











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
En la última década del siglo XX se descubrió un novedoso mecanismo epigenético de control postranscipcional en las plantas y en Caenorhabditis elegans, que se denominó RNA de interferencia, o RNAi. Los protagonistas del mecanismo son moléculas pequeñas de RNA que, en asociación con las proteínas de la familia Argonauta, crean complejos capaces de reconocer moléculas de RNA o DNA e interactuar con ellas 1. Uno de estos mecanismos es la vía de silenciamiento de genes mediado por microRNA o mi-RNA 1.
Estos mi-RNA tienen un alto potencial como biomarcadores para diversas enfermedades porque se pueden detectar en la circulación sanguínea, la saliva y el líquido cefalorraquídeo con una estabilidad sorprendente 2,3. Incluso se ha visto que permanecen estables durante el almacenamiento prolongado, la ebullición, los ciclos de fusión-congelamiento y los cambios de pH. Esto se debe al transporte acoplado a vesículas, proteínas como las de la familia Argonauta o complejos lipoproteicos como la lipoproteína de alta densidad (HDL) 3.
Los mi-RNA son ácidos nucleicos de ≈ 22 nucleótidos que regulan la traducción de mRNA codificantes. Los genes que codifican para estos RNA están localizados, comúnmente, en grupos policistrónicos dentro de los cromosomas. Algunos de estos mi-RNA son codificados por sus propios genes (≈ 42 % intergénicos), mientras que otros están incluidos dentro de genes codificantes para proteínas ya sean intrónicos (≈ 44 %), exónicos (≈ 7 %) o de las regiones no traducidas 3'(3'UTR) (≈ 1,5 %) y 5'UTR (≈ 1 %) 4.
La producción de mi-RNA maduros es un proceso dividido en dos fases (transcripción de los genes y procesamiento postranscripcional) 4,5. La primera fase es la producción de mi-RNA primarios la cual, dependiendo de si el mi-RNA es intergénico o intragénico, consta de uno o dos pasos. Si se trata de un mi-RNA intergénico, la RNA polimerasa II o III lo transcribe a un mi-RNA primario (pri-mi-RNA) que puede ser de algunas kilobases de largo. Los mi-RNA cuyos genes se encuentran dentro de un gen codificante para proteína tienen que pasar por un proceso de maduración del mRNA por corte de intrones y empalme de exones, para producir su pri-mi-RNA. La segunda fase consta de cuatro pasos; el primero es la escisión de una parte del pri-miRNA por parte de la RN-asa endonucleasa III (Drosha) y la proteína con la cual se acopla, DGCR8 (Pasha) para producir los miRNA precursores (pre-mi-RNA); el segundo paso es la translocación del pre-mi-RNA por las proteínas RanGTP y Exportina 5 (Exp5); una vez en el citoplasma, la ribonucleasa citoplasmática RNasa III (Dicer) junto con las proteínas TRBP y PACT cortan los bucles de los pre-mi-RNA dejando como producto un par de cadenas de ≈ 22 nucleótidos, uno de ellos el mi-RNA maduro, y el otro, una secuencia llamada pasajera que se degrada posteriormente; el cuarto y último paso de esta fase es la asociación con el complejo silenciador inducido por RNA (RISC), que es formado en los mamíferos por las proteínas Dicer, TRBP, PACT, Gremin 3 y una de la familia Argonauta (AGO) que lleva a cabo el silenciamiento de los mRNA (Figura 1) 4.
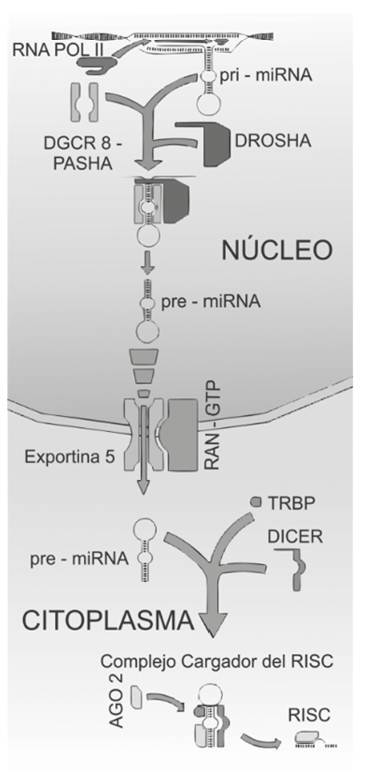
Figura 1 Biogénesis de los miRNA. RNA POL II: RNA polimerasa II. Primi RNA: micro-RNA primario. DGCR8-PASHA: proteína de la región crítica del síndrome de Di George 8. Pre-mi-RNA: micro-RNA precursor. Exportina 5 y RAN-GTP: transportadores de la membrana nuclear. AGO 2: proteína argonauta de unión a TRBP y Dicer con actividad RNasa. RISC: complejo silenciador inducido por RNA
En los mamíferos se han hallado cuatro proteínas AGO diferentes vinculadas a la conservación extracelular de los mi-RNA y, como se explicó anteriormente, en la formación del RISC. En solo una de las cuatro (AGO2) se ha evidenciado una actividad de ribonucleasa que es importante para el silenciamiento de los mRNA posterior al reconocimiento por el mi-RNA del RISC correspondiente. Cabe aclarar que el reconocimiento mi-RNA-mRNA se da por complementariedad de una secuencia semilla que forma parte del mi-RNA, pero no por toda la secuencia de nucleótidos 4. Así, la función de los mi-RNA consiste en la regulación postranscripcional de la expresión génica, mediante el bloqueo por complementariedad parcial antisentido en las regiones no traducibles (UTR) del mRNA blanco. Cada mi-RNA puede inhibir varios mRNA y cada mRNA puede ser el blanco de diferentes miRNA. La mayoría de los mi-RNA son altamente conservados y muestran una distribución específica de tejido, papel importante en la función celular y fisiopatológica de los diferentes órganos del cuerpo humano 6.
Los mi-RNA juegan un papel regulador importante en muchos procesos celulares, incluyendo la diferenciación, la transformación neoplásica, la replicación y la regeneración celular. Se han encontrado cambios en la expresión de diferentes mi-RNA en diversas enfermedades, lo que ha llevado a estudiarlos como biomarcadores potenciales. Como se mencionó anteriormente, los mi-RNA se pueden encontrar en diferentes fluidos en los que presentan varias características de buenos biomarcadores: son estables, las secuencias de la mayoría están conservadas entre diferentes especies, la expresión de algunos mi-RNA es específica de tejidos o escenarios biológicos y sus niveles se pueden estudiar fácilmente por varios métodos como los basados en microarreglos 7-9. Ya se han asociado los cambios en el nivel de varios mi- RNA en plasma, suero, orina y saliva con diferentes enfermedades incluyendo cánceres, daño hepático y cardiopatías. Además, el uso de los niveles de mi- RNA específicos para órganos en fluidos corporales puede ser una posibilidad importante para monitorear condiciones fisiopatológicas de órganos específicos.
Los mi-RNA circulantes tienen otras ventajas como biomarcadores informativos en comparación con los biomarcadores sanguíneos basados en proteínas. Mientras que una cantidad baja de proteína en sangre puede obstaculizar la detección de algunos biomarcadores basados en proteínas, la mayoría de los mi-RNA circulantes se pueden detectar con facilidad por métodos como la PCR. Además, los biomarcadores proteicos sufren diferentes modificaciones postraduccionales que pueden afectar la exactitud de la medición, mientras que las especies de mi-RNA son relativamente homogéneas. El hecho de que se hayan encontrado y establecido algunos mi-RNA con un perfil de expresión muy específico para algunos órganos abre la posibilidad importante de utilizar sus niveles en sangre para monitorear de forma precisa el estado de órganos específicos. Encontrar biomarcadores informativos no es solo clave para entender la fisiopatología de procesos patológicos, sino que también resulta crítico para el desarrollo de nuevas terapias 7.
Se ha informado el potencial de los mi-RNA como biomarcadores en diferentes enfermedades. La mayoría de los esfuerzos se han enfocado en el descubrimiento de biomarcadores en neoplasias, pero cabe destacar otros tipos de enfermedades no neoplásicas como cardiovasculares, hepáticas, pulmonares y autoinmunes. Hasta el momento las investigaciones con mayor evidencia se han hecho en cáncer y en menor medida en enfermedades hepáticas y cardiovasculares. A continuación se profundiza un poco más en estas tres áreas.
Micro-RNA como biomarcadores potenciales en cáncer
Debido a la regulación fundamental que tienen a su cargo, los mi-RNA no podían estar ausentes en el proceso de desarrollo y formación del cáncer; de ahí que jueguen un papel crucial en el establecimiento del diagnóstico, el pronóstico e incluso la respuesta a los tratamientos antineoplásicos 8-10.
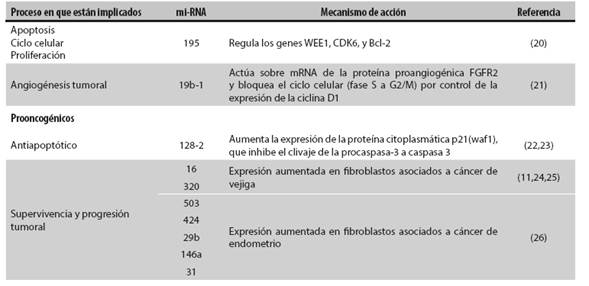
El gran potencial de los mi-RNA de influir sobre el proceso de desarrollo tumoral se debe a la acción reguladora que ejercen muchos de ellos sobre procesos vitales para el desarrollo de las neoplasias como la proliferación celular, la apoptosis y la angiogénesis, e incluso sobre la organogénesis y la hematopoyesis. Se ha implicado a los mi-RNA en el desarrollo de muchas neoplasias como leucemia, neuroblastoma, adenoma hipofisiario, cáncer de mama, de tiroides, colorrectal, de pulmón y hepatocarcinoma 11.
Se ha encontrado una serie de mi-RNA implicados en cada una de las etapas de desarrollo del cáncer. Esto se debe posiblemente a dos elementos, a saber: primero, que los mi-RNA están regulados por oncogenes o genes supresores de tumor 12,13 por lo que son elementos importantes en el desarrollo tumoral. En segundo lugar, se ha evidenciado que un gran número de mi-RNA identificados en tejidos tumorales tienen dianas similares a los oncogenes o a los genes supresores de tumor, modulando las vías de expresión de estos genes, con lo que adquieren gran importancia en el entendimiento del desarrollo tumoral 11. Por ejemplo, se ha visto que algunos mi-RNA juegan un papel importante en la regulación de los primeros pasos de las metástasis, específicamente en el proceso de invasión del mesénquima por las células epiteliales cancerosas, lo que le confiere la malignidad al tumor (Tabla 1) 14.
Su potencial como biomarcadores nace de la identificación de diversos mi-RNA circulantes tanto en sangre completa como en suero en concentraciones similares (15); su extracción permite identificar perfiles específicos de expresión de mi-RNA para los diferentes tipos de cáncer cuyos niveles difieren de los obtenidos en individuos sanos 11,16, lo que permite la distinción y convierte a los mi-RNA en una poderosa herramienta diagnóstica.
Igualmente, la identificación de perfiles específicos de expresión de mi-RNA ha permitido clasificar, con base en los niveles de expresión de algunos de estos biomarcadores, el tejido de origen de algunos tumores mal diferenciados 17. En la identificación del origen de metástasis los mi-RNA han superado a los mRNA; se considera que los primeros son biomarcadores más confiables y distintivos para este propósito 11.
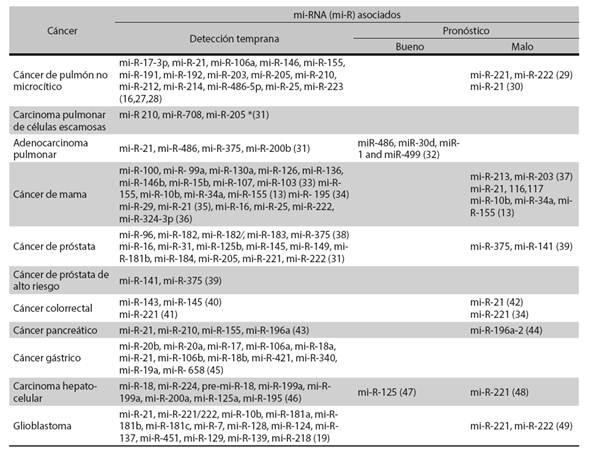
Aún no se ha descrito la importancia clínica de los perfiles de expresión de los mi-RNA para el cáncer, pero prometen ser excelentes biomarcadores específicos para el diagnóstico y pronóstico de esta enfermedad (18). Se ha propuesto a los mi-RNA como biomarcadores para la detección temprana y el pronóstico de cáncer de ovario, colorrectal, pancreático, mamario, pulmonar, prostático, gástrico, hepatocelular y glioblastoma (Tabla 2) 11,19.
Micro-RNA como biomarcadores potenciales en enfermedades cardiovasculares
La regulación mediada por mi-RNA ha sido vinculada a procesos patológicos de alteración endotelial relacionados con la edad que alteran la función protectora vascular de este tejido. En particular se ha documentado el aumento de miR-29 en células endoteliales senescentes en cultivo y se ha vinculado con una disminución en la producción de elastina y de colágeno que lleva a susceptibilidad a la formación de aneurismas. Esto a tal punto que se logró demostrar en modelos murinos un efecto protector de la inhibición de miR-29 para la formación de aneurismas 3.
Tabla 2 Micro-RNA identificados como biomarcadores potenciales para la detección temprana y el pronóstico de diferentes tipos de cáncer
Roncarati y colaboradores 50, por ejemplo, hicieron un estudio con el objetivo de caracterizar la expresión en plasma de 21 miRNA candidatos a biomarcadores en pacientes con cardiomiopatía hipertrófica (CH). Incluyeron 41 casos y 41 controles, apareados por edad y sexo, a quienes se les determinó el perfil de expresión de dichos mi-RNA y se relacionaron los niveles encontrados con cuatro parámetros imaginológicos de hipertrofia ventricular izquierda (HVI) y un puntaje de 0 a 4 para fibrosis usando captación tardía de gadolinio en la resonancia magnética cardíaca (RMC). Los parámetros para HVI fueron grosor máximo del ventrículo izquierdo (VI) e índice de hipertofia en el ecocardiograma transtorácico, y grosor máximo del VI e índice de masa del mismo (IMVI) en la RMC. Finalmente, compararon estos perfiles con los de HVI por aumento de presiones, específicamente en 12 pacientes con estenosis aórtica grave. Se encontró sensibilidad y especificidad de 5 mi-RNA (miR-29a, -27a, -199a-5p, -21, -155) para CH, y relación con al menos uno de los parámetros imaginológicos; de ellos, miR- 29a demostró relación con tres de cuatro parámetros y tendencia con el cuarto (IMVI en RMC). Además, este miR aumenta en la HVI por CH en contraste con otros mi-RNA hallados tanto en HVI por CH como por estenosis aórtica 50.
Otro mi-RNA cuyos niveles se elevan con la edad y que ha mostrado protección cardiovascular es miR- 146 por su acción antiinflamatoria, que logra al inhibir la producción de la proteína quinasa asociada al receptor de interleucinas (IRAK) (3), por lo que su disminución se puede interpretar como biomarcador de un estado proinflamatorio, factor de riesgo para la formación de trombos debida a la ruptura de una placa aterosclerótica.
De manera específica y comprobada para el caso de enfermedad arterial coronaria (EAC) se ha visto que un aumento de la expresión y la liberación al torrente sanguíneo de miR-133 y -208a y la disminución de miR-17, -92a, -126, -145 y -155 se han asociado con el calibre de la placa aterosclerótica, medido por tomografía computarizada coronaria 3.
Biomarcadores para el infarto agudo de miocardio (IAM) han sido muy útiles en la clínica y son muy específicos; por ejemplo, se han evidenciado mi-RNA marcadores como miR-208 propios del tejido cardíaco y los miR-1, miR-133a/b, miR-499, miR-663b y miR-1291 abundantes en miocitos estriados (3,51). Una ventaja adicional de estos marcadores sobre las troponinas específicas en el IAM es la rapidez con que se encuentran los mi-RNA en la sangre después del evento. En modelos murinos se ha observado un cambio en los niveles plasmáticos de estas moléculas entre 15 minutos y una hora después del evento isquémico
3.
Micro-RNA como biomarcadores potenciales en enfermedades hepáticas
El estudio de la organogénesis y desarrollo del hígado ha llevado a la identificación de los mi-RNA como moléculas importantes para el proceso, y es probable que también lo sean en la regulación y diferenciación de los distintos linajes celulares. La expresión de miR- 122 es específica para el hígado y la inhibición de su expresión en ratones ha llevado a la regulación negativa de enzimas del metabolismo del colesterol y de los lípidos. Se han identificado otros mi-RNA que se sobreexpresan en el hígado en formación, y debido a la relación entre los mi-RNA y el desarrollo hepático, se ha empezado a relacionar también diferentes miRNA con distintos estadios de enfermedades hepáticas. Los mi-RNA han surgido como biomarcadores potenciales para enfermedades hepáticas crónicas, aunque los estudios más recientes se han centrado en el papel de estas moléculas en la carcinogénesis hepática 51.
En una evaluación crítica sobre los mi-RNA como biomarcadores para enfermedades no neoplásicas llevada a cabo por Haider y colaboradores se analizaron 12 estudios relacionados con enfermedades hepáticas, en los cuales se reportaron en total 24 biomarcadores para lesión hepática. Los mi-RNA propuestos se validaron al determinar los patrones de expresión celular, considerando como un buen biomarcador el que se expresara en un tipo celular implicado en la enfermedad estudiada y que no presentara una expresión ubicua, es decir, que no se expresara ampliamente en distintos tipos celulares. Los autores determinaron que 6 de los 24 mi-RNA propuestos para enfermedades hepáticas (25 %) eran biomarcadores probables. De estos seis, miR-122, conocido como un mi-RNA específico para hígado, presentó una expresión elevada en 9 estudios separados. La mayoría de los mi-RNA reportados como posibles biomarcadores para enfermedad hepática fueron considerados por el grupo de investigadores como inespecíficos con patrones de expresión ubicuos 52.
Micro-RNA en hepatitis virales
Se ha demostrado que en infecciones virales se presenta un cambio en el perfil de expresión de los mi- RNA y en algunos estudios se ha reportado que algunos virus RNA interactúan directamente con los mi-RNA del hospedero y/o usan estas moléculas para aumentar su potencial de replicación (53). Se han estudiado varios mi-RNA en hepatitis virales y se ha encontrado que los niveles plasmáticos de miR-122 muestran muy buena correlación con la actividad necrosante e inflamatoria de VHB (54) y VHC (55). Se ha propuesto que miR-122 puede contribuir al tropismo hepático del VHC al acelerar la unión del RNA viral a los ribosomas y estimular la traducción del material genético del virus (56). Esto indicó que la inhibición de miR-122 podría ser una estrategia para bloquear la replicación del VHC, de lo que resultó un estudio basado en el silenciamiento terapéutico del mi-RNA en primates con infección crónica por VHC. Lanford y colaboradores utilizaron un oligonucleótido modificado para silenciar el mi-R122 y encontraron que con ello se logró suprimir la viremia sin generar resistencia viral ni efectos secundarios en los chimpancés 57. Esto demuestra que los mi-RNA no solo tienen potencial diagnóstico, sino además un importante potencial terapéutico.
Se han hecho estudios de mi-RNA en otras enfermedades hepáticas como el daño por drogas, la fibrosis, la hepatopatía alcohólica, la esteatosis no alcohólica y el daño hepático agudo. Entre los mi-RNA reportados, los datos más consistentes corresponden a incrementos en los niveles de expresión de mi-RNA-221 y mi- RNA-223 en pacientes con carcinoma hepatocelular, y con respecto a las enfermedades no neoplásicas, los datos más consistentes corresponden a elevación de miRNA-21 y -122 en pacientes con hepatitis crónica u otros tipos de lesiones hepáticas virales o tóxicas 58.
CONCLUSIÓN
Las limitaciones de los estudios y las dificultades técnicas de los métodos empleados para la extracción, purificación, procesamiento y análisis de la expresión de los mi-RNA se reflejan en resultados discordantes sobre los perfiles de expresión de estas moléculas en diferentes estudios. Aunque parece que algunos mi-RNA candidatos para biomarcadores presentan consistencia en su perfil de expresión, otros han sido reportados como posibles marcadores para más de una enfermedad, lo que implica una limitación con respecto a su especificidad y utilidad en el diagnóstico. Sin embargo, el potencial de los mi-RNA como biomarcadores es promisorio por lo que es pertinente hacer nuevos estudios que ahonden sobre su significado en procesos patológicos y su relevancia como posibles biomarcadores de los mismos. Además, pueden ser dianas de nuevas intervenciones terapéuticas ofreciendo alternativas para diferentes pacientes.

