytoplasma - MBSP) and Spiroplasma kunkelii (corn stunt spiroplasm - CSS) that cause the severe corn stunt complex disease symptoms that affect corn yield (Moya-Raygoza, 2002; Varón de Agudelo & Sarria Villa, 2007). The maize rayado fino virus (MRFV) belongs to this complex. However, it has less symptomatic effects compared to CSS an MBSP. The vector D. maidis can individually or simultaneously transfer the pathogens CSS, MBSP, and MRFV during the early vegetative stages of corn growth. While the pathogens multiply within the plant, at the early stages of infection, the plant may not display symptoms and has apparently normal growth. However, at the late phases of the vegetative and early reproductive stages of the corn, leaf yellowing and reddening of the leaf lamina edge is observed. Simultaneously, an alteration of the cell cycle of the plant is noted and expressed as premature flowering, the proliferation of branches, and wilting. The most severe symptoms of the disease, such as dwarfism, shortening of nodes, excessive branching, and reduction in size and quantity of the grain, are evident at the reproductive stage of corn (Varón de Agudelo & Sarria Villa, 2007). Yield losses in maize crops of 50% to 90% have been reported due to maize stunting in Colombia as well as Brazil and Argentina (Druetta et al., 2018).
Knowledge of D. maidis populations makes it possible to establish the dynamics of the corn stunt complex disease. Additionally, the morphological description of insect vector populations is the first step in understanding the biology and ecology of a species, its relationship with the environment (Varón de Agudelo & Sarria Villa, 2007) and, in this case, how these factors may subsequently relate to its role as a vector. In insects, several studies show that factors such as temperature, latitude, and elevation affect the species morphology and development (Hawkins & Lawton, 1995; Krasnov et al., 1996; Smith et al., 2000; Hodkinson, 2005; Rodríguez-Zabala et al., 2016). Larsen and Nault (1994), in Mexican populations of D. maidis, observed changes in size, weight, and pigmentation according to low temperatures, as well as morphological variations in the populations according to the season of the year. In Brazil, the influence of latitude and elevation on populations of D. maidis has been evaluated, finding morphological differences associated with variations in these two factors in the studied sites. Greater body weight, size, and pigmentation are seen at higher elevations, as well as the separation of populations into two groups (populations in the northeastern region and populations in the central-southern region) according to the sampling site (Oliveira et al., 2004).
Dalbulus maidis has a wide distribution in the Americas, from the United States to Argentina, including some Caribbean islands (CABI, 2020). Dalbulus maidis population dynamics can directly influence the disease incidence and severity (Varón de Agudelo & Sarria Villa, 2007; Oliveira et al., 2013).
Studies of D. maidis populations have not yet been carried out in Colombia. During the last few years, the Huila region has been affected by an increasing spread of corn stunting complex and increased D. maidis populations (ICA, 2019). Therefore, studies of the characterization of D. maidis are necessary, allowing for an approach to the situation in this corn-producing region, based on the morphological evaluation and abundance of individuals. Considering the hypothesis that morphological variations exist within the populations of D. maidis associated with different corn-producing areas from Huila, this research was carried out as part of the project of integrated management of phytosanitary alternatives in pests and diseases for rice-corn-cotton systems in the inter-Andean valleys and the Caribbean. The objective of this research was to characterize morphologically D. maidis populations from the corn-producing region of Huila and determine possible morphological variations between them.
Materials and methods
Insect sampling
Insect sampling was carried out according to the permit for the collection of wild species for scientific research modified by resolution 1466 of 2014 granted by ANLA to the Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA). Samples were collected in yellow and white corn crops from 40 to 70 d after planting that were conventionally established by farmers for commercial purposes. Sampling localities within the Huila region were considered based on the reported presence of D. maidis and the prevalence gradient of the disease provided by the Instituto Colombiano Agropecuario (ICA) - Huila region. The selection of localities was based on the following criteria: localities in Huila where the disease was reported for the first time related to increased leafhopper populations in 2015: Campoalegre and Palermo; localities selected as the escape zone for the disease: Tarqui and Garzón; localities with new disease reports and no reported presence of leafhoppers: Aipe and Villavieja; localities with no reports of the disease or presence of the genus Dalbulus: La Plata and Neiva. In this way, it was possible to cover areas from the south to the north of Huila (Fig. 1). After selecting the localities, samples were collected from October 2018 to May 2019. Two samples were collected from each farm visited using a Dietrick vacuum sampler or vacuum insect collector (D-VAC model 22, USA) from linear transects of 20 m from the edge of the crop towards the center since this is the pattern of arrival and dissemination of D. maidis in corn crops. The D-VAC vacuum sampler allowed the massive capture of specimens considering the behavior of D. maidis as a leafhopper and its great capacity to move from one plant to another. Additionally, the equipment allowed rapid access to the parts of the plants where D. maidis is most frequently found. A total of 72 samples corresponding to 36 corn farms were collected (Tab. 1). Subsequently, the collected material was stored in paper bags and transported in Styrofoam coolers to the Entomology laboratory of the Nataima research center of AGROSAVIA. Once in the laboratory, the samples were cleaned by removing the plant material and separating the individuals by taxonomic order, and depositing the specimens in vials with 70% alcohol at 4°C.
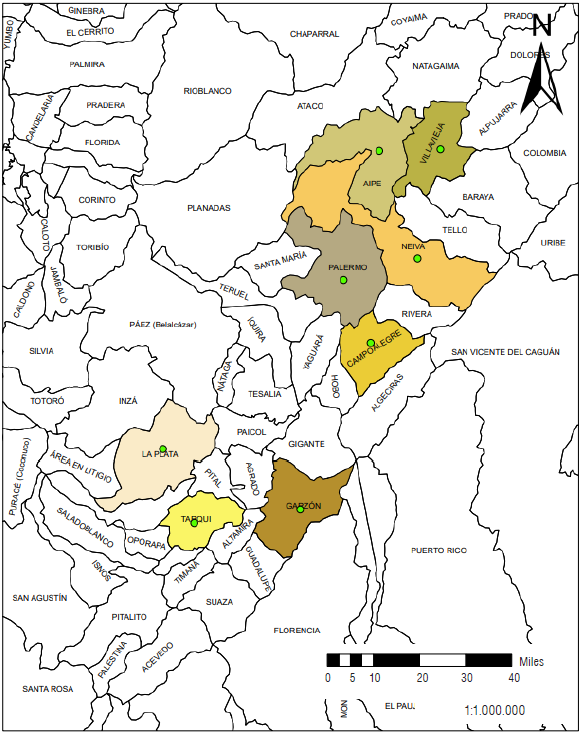
TABLE 1 Weather conditions of the localities and location of the farms where samples were collected in the Huila region.
| Sampling | Date | Location | Altitude (m a.s.l.) | Temperature (°C) | Relative humidity (%) | Precipitation (mm) | Coordinates |
|---|---|---|---|---|---|---|---|
| 1 | 10/17/2018 | Aipe | 360 | 26 - 28 | 65 - 70 | 50 - 100 | 4°26'46.9'' N 75°11'57.0" W |
| 2 | 10/17/2018 | Aipe | 359 | >28 | 65 - 70 | 50 | 3°26'42.4'' N 75°08'20.3'' W |
| 3 | 04/25/2019 | Aipe | 363 | >28 | 65 - 70 | 50 | 3°27'01.8'' N 75°08'32.2'' W |
| 4 | 11/19/2018 | Villavieja | 379 | >28 | 70 - 75 | 50 - 100 | 3°14'00.9'' N 75°13'38.9'' W |
| 5 | 11/19/2018 | Villavieja | 373 | >28 | 70 - 75 | 50 - 100 | 3°14'01.0'' N 75°13'37.4" W |
| 6 | 11/19/2018 | Villavieja | 391 | >28 | 70 - 75 | 50 - 100 | 3°14'00.8'' N 75°13'40.6'' W |
| 7 | 11/19/2018 | Villavieja | 390 | >28 | 70 - 75 | 50 - 100 | 3°14'06.7'' N 75°13'40.1'' W |
| 8 | 10/17/2018 | Campoalegre | 540 | 26 - 28 | 70 - 75 | 50 - 100 | 2°40'21.9'' N 75°21'35.0" W |
| 9 | 10/17/2018 | Campoalegre | 549 | 26 - 28 | 70 - 75 | 50 - 100 | 2°39'41.9 N 75°21'52.0'' W |
| 10 | 10/17/2018 | Campoalegre | 584 | 24 - 26 | 70 - 75 | 50 - 100 | 2°37'48.8'' N 75°21'47.7'' W |
| 11 | 11/20/2018 | Campoalegre | 535 | 26 - 28 | 70 - 75 | 50 - 100 | 2°37'46.9 N 75°21'37.5" W |
| 12 | 11/20/2018 | Garzón | 795 | 20 - 22 | 80 - 85 | 50 - 100 | 2°14'44.8'' N 75°37'06.9'' W |
| 13 | 11/20/2018 | Garzón | 859 | 20 - 22 | 80 - 85 | 50 - 100 | 2°13'20.0'' N 75°36'43.5'' W |
| 14 | 11/20/2018 | Garzón | 879 | 20 - 22 | 80 - 85 | 50 - 100 | 2°13'36-9'' N 75°36'44.5'' W |
| 15 | 11/20/2018 | Garzón | 775 | 22 - 24 | 80 - 85 | 50 - 100 | 2°09'51.3'' N 75°41'34.8'' W |
| 16 | 11/20/2018 | Garzón | 767 | 22 - 24 | 80 - 85 | 50 - 100 | 2°09'51.6'' N 75°41'34.9'' W |
| 17 | 11/20/2018 | Garzón | 723 | 22 - 24 | 80 - 85 | 50 - 100 | 2°08'51.8'' N 75°40'43.2'' W |
| 18 | 05/28/2019 | Garzón | 797 | 20 - 22 | 80 - 85 | 50 - 100 | 2°11'43.3'' N 75°37'42.2" W |
| 19 | 05/28/2019 | Garzón | 757 | 22 - 24 | 80 - 85 | 50 - 100 | 2°11'06.1" N 75°39'17.1" W |
| 20 | 05/28/2019 | Garzón | 804 | 20 - 22 | 80 - 85 | 50 - 100 | 2°14'45.6'' N 75°37'39.0'' W |
| 21 | 05/28/2019 | Garzón | 808 | 20 - 22 | 80 - 85 | 50 - 100 | 2°14'49.6'' N 75°37'42.9' 'W |
| 22 | 05/28/2019 | Garzón | 817 | 20 - 22 | 80 - 85 | 50 - 100 | 2°12'41.0'' N 75°37'35.7'' W |
| 23 | 10/18/2018 | La Plata | 1123 | 18 - 20 | 80 - 85 | 150 - 200 | 2°22'06.6'' N 75°53'44.0'' W |
| 24 | 10/18/2018 | La Plata | 1060 | 18 - 20 | 80 - 85 | 150 - 200 | 2°22'27.8'' N 75°53'38.3'' W |
| 25 | 04/24/2019 | Neiva | 417 | 26 - 28 | 70 - 75 | 50 - 100 | 3°00'35.1'' N 75°15'57.8'' W |
| 26 | 11/19/2018 | Palermo | 508 | 26 - 28 | 75 - 80 | 50 - 100 | 2°51'32.2'' N 75°21'18.2'' W |
| 27 | 11/19/2018 | Palermo | 496 | 26 - 28 | 75 - 80 | 50 - 100 | 2°50'44.6'' N 75°20'46.3'' W |
| 28 | 05/27/2019 | Tarqui | 810 | 22 - 24 | 75 - 80 | 150 - 200 | 2°04'25.2'' N 75°21'57.3'' W |
| 29 | 05/27/2019 | Tarqui | 799 | 22 - 24 | 75 - 80 | 150 - 200 | 2°06'41.1'' N 75°47'19.0'' W |
| 30 | 05/27/2019 | Tarqui | 801 | 22 - 24 | 75 - 80 | 150 - 200 | 2°06'45.3'' N 75°47'11.7'' W |
| 31 | 05/27/2019 | Tarqui | 798 | 22 - 24 | 75 - 80 | 150 - 200 | 2°06'48.0'' N 75°47'10.8'' W |
| 32 | 05/27/2019 | Tarqui | 797 | 22 - 24 | 75 - 80 | 150 - 200 | 2°06'47.9'' N 75°47'10.1'' W |
| 33 | 05/27/2019 | Tarqui | 775 | 22 - 24 | 75 - 80 | 150 - 200 | 2°07'00.8'' N 75°46'50.8'' W |
Taxonomic confirmation of the Dalbulus maidis species
Species level confirmation was done through higher taxon order Hemiptera, family Cicadellidae to species. The morphological characters used in this study were selected following four taxonomic keys of the group that included the characters of the antennal flagellum, distal forewing cells, first posterior tarsomere, ocellar spot, and body coloration (Triplehorn & Nault, 1985; Knight & Webb, 1993; Dietrich, 2005; Triplehorn & Johnson, 2005). Female and male insects with complete morphological structures were used for determination. Morphological characters were observed with a Leica EZ4® stereoscope (Heerbrugg, Switzerland). The specimens to be characterized were entomologically mounted and deposited in the National Taxonomic Collection of Insects "Luis María Murillo" (CTNI) under custody of AGROSAVIA. The species were corroborated by digital photographs sent to an expert, Dr. Gustavo Moya-Raygoza (Department of Botany and Zoology, the University of Guadalajara, Mexico) (G. Moya-Raygoza, personal communication, November 9, 2020).
Dalbulus maidis abundance
The population frequency of D. maidis was evaluated by counting individuals collected from each of the corn-producing areas in the Huila region. The frequency of D. maidis was measured by the total number of individuals collected at each locality. To identify a possible relationship between morphology, abundance, and presence of D. maidis, the database of the agroclimatic network of Huila, generated by the Institute of Hydrology, Meteorology and Environmental Studies (IDEAM) was consulted. The average temperature, precipitation, and relative air humidity (RH) data generated by the network for each locality were included in the analyses.
Intraspecific morphological variations of Dalbulus maidis
After confirming that the insect populations collected in the eight maize-producing areas of Huila belonged to the species D. maidis, morphological differences within the population were determined, using a subsample of 10% of the individuals from areas with an abundance greater than 1,000 individuals. All individuals were analyzed in those areas where the abundance was less than 1,000 individuals. The separation of individuals into morphotypes was carried out in a dichotomous manner (presence/absence) by observing the following qualitative characters: head color, abdomen pigmentation, number of ocellar spots, and presence of anterior spots on the head (Triplehorn & Nault, 1985; Knight & Webb, 1993; Oliveira et al., 2004).
Seven morphometric characters were selected to determine differences within the populations across localities, morphotypes, and sex of individuals. The characters were selected according to their taxonomic significance to the species and for their usefulness for separating population morphotypes (Larsen & Nault, 1994; Oliveira et al., 2004). The characters tested were antenna length, ocellar spot diameter, head length, distance between compound eyes, right-wing length, left-wing length, and total body length. Finally, genitalia of males and females were extracted, and the aedeagus length, distance between the divergent apical processes, and ovipositor length were measured to determine any differences in the chosen specimens (Oliveira et al., 2004).
A photographic record of morphological characters was taken using a Leica EZ4W® stereoscope with a built-in 5.0-megapixel CMOS WiFi camera with multiple output options, capturing images of the dorsal and ventral views (20x and 35x) of the individuals. Using the Leica AirLab® application, morphometric character measurements were obtained.
Data analysis
The suitability of each morphological character for morphotype separation was evaluated employing a monothe-matic clustering analysis of binary variables using the MONA function (Struyf et al., 1997). For each morphometric character measured, an analysis of variance was performed to compare significant differences between individuals by locality (with different environmental conditions), morphotypes determined, and sex (LSD-Fisher). A factor analysis for mixed data (FAMD) was performed that combines the principal component technique with multiple correspondence analysis of qualitative and quantitative variables to observe their interactions (data not shown). A locality and morphotype cluster analysis was performed using Ward's method (Ward Jr., 1963) and the Bray-Curtis distance (Bray & Curtis, 1957). Analyses were performed using the statistical software R version 4.0.2 (R Core Team, 2020) with the packages FactoMineR, Factoextra, and Vegan (Lê et al., 2008; Kassambara & Mündt, 2017; Oksanen et al, 2019).
Results and discussion
Dalbulus maidis abundance
A total of 6,722 specimens of Dalbulus maidis were found in the study area. The areas with the highest abundance were Garzón and Tarqui with 3,820 and 2,650 individuals, respectively. In the Aipe-Villavieja, La Plata and Palermo areas, D. maidis abundance levels were null (Fig. 2). The availability of corn is an important factor affecting the abundance of D. maidis since they complete their biological cycle on this plant species (Meneses et al., 2016). The localities whose abundance was zero had the areas planted with corn decreased by the time of sampling. The migration processes of the species must be considered since D. maidis has good migration capacity that allows it to move long distances to a maximum of 10 km (Taylor et al., 1993). In our study, we must consider the displacement of corn crops in the producing areas of Huila since the localities of Tarqui and Garzón were selected by farmers for planting corn as escape zones from the problems caused by the stunting of corn in Campoalegre and Palermo. However, with the greater availability of food in these areas, there may be greater migration and an increase in the populations in Tarqui and Garzón with a greater abundance of leafhoppers in these areas.
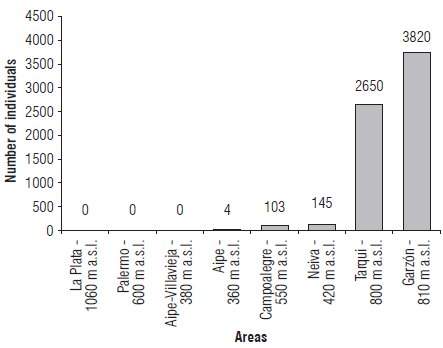
In combination with corn availability, climatic conditions can become important influences on the abundance of D. maidis in crops (Oliveira et al., 2013; Meneses et al., 2016). The eight localities were arranged into three groups according to climatic and altitude conditions (Tab. 2). The highest abundance was observed in Garzón and Tarqui, areas that had warm, semi-humid weather with an altitude range from 720 to 880 m a.s.l. and temperature from 22°C to 24°C. In contrast, the areas of Neiva and Campoalegre had warm-semiarid weather and were located from 360 to 585 m a.s.l. showing a much lower abundance compared to the Garzón and Tarqui areas (Tab. 2). The localities of Tarqui and Garzón showed temperatures optimal for D. maidis survival and fecundity (Meneses et al., 2016). Therefore, the strategy of establishing new production zones in locations with favorable conditions for the development and multiplication of D. maidis may increase the risk of introducing high populations of this insect to new areas and, consequently, the risk of dissemination of the maize stunt complex.
TABLE 2 Abundance of Dalbulus maidis and weather conditions of eight corn-producing areas from Huila.

Nelva 1:1 Campoalegre 1:1
The time of sampling should be considered since the environmental conditions that may affect populations vary over time. In Brazil, variations in the abundance of Cicadellidae species were observed in 2005, 2006 and 2007 in 48 localities (Oliveira et al., 2013). Dalbulus maidis has shown changes in the abundance of populations considering the growing seasons with an increase of individuals in the months of the dry season compared to the abundance in the rainy season (Ávila & Arce, 2008; Meneses et al., 2016).
Morphotype evaluation
The data matrix cluster allowed identifying the separation of five groups by the morphological characteristics observed. Among the individuals evaluated, four morphotypes were described, and a group corresponding to another Cicadellidae genus, considered an external group, was discarded from the morphotype analysis of this study: morphotype II: absence of the dark abdomen character (Ab_dark), present in the other four morphotypes; morphotype III: missing opaque yellow head character (He_opye); external group: number of ocellar spots (N_ocs). Morphotypes I and IV were differentiated by the character anterior spots (Sp_ant), present in morphotype IV and absent in morphotype I (Fig. 3).
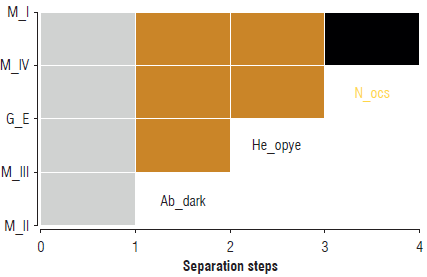
FIGURE 3 Separation banner cluster of the four Dalbulus maidis population morphotypes from Huila M - morphotype; G_E - external group; Ab_dark - dark abdomen character; He_opye - opaque yellow head character; N_ocs - number of ocellar spots. The black color indicates absence of the character, and the orange color indicates presence of the character.
The main characteristics of morphotype I were the opaque yellow head color and the dark brown abdomen pigmentation (Fig. 4 A, D, G). Morphotype II was characterized by the opaque yellow head and mainly by the white or pale-yellow abdomen (Fig. 4 B, E, H). Morphotype III was characterized by the abdomen and head with dark brown coloration (Fig. 4 C, F, I). Morphotype IV had similar characteristics to morphotype I; however, it was distinguished by the presence of a pair of spots on the back of the head, very close to the ocellar spots (Fig. 4 J-M).
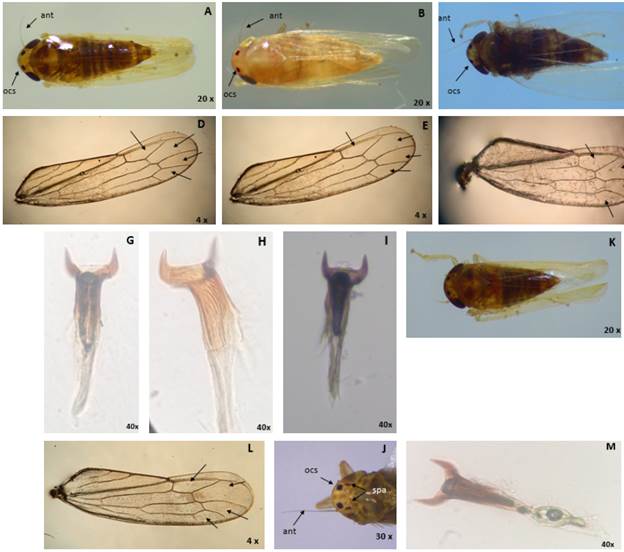
FIGURE 4 Morphological characters color of the head, ocellar spots (ocs), antenna (ant), and pigmentation of the abdomen: A) morphotype I; B) morphotype II; C) morphotype III. Anterior wing apical cells: D) morphotype I; E) morphotype II; F) morphotype III. Aedeagus with divergent apical processes: G) morphotype I; H) morphotype II; I) morphotype III. J) ocellar spots (ocs), antenna (ant), anterior spots (spa) in morphotype IV. K) Pigmentation of the abdomen and head in morphotype IV. L) apical cells anterior wings in morphotype IV. M) Aedeagus with divergent apical processes in morphotype IV.
Morphotypes I, II and IV were found in Neiva, Campoalegre, Garzón and Tarqui. Morphotype III was located only in Tarqui. The highest abundance of morphotypes was observed in Tarqui.
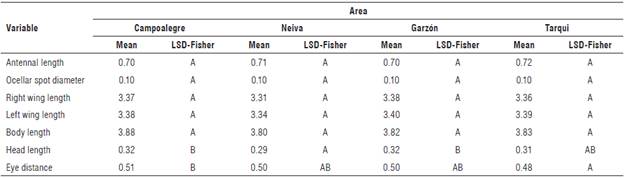
Morphological comparison of individuals by locality
Normality and statistical homogeneity were corroborated for the seven morphometric characters. Within the populations of D. maidis, antennal length, ocellar spot diameter, right and left-wing length and total body length did not show significant differences among the four corn-producing areas (Tab. 3). However, the characters of head length and distance between the eyes did show significant differences among corn-producing areas. Individuals from Garzón and Campoalegre populations differed significantly from those from Neiva; additionally, individuals from Tarqui showed significant differences with Campoalegre with respect to the character distance between eyes. Significant differences were found in the head length and distance between the eyes in the populations of D. maidis evaluated in the four localities of Huila; however, the average values for the characters are close, showing little variation in the characters. Thus, Neiva and Tarqui showed an average of 0.29 and 0.31 mm for head length, respectively, while Garzón and Campoalegre showed an average of 0.32 mm (Tab. 3). The characters antennal length, right-wing length, left-wing length, total length, and ocellar spot diameter did not show significant differences between them. Considering that the environmental factors reported as contributing to morphological variations (temperature, elevation, and latitude) did not show large variations in the populations of Neiva, Garzón, Campoalegre, and Tarqui (Tab. 2), these environmental factors should be considered for the evaluation of the differences between Colombian populations of D. maidis from different localities. Additionally, the distances between localities are relatively short, so migration events of individuals between populations are possible.
Morphological comparison of individuals by morphotypes
The morphometric character comparison among the morphotypes showed that five of them did not show significant differences (Tab. 4). As in the comparisons by area, the character distance between eyes showed a significant difference between morphotypes, separating morphotype III from the others. Total body length showed significant differences between morphotypes I and II. The antennal length was measured because the genus Dalbulus has the third antennal segment longer than other Cicadellidae family members (Knight & Webb, 1993; Triplehorn & Johnson, 2005). However, no records about antennal size were found to be specific for D. maidis, so the length range of a diagnostic character at the genus level may be somewhat broad and not show a difference at the morphotype level. The ocellar spot diameter is a relevant character in D. maidis since it is defined in this specimen as twice the size of the ocelli (Nielson, 1968; Knight & Webb, 1993; Pinedo-Escatel & Blanco-Rodríguez, 2016). The ocelli of D. maidis are very small, so the ocellar spots size is in proportion to the character of the species, but it does not allow seeing differences at the level of morphotypes in the species. The distance between the compound eyes and the length of the head are characters that have been previously evaluated for the observation of morphological variations (polymorphisms) within the species D. maidis (Oliveira et al., 2004). However, in our study, these characters did not show significant differences among the four morphotypes found. Variations in these two characters could be influenced by altitude and temperature; among the populations evaluated, these environmental conditions do not show large differences, despite their fluctuation in each locality. The length of the anterior wings has also been used to determine morphological variations among D. maidis populations, as in the case of the Brazilian populations, which allow separating the evaluated localities into two groups (Oliveira et al., 2004). However, the analysis of the morphotypes of the populations showed no significant differences for this character, although the left wing is slightly longer than the right wing (Tab. 4).
TABLE 4 Statistical analysis with the Fisher-LSD test of ten morphometric characters of Dalbulus maidis populations morphotypes from Huila.
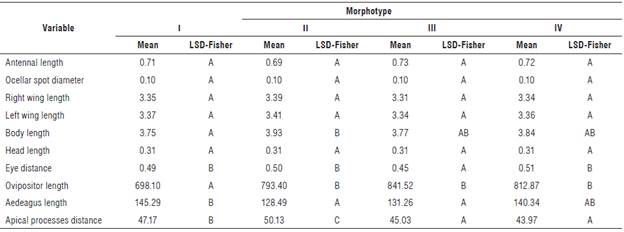
Significant differences were also found between morphotypes for the characteristic of the distance between the eyes, where morphotype III showed the lowest value for this characteristic (0.45 mm) separating it from the other morphotypes (Tab. 4). Thus, morphotype III separates a little more from the other individuals with another character of the head, since this morphotype is the only one showing a darker coloration of the head different from the opaque yellow characteristic of D. maidis. Additionally, considering that the morphotype III was only found in Tarqui, it is the most differentiated morphotype of those evaluated in the populations of Huila.
Statistical analysis of the ovipositor length from females of the population showed significant differences for morphotype I compared to the other three morphotypes (P-value 0.0016). We observed that morphotype I had a shorter ovipositor (698.10 µm) on average, compared to morphotypes II, III, and IV that had an average length of 800 µm (Tab. 4). A comparison of the aedeagus length showed significant differences between morphotype I and morphotypes II and III (Tab. 4). Morphotype I had the longest average length at 145.29 µm, while morphotype II had the shortest length among the four morphotypes with around 128.49 µm. Morphotype III showed no significant differences from morphotypes II and IV, while morphotype IV was between the values of morphotypes II and III and I (140.34 µm). The observations of the ovipositor and aedeagus provide information for future research to clarify the reproductive processes and clarify if the differences in size of the genitalia could generate the reproductive isolation between morphotypes.
The distance between the divergent apical processes of the aedeagus was also evaluated, and significant differences were found across the four morphotypes (Tab. 4). Morphotypes IV and III did not differ among themselves; however, they differed from morphotypes I and II. Morphotype II showed the greatest distance between divergent processes, 50.13 µm on average, while morphotypes IV and III obtained the lowest values, 43.97 and 45.03 µm, respectively. Regarding the comparison between males and females, significant differences were only found in the total body length, with females being longer than males (Tab. 5). The total sized body reported for the Dalbulus species is between 3.5 and 4.0 mm, a range in which the four morphotypes were found (Nielson, 1968; Larsen & Nault, 1994; Oliveira et al, 2004).
TABLE 5 Statistical analysis with the Fisher-LSD test of seven morphometric characters of males and females of Dalbulus maidis populations from four Huila areas.
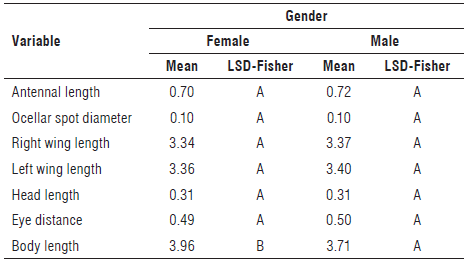
Differences in the variable of total body length could be seen between males and females of D. maidis. In total body length, the females tended to be slightly larger than males as previously reported in several studies for this species (Nielson, 1968; Triplehorn & Nault, 1985; Larsen & Nault, 1994; Oliveira et al, 2004), with a higher frequency of females ranging in size from 3.70 to 4.70 mm, while the male range was mainly between 3.50 and 3.70 mm. Although most males were in the 3.50 to 3.70 mm size range, the overall spread of the data showed a range from 3.40 to 4.00 mm. These values are within the range defined by Nielson (1968), who described that males of D. maidis have a length from 3.50 to 4.00 mm, while females have a length from 4.00 to 4.10 mm. This also matches the results of our study, since the individuals had values close to 4.50 mm (Tab. 5). Female insects are generally larger than males (Triplehorn & Johnson, 2005). In D. maidis, the difference in size between males and females is related to the development rate of the individuals, since females have a slower development with more time for feeding and growth than males (Oliveira et al, 2004). Additionally, an adaptive explanation is given for this characteristic since females are responsible for breeding and laying eggs and being larger increases their probability of reproduction and breeding compared to smaller females (Larsen & Nault, 1994). The diameter of the ocellar spot, antennal length, length of the head, distance between the eyes, and length of the right and left wing are not differential characters between males and females.
A separate cluster of individuals was obtained from the average of the seven morphometric characters of each morphotype by corn-producing area, allowing the identification of two main clades (Fig. 5). Thus, in the dendrogram, morphotype I from Garzón and morphotype IV from Neiva was the group most separated from the other individuals; the second clade showed three subgroups. Morphotypes II and IV from Campoalegre and morphotype II from Tarqui showed greater similarity among the characters, so they were grouped in the same clade. Morphotypes IV from Garzón, I from Tarqui and Neiva and IV from Tarqui formed the next group. Morphotypes III from Tarqui, I from Campoalegre, and II from Neiva were grouped in the third subgroup on the dendrogram.
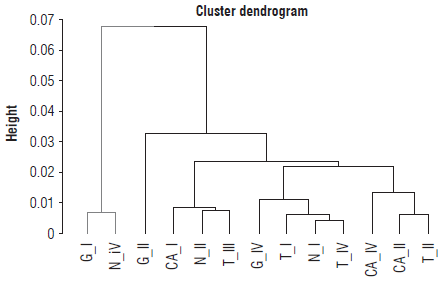
FIGURE 5 Clustering by area and morphotype of Dalbulus maidis populations from corn-producing areas in Huila. G_l - Garzón morphotype I; G_ll - Garzón morphotype II; GJV- Garzón morphotype IV; N_l - Neiva morphotype I; N_ll - Neiva morphotype II; NJV - Neiva morphotype IV; CA I - Campoalegre morphotype I; СА М - Campoalegre morphotype II; CAJV - Campoalegre morphotype IV; T_l - Tarqui morphotype I; T_ll - Tarqui morphotype II; TJII - Tarqui morphotype III; T IV - Tarqui morphotype IV
Conclusions
The morphological and morphometric analyses carried out in this study made it possible to separate the individuals of the D. maidis populations of the department of Huila into four morphotypes. Although morphological variations did not show significant interpopulation differences, variations at the intraspecific level were observed in the four localities. As such, this study could serve as a baseline for further studies of Colombian populations of Dalbulus maidis with a morphological description of the species in the department of Huila.

