










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de la Universidad Industrial de Santander. Salud
Print version ISSN 0121-0807On-line version ISSN 2145-8464
Rev. Univ. Ind. Santander. Salud vol.42 no.3 Bucaramanga Sept./Dec. 2010
La primera entrada en el laberinto en
cruz elevado como predictor
del nivel de ansiedad
Primera entrada como predictor de ansiedad
Cesar A. Celis1,2, Diana P. Martínez1, Carlos A. Conde1
1. Grupo de Neurociencias y Comportamiento UIS-UPB, Departamento de Ciencias Básicas, Facultad de Salud, Universidad Industrial de Santander, Bucaramanga, Colombia
2. CIMBIOS, Escuela de Física, Facultad de Ciencias, Universidad Industrial de Santander, Bucaramanga, Colombia
Correspondencia: Cesar A. Celis, Físico, Grupo de Neurociencias y Comportamiento UIS-UPB, Departamento de Ciencias Básicas, Facultad de Salud,
Universidad Industrial de Santander, Bucaramanga, Colombia Teléfono: 6344000 Ext.: 3126. E-mail: cescelis@gmail.com.
Recibido: 1 de diciembre de 2010- Aceptado: 20 de diciembre de 2010
RESUMEN
Introducción: Generalmente el análisis del comportamiento exploratorio de ratas en laberinto en cruz elevado es realizado a la población general sin llevarse a cabo ningún tipo de discriminación entre individuos en función de sus decisiones exploratorias. Considerando la toma de decisiones para entrar a una zona del laberinto como un conflicto entre motivación de explorar y aversión, el presente trabajo pretende evaluar la primera entrada a los brazos del laberinto como indicador del estado inicial de los niveles de motivación y aversión del entorno y en consecuencia, como indicador temprano de la evolución del comportamiento exploratorio. Materiales y métodos: Se evaluó el desempeño de ratas Wistar machos en laberinto en cruz elevado sometidas a una primera exposición, separándolas en dos grupos, de acuerdo a si su primera entrada fue en los brazos abiertos o en los brazos cerrados. Resultados: Aquellas ratas que primero entraron en los brazos abiertos obtuvieron valores significativamente mayores para el número de entradas y tiempo de permanencia en estos brazos en comparación con aquellas que entraron primero a los brazos cerrados; mientras que no se evidenció diferencias en el número de entradas en los brazos cerrados. Conclusión: La primera decisión del animal resulta predictiva del comportamiento durante los 5 minutos de exploración y permite discriminar las ratas entre dos niveles de ansiedad, sugiriéndose como variable discriminatoria el porcentaje de entradas a los brazos abiertos con punto de corte en 30,77%. Salud UIS 2010; 42: 220-228
Palabras Clave: Aversión, comportamiento exploratorio, motivación, toma de decisión, ratas Wistar
The susceptibility to infection by Leishmania panamensis and the
sensitivity to drugs differ according to the macrophage type
ABSTRACT
Introduction: Usually, the analysis of the rat exploratory behavior in elevated plus maze is carried out to the general population, without any discrimination between individuals as a function of their exploratory decisions. Considering the decision making to enter into a maze zone as a conflict between motivation to explore and aversion, the current work pretends to assess the first entry into the maze arms as indicator of the initial state of the motivation and aversion levels of the environment and therefore, as early indicator of the evolution of the exploratory behavior. Materials and methods: It was evaluated the performance of male Wistar rats in elevated plus maze subjected to a first exposure, divided into two groups, according to whether its first entry was made into the open or closed arms. Results: Those rats that entered first into the open arms obtained higher significant values in the number of entries and time spent in these arms with respect to those rats that entered first into the closed arms; but it was not obtained differences in the number of entries into the closed arms. Conclusion: The first animal choice predicts the behavior during the 5 minutes of exploration and allows to discriminate the rats between two anxiety levels, being suggested the percentage of entries in open arms as discriminatory variable with cut-off value in 30.77%. Salud UIS 2010; 42: 220-228
Keywords: Aversion, decision making, exploratory behavior, motivation, Wistar rats
INTRODUCCIÓN
El laberinto en cruz elevado (LCE) es reconocido como uno de los modelos eto-experimentales de ansiedad más utilizados para bioensayo de fármacos ansiolíticos1,2. Este modelo murino, basado en la libre exploración que despliegan los roedores frente a un ambiente novedoso, permite a través de su arquitectura, elegir entre dos pares de zonas distintas: un par de brazos potencialmente peligrosos (brazos abiertos) y un par de brazos seguros (brazos cerrados)3,4. Entre otros factores, la genética del animal expuesto al LCE determina en gran medida el repertorio exploratorio que este exhibe5,6,7. Sin embargo, a pesar de las diferencias numéricas entre parámetros, reportadas por los diferentes grupos de investigación alrededor del mundo8, una primera aproximación del estudio del comportamiento exploratorio de ratas en LCE ha mostrado cierta uniformidad en los patrones exploratorios caracterizados a través de parámetros comportamentales espacio-temporales (PCET) que definen de la forma más general posible el fenómeno. Dentro de estos patrones, la preferencia por los brazos cerrados es mantenida, aún en diferentes especies y cepas de experimentación5,6,7. Sin embargo, el avance en el entendimiento de este fenómeno exige ir cada vez más profundo en el análisis de la información, de modo que no solo las diferencias entre poblaciones, sino también entre individuos se vuelven relevantes9,10. Un primer paso en esa dirección exige que se realicen clasificaciones al interior de las cepas, separando las ratas en al menos dos grupos, uno con mayor nivel de ansiedad que el otro, esto en función de diferencias sugeridas o encontradas en alguno de estos PCET.
Varios podrían ser los criterios de clasificación y de igual forma el método de encontrarlos, ya sea siendo estos propuestos por el investigador o encontrados a través del uso de técnicas matemáticas11,9,12,13. Un reporte reciente ha mostrado la posibilidad de discriminar entre distintos estados comportamentales en la prueba de Campo Abierto en sesiones de 10 min, basados en el análisis del comportamiento del primer minuto, estableciendo la capacidad de discriminación de toda una sesión experimental a partir de eventos iniciales14. Partiendo de la conceptualización del LCE como un modelo animal de exploración libre donde la toma de decisiones para entrar a una zona del laberinto podría interpretarse como un conflicto entre motivación de explorar y aversión15, 16, sería factible pensar que la primera entrada a los brazos aludiría el estado inicial de los niveles de motivación y aversión del entorno y en consecuencia, la evolución del comportamiento exploratorio.
El presente trabajo se centró en determinar si la elección de la primera entrada a los brazos podría ser un indicativo del comportamiento exploratorio posterior observado durante los 5 min de libre exploración de ratas Wistar en LCE.
MATERIALES Y MÉTODOS
Animales
Los experimentos fueron realizados con 33 ratas Wistar machos con peso entre 190 y 250 g, provenientes del bioterio de la Universidad Industrial de Santander, alojadas en grupos de 6 animales por caja17, (50x30x15 cm), con comida y agua ad libitum, en un ciclo de luz-oscuridad de 12 horas, con las luces encendidas a las 7:00, temperatura promedio de 23±1°C, y humedad relativa de 66±5%. El periodo de habituación a la sala de experimentación fue de 8 días; 3 días antes de exponer los animales al LCE fueron manipulados por 1 minuto. Los experimentos se llevaron a cabo entre las 14:00 y las 18:00 horas. Todos los procedimientos realizados dentro de este proyecto se acogieron a la ley 84 y a la resolución 8430 de 1993 (Título V) y a la aprobación del comité de ética para proyectos de investigación de la Universidad Industrial de Santander.
Laberinto en cruz elevado
El dispositivo utilizado consistía en un laberinto compuesto de 4 brazos en forma de cruz elevados a una altura de 50 cm. Cada brazo media 12x50 cm y la zona central 12x12 cm. Dos brazos cerrados (BC), uno frente al otro, rodeados con paredes de madera de 40 cm de altura, y dos brazos abiertos (BA) con bordes de Plexiglás de 1 cm de altura.
Procedimiento
Las ratas fueron clasificadas en dos grupos en función de la primera entrada a los brazos del laberinto, el primer grupo llamado PEBA (n = 7) fueron aquellas ratas que entraron primero a los BA y el segundo llamado PEBC (n = 26), aquellas que entraron primero a los BC. El protocolo de experimentación en el LCE consistió en ubicar la rata en la zona central con la cabeza orientada hacia uno de los brazos cerrados, permitiendo la libre exploración durante 5 min. Para disminuir pistas odoríficas dejadas por las ratas, el laberinto fue limpiado con alcohol y algodones secos. La entrada a un brazo correspondía cuando todas las 4 patas estaban dentro de este.
Posterior a la ejecución de los experimentos, se calcularon los siguientes PCET: Entradas a los BA (EBA), entradas a los BC (EBC), tiempo de permanencia en los BA (TBA), tiempo de permanencia en los BC (TBC), porcentaje de entradas a los BA (%EBA = EBA*100/(EBA+EBC)), porcentaje de tiempo de permanencia en los BA (%TBA = TBA*100/(TBA+TBC)), tiempo promedio por entrada a los BA (TBAp = TBA/EBA), tiempo promedio por entrada a los BC (TBCp = TBC/EBC). También se calculó el numero de transiciones entre brazos para cada rata, obteniéndose cuatro tipos de transición: cuando la rata sale de un BC e inmediatamente después entra a un BC (BCBC), cuando sale de un BC y entra a un BA (BCBA), cuando sale de un BA y entra a un BC (BABC) y cuando sale de un BA y entra a otro (BABA). Estas transiciones componen las entradas a los brazos de manera que: EBC = BCBC + BABC; y EBA = BABA + BCBA. Como parámetro extra se analizó la latencia de la primera entrada a los brazos.
Análisis estadístico
Como análisis estadístico se realizó prueba t para los PCET y la latencia; y ANOVA de dos vías no paramétrica para las transiciones, con GRUPO x TRANSICION como factores; seguida de comparaciones múltiples con la prueba de Bonferroni (post hoc). Para evaluar la capacidad de discriminación de las variables se realizó un análisis del área bajo la curva ROC (Receiver Operating Characteristic), usando como diagnostico la primera entrada a los brazos (Dx0 = entró primero a los BA y Dx1= entró primero a los BC). Finalmente se realizaron correlaciones de Pearson con el fin de establecer el grado de relación entre los PCET16. Se estableció el nivel de significancia en p<0,05.
RESULTADOS
Se encontraron diferencias significativas para los valores de EBA (p = 0,005), TBA (p = 0,004), TBC (p = 0,002), %EBA (p < 0,001), %TBA (p = 0,004), donde las ratas PEBA, mostraron menor TBC y mayor EBA, TBA, %EBA y %TBA que las PEBC. EBC (p = 0,652), TBAp (p = 0,482) y TBCp (p = 0,495) no fueron significativamente diferentes entre grupos. Estos resultados se muestran en la (Figura 1).
ANOVA de dos vías evidenció un efecto para el factor TRANSICION (p < 0,001), pero no para el factor GRUPO (p = 0,141); también se encontró interacción entre los factores TRANSICION x GRUPO (p < 0,001). Se presentaron diferencias significativas entre grupos para las transiciones BCBC (p = 0,001), BCBA (p = 0,014) y BABC (p = 0,014), pero no para BABA (p = 0,196). No hubo diferencias significativas entre las transiciones BCBC y BCBA (p = 1) y BCBC y BABC (p = 1) para el grupo PEBA, mientras que el grupo PEBC presentó diferencias significativas entre BCBC y BCBA (p < 0,001), y BCBC y BABC (p < 0,001). Independiente del grupo, se presentaron diferencias significativas de BABA respecto a BCBC (p = 0,008 en PEBA, p < 0,001 en PEBC), pero no respecto a BCBA o BABC (p = 0,203 en PEBA, p = 0,138 en PEBC). Los resultados se muestran en la (Figura 2).
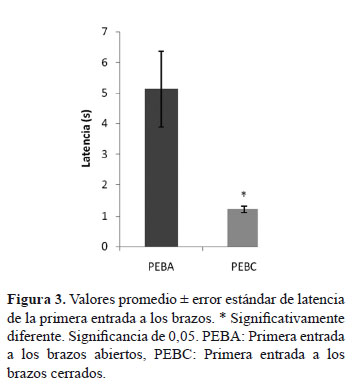
La latencia de la primera entrada a los brazos fue significativamente mayor para el grupo PEBA (p = 0,002). Los resultados se muestran en la (Figura 3). En la (Tabla 1) se consignaron los resultados obtenidos del análisis del área bajo la curva ROC para los PCET propuestos. Se encontró que la selección de la primera entrada a los brazos podría discriminar el comportamiento de las variables TBA, TBC, %TBA, EBA y %EBA después de 5 minutos de exploración libre, de modo que aquellas ratas que entraron primero a los BA se caracterizarían con valores de TBA mayores a 70,39 s, con sensibilidad 0,43 y especificidad 1,00; con valores de %TBA mayores a 27,15%, con sensibilidad 0,43 y especificidad 1,00; con valores de EBA mayores a 7,00, con sensibilidad 0,43 y especificidad 0,96; con valores de %EBA mayores de 30,77%, con sensibilidad 0,86 y especificidad 0,92. De igual forma, aquellas ratas que entraron primero a los BC se caracterizarían con valores de TBC mayores a 198,02 s, con sensibilidad 0,92 y especificidad 0,57. Se realizó un análisis de correlaciones entre los PCET utilizando todos los animales del experimento. La gran mayoría de parámetros obtuvieron correlaciones significativas, excepto EBC que solo correlacionó significativamente con TBCp, mientras que TBAp no correlacionó con EBA, EBC, %EBA y TBCp. Los resultados se consignaron en la (Tabla 2).
DISCUSIÓN
Las entradas a los brazos cerrados no fueron significativamente diferentes entre grupos, indicando que las diferencias en los parámetros EBA, TBA, TBC, %EBA, %TBA no son consecuencia de un cambio en la locomoción. Otros estudios han reportado que esta tendencia locomotora en los BC permanece invariante para animales expuestos a una primera sesión en LCE bajo iguales condiciones experimentales, a pesar de encontrarse diferencias significativas para los otros parámetros, en ratas de la cepa Wistar con diferentes niveles de ansiedad19, e incluso comparando entre ratas de distinta cepa5. La falta de correlación de EBC con todos los demás parámetros, excepto TBCp, concuerda con lo reportado sobre su relación con la actividad locomotora del animal, independiente de los estados de ansiedad de los individuos20.
TBAp y TBCp no fueron significativamente diferentes, lo que indica que el aumento en TBA para el grupo PEBA corresponde a un aumento en EBA. Estos resultados son consistentes con lo reportado en la literatura21. La disminución en TBC para el grupo PEBA podría corresponder a una leve disminución en TBCp, aunque esta no fuera lo suficientemente grande para notarse en el análisis de varianza, ya que EBC permaneció invariante.
Se sugiere como explicación de la preferencia de algunas ratas en entrar primero a los BA que a los BC, un análisis basado en el modelo computacional desarrollado por Salum et al. 15, en el cual se propone que la toma de decisión sobre a cual zona entrar es una competencia entre múltiples elecciones representables por valores numéricos, las cuales son ponderadas por la rata. Estos valores son obtenidos a partir de la ecuación w = M – A, donde w representa la tendencia por entrar, M la motivación de explorar y A la aversión asociada con una determinada zona. De acuerdo a esta ecuación, la elección de entrar primero a los BA, sería probablemente el resultado de un nivel de motivación por explorar estas zonas mayor que la aversión percibida por el animal y mayor que la motivación por explorar los BC en un primer instante. Lo anterior expresado matemáticamente corresponde a: MBA > ABA y MBA > MBC. Además, si esta motivación más alta se mantiene daría como consecuencia un mayor EBA durante la sesión experimental, sin entrar a modificar necesariamente TBAp, tal como se evidencia en los resultados.
La variación en EBA, la invarianza de TBAp, y la falta de correlación entre estos parámetros, sugiere la presencia de dos diferentes mecanismos aversivos que al parecer actúan de modo independiente: el primero influye sobre la probabilidad de entrar a la zona aversiva, en este caso, los BA, y se caracteriza a través del parámetro %EBA; mientras que la segunda influye sobre el tiempo en que se tolera el estimulo aversivo, en este caso, la permanencia en los BA caracterizada a través del parámetro TBAp. De hecho, TBAp no correlaciona con ninguna variable exploratoria (EBA, EBC, %EBA), sugiriendo una independencia de esta variable con la actividad locomotora y el hasta ahora reportado estado de ansiedad del animal. Posiblemente, TBAp se relacione con un segundo estado de ansiedad determinado por la resistencia del animal a la presencia del estimulo aversivo, medida por el lapso que permanece en los BA.
De manera contraria a lo que se pensaría, basados en lo expuesto anteriormente, TBCp si correlaciona con las variables exploratorias (EBA, EBC, %EBA), lo cual, junto con la falta de correlación entre TBCp y TBAp, sugiere una diferencia en los mecanismos que producen los tiempos de permanencia en los BC respecto a los BA.
La diferencia significativa entre las transiciones BCBC y BCBA para el grupo PEBC, indica que la rata cada vez que sale de un BC y toma la decisión de si entrar a un BA o a un BC, lo hace de modo que prefiere los BC sobre los BA; mientras que para el grupo PEBA, la selección entre brazos es más homogénea16. Asimismo, el análisis cuantitativo ilustrado en la Fig. 2 sugiere una compensación entre las transiciones BCBA y BCBC, de modo que cuando BCBA aumenta, BCBC disminuye y viceversa, por lo cual se mantiene la invarianza en EBC, tal como se reportó en los resultados.
Las latencias de la primera entrada a los brazos, mostraron que las ratas del grupo PEBC deciden mucho más rápido a cual brazo entrar que las del grupo PEBA. Esta rapidez en la decisión pone en evidencia una ausencia de conflicto en las ratas del grupo PEBC, producto de su baja probabilidad de entrar a los BA, dando como única opción la entrada a los BC, mientras que las ratas del grupo PEBA, al presentar una mayor preferencia a arriesgarse a explorar los BA, sin dejar de lado la atracción por permanecer en una zona segura (BC), les toma un poco más de tiempo decidir a cuál de los brazos entrar primero, dando como consecuencia, la elección de los BA.
Además de la utilidad que presentan estos resultados para el entendimiento de los fenómenos presentes durante la exploración de ratas en LCE, se propone la aplicación del conocimiento desarrollado a partir de este análisis para retroalimentar los modelos computacionales propuestos por Salum et al. 15 y Giddings y Stephenson 22, en los cuales la elección inicial de entrada a los brazos no parece influir en el posterior desarrollo de la exploración. Los autores de este articulo consideran indispensable para el avance del conocimiento en esta área el desarrollo y mejoramiento de modelos teóricos que permitan condensar y conceptualizar los datos experimentales, siendo parte de estos modelos los desarrollos computacionales15,22,23.
Finalmente, es posible concluir que los parámetros EBA y %EBA permiten atribuir diferencias entre los grupos PEBA y PEBC para el nivel de riesgo de entrar a una zona aversiva, siendo menor para el grupo PEBA, mas no para el tiempo de permanencia en que ese estimulo se tolere, como lo sugiere TBAp. De igual forma, a partir de las diferencias significativas para %EBA y %TBA, se concluye que las ratas que entraron primero a los BA (PEBA) presentan un nivel de ansiedad menor que aquellas que entraron primero a los BC (PEBC). Luego, la primera decisión de entrar a los brazos del laberinto es un evento predictor de la evolución exploratoria de la rata cuando esta es expuesta al LCE por 5 minutos, y que permite discriminar entre ratas con distintos niveles de ansiedad, sugiriéndose como variable discriminatoria %EBA, basándose en su alto valor ROC (0,90), sensibilidad (0,86), especificidad (0,92) y porcentaje de bien clasificados (90,91%)18, la cual permite predecir que los animales que primero entren a los BA obtendrán valores superiores a 30,77% para esta variable, mientras que los animales que entren primero a los BC obtendrán valores inferiores a este. Es por esto que se propone la primera entrada a los brazos como un método más, entre los actuales ya explorados, para la clasificación de ratas en función de las diferencias individuales bajo dos niveles de ansiedad, uno alto y otro bajo9,10,11,12,13.
AGRADECIMIENTOS
El presente trabajo fue realizado gracias al apoyo de la Vicerrectoría de Investigación y Extensión en su programa para la consolidación de los grupos de investigación código 5635 y a la Dirección de Investigación y Extensión Facultad de Salud de la Universidad Industrial de Santander
CONFLICTOS DE INTERÉS
Los autores manifiestan no tener ningún conflicto de intereses al realizar este trabajo.
REFERENCIAS
1. Pellow S, File SE. Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: A novel test of anxiety in the rat. Pharmacol Biochem Behav 1986; 24: 525-529. [ Links ]
2. Pellow S. Anxiolytic and anxiogenic drug effects in a novel test of anxiety: are exploratory models of anxiety in rodents valid?. Methods Fin. Exp Clin Pharmacol 1986; 8: 557-565. [ Links ]
3. Pellow S, Chopin P, File SE, Briley, M. Validation of open: closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J Neurosci Methods 1985; 14: 149-167. [ Links ]
4. Schwarting RKW, Borta A. Analysis of behavioral asymmetries in the elevated plus-maze and in the T-maze. J Neurosci Methods 2005; 14:251-260. [ Links ]
5. Izídio GS, Spricigo JrL, Ramos A. Genetic differences in the elevated plus-maze persist after first exposure of inbred rats to the test apparatus. Behav Proces 2005; 68: 129-134 [ Links ]
6. Ramos A, Berton O, Mormède P, Chaouloff F. A multiple-test study of anxiety-related behaviours in six inbred rat strains. Behav Brain Res 1997; 85: 57-69. [ Links ]
7. Shepard JD, Myers DA. Strain differences in anxiety-like behaviour: Association with corticotrophin-releasing factor. Behav Brain Res 2008; 186: 239-245. [ Links ]
8. Hogg S. A review of the validity and variability of the elevated plus-maze as an animal model of anxiety. Pharmacol Biochem Behav 1996; 54: 21-30. [ Links ]
9. Liebsch G, Montkowski A, Holsboer F, Landgraf R. Behavioural profiles of two Wistar rat lines selectively bred for high or low anxiety-related behavior. Behav Brain Res 1998; 94: 301-310. [ Links ]
10. Henniger M, Ohl F, Hölter SM, WeiBenbacher P, Toschi N, Lörscher P, et al. Unconditioned anxiety and social behaviour in two rat lines selectively bred for high and low anxiety-related behaviour. Behav Brain Res 2000; 111: 153-163. [ Links ]
11. Henriques AS, Araújo AFR, Morato S. Clustering Exploratory Activity in an Elevated Plus-Maze with Neural Networks. IEEE-INNS-ENNS International Joint Conference on Neural Networks 2000; 4: 4017. [ Links ]
12. Borta A, Wöhr M, Schwarting, RKW. Rat ultrasonic vocalization in aversively motivated situations and the role of individual differences in anxiety-related behavior. Behav Brain Res 2006; 166: 271-280. [ Links ]
13. Ho YJ, Eichendorff J, Schwarting RKW. Individual response profiles of male Wistar rats in animal models for anxiety and depression. Behav Brain Res 2002; 136: 1-12. [ Links ]
14. Brenes JC, Padilla M, Fornaguera J. A detailed analysis of open-field habituation and behavioral and neurochemical antidepressant-like effects in postweaning enriched rats. Behav Brain Res 2009; 197: 125-137. [ Links ]
15. Salum C, Morato S, Roque-da-silva AC. Anxiety-like behavior in rats: a computational model. Neural Networks 2000; 13: 21-29. [ Links ]
16. Conde C, Nieto J, Orozco LC. Análisis comportamental de ratas expuestas al laberinto en cruz elevado bajo dos horarios de alimentación. Salud UIS 2008; 40: 83-91. [ Links ]
17. Botelho S, Estanislau C, Morato S. Effects of under- and overcrowding on exploratory behavior in the elevated plus-maze. Behav Processes 2007; 74: 357-362. [ Links ]
18. Burgueño MJ, Garcia-Bastos JL, Gonzalez-Buitrago JM. Las curvas ROC en la evaluación de las pruebas diagnósticas. Med Clin (Barc) 1995; 104: 661-670. [ Links ]
19. Salomé N, Viltart O, Darnaudery M, Salchner P, Singewald N, Landgraf R, et al. Reliability of high and low anxiety-related behavior: influence of laboratory environment and multifactorial analysis. Behav Brain Res 2002; 136: 227-237. [ Links ]
20. Rodgers RJ, Johnson NJT. Factor analysis of spatiotemporal and ethological measures in the murine plus-maze test of anxiety. Pharmacol Biochem Behav 1995; 52: 297-303. [ Links ]
21. Garcia AMB, Cardenas FP, Morato S. Effect of different illumination levels on rat behavior in the elevated plus-maze. Physiol Behav 2005; 85: 265-270. [ Links ]
22. Giddings J, Stephenson P. Modeling the behaviour of rats in an elevated plus maze (Bachelor's thesis). Acadia University 2002. [ Links ]
23. Miranda DA, Conde CA, Celis CA, Corzo SP. Modelado del comportamiento de ratas en laberinto en cruz elevado basado en redes neuronales artificiales. Rev Col Fis 2009; 41: 406-408. [ Links ]

