










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista de Ciencias
Print version ISSN 0121-1935
rev. cienc. vol.18 no.2 Cali July/Dec. 2014
Food Habits for a Population of Eleutherodactylus johnstonei in an Area on the Bank of River Melendez, South of Cali, Valle del Cauca
Hábitos alimenticios de una población de Eleutherodactylus johnstonei en una zona a las orillas del río Meléndez, al sur de Cali, Valle del Cauca
Oscar Darío Hernández Córdoba
Grupo de Investigación Laboratorio de Herpetología, Universidad del Valle, Cali - Colombia
E-mail: oscarhernandezcordoba@gmail.com
Received: June 13, 2013
Accepted: November 18, 2014
Abstract
Due to the ecological importance of knowing the variation in the diet of a species along its geographic range, the objective of this work was to establish the diet of Eleutherodactylus jhonstonei in a population located at south in the city of Cali. A total of 43 individuals were captured, and the food preferences and niche breadth were calculated by stomach content analysis. Finally, diet structure was compared between sexes and showed significant differences (p=0,017). Additionally, It was found that E. jhonstonei is a generalist species (Niche breadth=0,6) whose diet depends on the abundance of prey in the environment withouth showing a food preference, this is a reason why the species has become a successful colonizer.
Keywords: food preference, diet, niche breadth, stomach content, trophic ecology.
Resumen
Debido a la importancia ecológica que implica el conocer las diferentes dietas de una especie dentro de su rango de distribución, este trabajo consistió en establecer la dieta de una población de Eleutherodactylus jhonstonei al sur de Cali, 44 individuos fueron capturados, analizando el contenido estomacal se calculó el grado de preferencia alimenticia, la amplitud de nicho y se comparó la estructura de la dieta entre los sexos, la cual tuvo diferencias significativas (p=0,017). Se encontró que E. jhonstonei es una especie generalista (amplitud de nicho = 0,6) cuya dieta depende de la abundancia de presas en los ambientes mas no de una preferencia marcada, por lo que se ha convertido en un colonizador exitoso.
Palabras clave: amplitud de nicho, contenido estomacal, dieta, ecología trófica, grado de preferencia alimenticia.
1. Introducción
Eleutherodactylus johnstonei es una rana de la familia Eleutherodactylidae, especie nativa de las Antillas menores que ha sido introducida en el Caribe y el norte de Suramérica (Schwartz, 1967). En Colombia, esta especie se ha reportado en la ciudad de Barranquilla, Cartagena, Cali y Bucaramanga (Acosta-Galvis, 2000; Kaiseret al., 2002; Ortega et al., 2005; Ortega et al., 2005) y ocupa el 10.5% de la extensión total del país (Urbina-Cardona y Castro, 2011) sin embargo, se desconocen los impactos sobre las especies nativas (Corredor et al., 2010) a pesar de que actualmente ha llegado a habitar zonas rurales como Dapa (Yumbo, Valle del Cauca; Obs. Pers.) y se plantea que puede entrar a competir con especies nativas como Pristimantis palmeri, ya que las dos especies tienen una preferencia de hábitat similar y que tienen vocalizaciones correlacionadas en un 70% (Gutierres-Zuñiga, 2013). E. jhonsonei es una especie de tamaño pequeño donde los machos pueden medir desde 17 a 25 mm de longitud y las hembras de 17 a 35 mm. (Wang, 2000). Se ha descrito como una especie que habita en zonas de alta intervención en los arbustos o en el suelo (Hedges, 2008), de hecho se encuentra asociada a la vegetación baja de los jardines de las ciudades donde se ha introducido deliberadamente y ha llegado a ser un problema por la intensidad de su canto que puede ser molesto en las áreas urbanas (Corredor, 2010) puesto que su actividad acústica se prolonga desde el inicio de la noche hasta las primeras horas de la madrugada.
Se conoce que la morfología, tácticas de forrajeo y comportamiento antipredador de las ranas covarían con la dieta (Toft, 1980); por lo que se hace importante el tener información de las dietas de diferentes partes de su rango de distribución para entender las adaptaciones ecológicas dentro de los grupos de anuros (Ovaska, 1991). Los hábitos alimenticios de E. jhonstonei han sido estudiados en varias poblaciones, una de estas en Barbados (Ovaska, 1991) donde se encontró un 79.6% de estómagos con hormigas, seguido de un 36.3% de estómagos que contenían colémbolos, además el 51.4% de las presas consumidas, fueron hormigas. Otra población de E. johnstonei estudiada en Bucaramanga (Ortega et al., 2005) donde calcularon el índice de importancia relativa para cada presa, reporta los dos más altos valores de dicho índice para la familia Formicidae, seguido de Diptera.
La relevancia de este aporte radica en la necesidad de conocer los hábitos alimenticios de la especie invasora E. jhonstonei en una población al suroccidente Colombiano con el fin de evaluar si existe algún tipo de preferencia y comparar con las poblaciones de diferentes lugares en su rango de distribución, reuniendo información que permita elucidar aspectos de la ecología que arrojen respuestas del por qué esta especie es un colonizador altamente exitoso.
2. Materiales y métodos
En octubre de 2010 se realizaron búsquedas a las orillas del río Meléndez, al sur de la ciudad de Cali, una zona urbana donde la vegetación predominante son pastos y algunos arbustos. El muestreo fue realizado en horas de la noche durante el período de más actividad acústica (19-21 pm), con un esfuerzo total de 15 horas hombre sobre transectos libres limitados por el tiempo. Se capturaron un total de 44 individuos, los cuales fueron sacrificados con etanol (30%) inmediatamente después de la captura, con el fin de interrumpir los procesos digestivos. Posteriormente fijaron en formol al 70% y se conservaron en alcohol al 70%.
Los individuos fueron llevados a los laboratorios de Biología en la Universidad del Valle, donde se midieron (SLV), se sexaron y se hizo la extracción de los tractos digestivos para analizar el contenido estomacal clasificándolo en OTU's (Unidades taxonómicas operacionales).
Posteriormente, se realizó un muestreo de entomofauna en la zona donde los individuos fueron capturados a la misma hora, más no simultáneamente con la colecta de ranas, se utilizaron trampas de caída, embudo de berlesse y jameo, para así obtener una mayor representatividad de la zona; esto se realizó con el fin de tener una idea de la riqueza y abundancia de entomofauna del lugar y un punto de comparación a la hora de la identificación de las partes de individuos en los tractos digestivos analizados.
Se realizó una curva de acumulación y se calculó el indicador Chao2 con el fin de evaluar la efectividad del muestreo, se escogió este indicador debido a que es menos sesgado para muestras pequeñas, se calcularon las frecuencias de aparición de cada grupo alimentario y el grado de preferencia alimenticia DFP (Braga, 1999), cuando un estomago presentó más de un grupo, el valor asignado fue 3 al más abundante, 2 al segundo más abundante y 1 al menos abundante. Posteriormente se calculó DFP = S (i) /N; donde S(i) es la suma de valores dados a un grupo de alimento i en el estómago y N es el número total de estómagos.
Para comparar la estructura de la dieta entre machos y hembras se realizó una prueba de t de diversidad Shannon, también se calculó el índice de Bray-Curtis.
Se calculó la amplitud de nicho mediante el índice de Levins (Krebs, 1989) por medio de la siguiente fórmula: Ba=(B-1)/(n-1).
Donde BA es el índice de Levins para (n) ítems y B = 1 / Σ pi2, siendo pi la proporción de un ítem en el total de la muestra.
3. Resultados y discusión
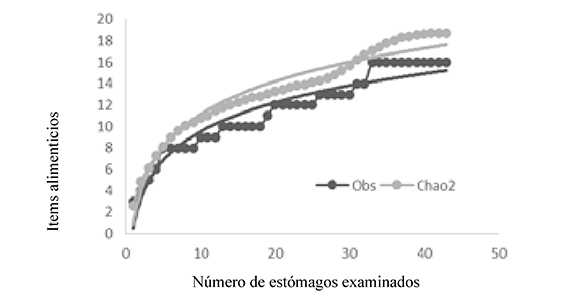
Para comprobar que la muestra fue representativa, se realizó una curva de acumulación, donde se puede observar que a partir del estómago 30, la curva se empieza a estabilizar; indicando a este como la cantidad de muestreo mínima, el muestreo presento un Chao2 del 79%, lo que quiere decir que pueden aparecer más ítems alimenticios de los detectados.
Se capturaron 44 individuos de E. Jhonstonei adultos (21 hembras y 23 machos), el tamaño medio (SLV) fue de 22 mm para las hembras y de 19.8 para los machos. Al ser los individuos sacrificados inmediatamente después de la colecta, en el análisis de contenidos estomacales se encontró que el 95% de los individuos presentó algún tipo de presa o contenido, de estos se obtuvo un espectro trófico de 17 categorías (OTU's) con un total de 179 ítems donde, 76 (43%) fueron encontrados en las hembras y 106 (57%) ítems en los machos (Ver gráfico 1). Sumado al espectro trófico, en todos los estómagos analizados se encontró un contenido mineral y vegetal considerable, sin embargo no fue tenido en cuenta para este análisis.
Gráfico 1. Curva de acumulación, en el eje Y se encuentran las categorías nuevas del espectro trófico, mientras que el eje X representa el número de estómagos
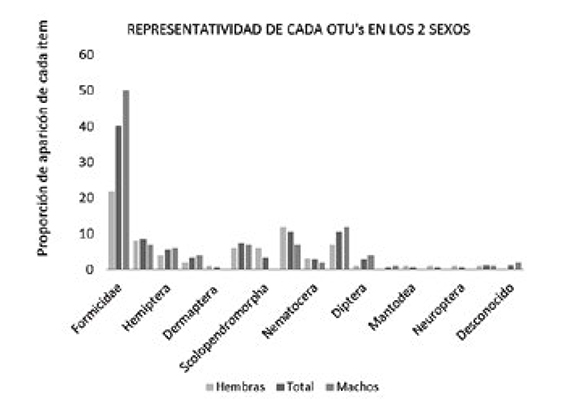
La estructura dietaría presentó una similitud del 61% entre machos y hembras, siguiendo el índice de Bray-Curtis; por otra parte, la prueba de diversidad t de Shannon mostró que hay diferencias significativas entre los sexos (p=0,017), las hembras tuvieron un mayor espectro trófico con 15 taxa vs. 12 taxa en los machos, esto posiblemente se deben a las diferencias de percha que tienen los distintos sexos de estas ranas, diferencias ya reportadas por Ortega et al. (2005). Al igual que en la población del presente trabajo, donde se encontró que los machos perchan en plantas arbustivas a más de 25 cm del suelo, mientras que la mayoría de las hembras se encontraron en perchas más bajas o en el suelo, sin embargo hay que tener en cuenta que los machos pueden descender al suelo donde cuidan las nidadas (Ortega et al., 2005). Las consecuencias de estas diferencias se ven reflejadas en que las hembras al estar más cerca del suelo pueden obtener presas que se movilizan generalmente por este como lo son Dermáptera y Scolopendromorpha. Se observó también una variación en la proporción de captura de hormigas entre los machos y las hembras, siendo más bajo en las hembras (Gráfico 2).
Gráfico 2. En el eje Y, se encuentran los porcentajes de aparición de cada categoría (taxa) para el total de los indivi- duos analizados, porcentajes de aparición solo en hembras y porcentajes de aparición solo en machos
Como se puede ver en los resultados mostrados en la Tabla 1. Formicidae estuvo presente en el 93% de los estómagos, representando el 40% del total de las presas encontradas, los valores más altos siguientes fueron los encontrados para Drosophila y Aranae, los cuales tuvieron presencia tan solo en el 26% de los estómagos y representan el 10% del total de las presas cada uno. Al comparar estos resultados con lo obtenido por Ovaska (1991) y Ortega et al. (2005), en poblaciones de Barbados y Bucaramanga, respectivamente, se puede notar una reducción en la frecuencia de aparición de hormigas en los tractos digestivos, pues en el primer trabajo se registró un 54%, y en el segundo se registró que el 73% de las presas pertenecían a la familia Formicidae. De cualquier forma los dos estudios de referencia y la aproximación actual presentan a las hormigas como la presa más abundante para E. jhonstonei. Las hormigas han sido reportadas como presa principal de una gran variedad de especies pertenecientes antiguamente al género Eleuthuerodactylus (Lavigne & Drewry, 1970; Duellman, 1978), de la misma manera, la importancia de las hormigas como ítem alimenticio no está limitada a la dieta de estas especies, pues muchas especies de anuros de tamaño pequeño tienen en su dieta a las hormigas como componente principal, de hecho se cataloga a la mayoría de dendrobatidos como especialistas para alimentarse de hormigas. Toft (1980), en su trabajo divide las especies como especialistas de hormigas, generalistas y como no comedores de hormigas, en el que los dendrobatidos son clasificados como especialistas de hormigas y los Eleutherodactylus son clasificados como generalistas; que por otra parte, tienen una estrategia de forrajeo pasiva, de quieto y espera o sit-and wait, diferente a la estrategia de captura activa que presentan los dendrobatidos (Toft, 1959).
Tabla 1. Espectro trófico encontrado en los estómagos categorizado en 16 OTU's y una categoría desconocida, para cada una se calculó la proporción con relación al total de estómagos (columna 1), el porcentaje de aparición en el total de estómagos analizados (Columna 2) y el grado de preferencia alimenticia (DFO valores menores a 1 son tomados como ocasionales y valores entre 1 y 2 son tomados como secundarios*)
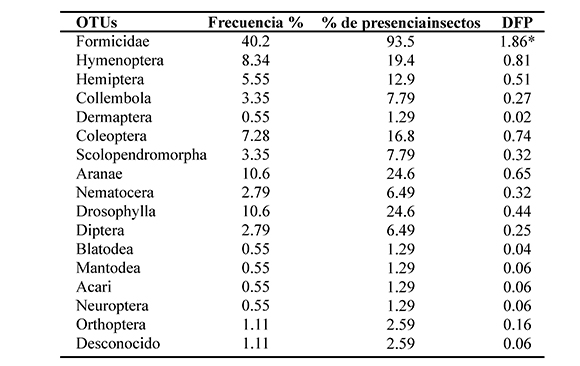
En este trabajo se calculó el grado de preferencia alimenticia DFP (Braga, 1999), el resultado no muestra valores que indiquen preferencia por presa alguna; según este indicador, todos los valores que estén entre el rango 0<DFP<1 se toman como presas como ocasionales, y los valores de 1<DFP<2 como secundarios, 2<DFP<3 como preferencial y 3<DFP<4 como altamente preferencial. Como se puede observar en los resultados (Tabla 1), todos los valores aplican para que las presas sean clasificadas como ocasionales, con la única excepción de Formicidae, que tuvo un valor DFP= 1,8 quedando en la categoría de presa secundaria.
En la población de estudio no se detectó ningún grado de preferencia alimenticia por alguna de las categorías encontradas, esto es muestra de que E. jhonstonei es un carnívoro generalista y al tener un espectro trófico amplio, lo es también su nicho (valor Ba=0,6 calificándolo como generalista); por lo que la gran abundancia de hormigas encontradas en su dieta, está relacionada con la abundancia en el ambiente que con algún grado de preferencia, pues existe una relación entre la abundancia de la presa en el ambiente y en la dieta de los anuros (Turner, 1959), de hecho al realizar el muestreo de entomofauna en la zona de estudio, Formicidae fue el grupo más abundante con casi el 80% de la muestra, cabe resaltar, que no todas las categorías encontradas en los estómagos estuvieron representadas en este muestreo, posiblemente a que hace falta incluir más métodos de captura que cubran los diferentes taxa, que pueden ser ítems alimenticios para los individuos de la población de estudio.
Los tamaños de la mayoría de presas fueron uniformes, con excepción de algunos animales como escolopendras y coleópteros, la relación volumétrica de la presa, con respecto al SLV y MW (ancho de la boca), no se realizó para esta población; sin embargo, es conocido que para muchos anuros y para esta especie reportado por Ovaska (1991) y Ortega et al. (2005), el tamaño corporal está relacionado directamente con el consumo de presas más grandes, es decir, ranas más grandes consumen presas más grandes. Una posible razón al por que las hembras consumieron presas excepcionalmente grandes, como las mencionadas anteriormente.
E. jhonstonei es una especie oportunista, cuya dieta depende de la abundancia de presas en el ambiente: la población de estudio no presentó preferencia alimenticia por ningún ítem. De igual forma, la amplitud de nicho calculada la califica como generalista, lo que concuerda con lo encontrado por Urbina-Cardona y Castro (2012), que propone a E. jhonstonei como una de las especies invasoras con amplitud de nicho más alto (10.5% de la extensión nacional) y pronostica incremento en su distribución conforme al crecimiento urbano y al desarrollo de la infraestructura vial.
Un punto relevante reportado en el presente trabajo, es que las ya reportadas diferencias en la percha según el sexo en esta especie, significan también diferencias en la consecución diferencial de presas para cada sexo; igualmente sucede con las diferencias en el tamaño de estos individuos, pues un individuo más grande, tendrá acceso a un mayor espectro trófico. La poca especialización en la dieta, reportada para varias poblaciones de esta especie, sumada a otros aspectos como: el ser una especie asociada a los asentamientos humanos, hacen de esta rana un colonizador exitoso y explican el que su distribución geográfica esté en aumento, incluso en las áreas rurales cercanas a la ciudad donde puede empezar a competir con especies nativas.
Agradecimientos
Agradezco al grupo GERA por la ayuda en la fase de campo de este trabajo, a Fernando Castro por sus aportes y al departamento de Biología de la Universidad del Valle, donde realicé la parte de laboratorio. Agradezco también a los revisores anónimos que enriquecieron con sus valiosos comentarios este manuscrito.
Referencias
Acosta-Galvis, A. (2000). Ranas, salamandras y caecilias (Tetrapoda: Amphibia) de Colombia. Biota Colombiana, 1, 289-319. [ Links ]
Braga, F.M.S. (1999). O grau de preferência alimentar: método qualitativo e quantitativo para o estudo do conteúdo estomacal de peixes. Acta Scientiarum, Maringá, 21, 291-295. [ Links ]
Corredor, G., Belasquez, B., Velasco, J., Castro, F., Bolívar, W., y Salazar, M. (2010). Plan de acción para la conservación de anfibios del Departamento del Valle del Cauca. Cali, Colombia: Corporación Autónoma Regional del Valle del Cauca. [ Links ]
Duellman, W. E. (1978). The biology of an equatorial herpetofauna in Amazonian Ecuador (n. 65). Lawrence, USA: University of Kansas. [ Links ]
Gutierres-Zuñiga, J., Castaño, S., & Castro, F. (2013). Potencial efecto negativo del canto entre Eleutherodactylus johnstonei (Eleutherodactylidae: Amphibia) y Pristimantis palmeri (Craugastoridae: Amphibia). Tesis de pregrado, Universidad del Valle. Cali, Colombia. [ Links ]
Hedges, B., Ibéné, B., Koenig, S., La Marca, E., Ibáñez, R., & Hardy, J., (2008). Eleutherodactylus johnstonei. In: IUCN 2010. IUCN Red List of Threatened Species. Recovered 12/10/2010 www.iucnredlist.org [ Links ]
Kaiser, H., Barrio-Amoros, C., Trujillo, J.D., & Lynch, J.D. (2002). Expansion of Eleutherodactylus johnstonei in northern South America: rapid dispersal through human interactions. Herpetol. Rev., 33, 290-294. [ Links ]
Krebs, C.J. (1989). Ecological Methodology. New York, USA: Harper e Row, publishers. [ Links ]
Lavigne, R.J., & Drewry, G. (1970). Feeding behavior of the frogs and lizards in the tropical wet forest preliminary report. In, R. G. Clements, G. E. Drewry, & R. J. Lavigne (Eds.). The Rain Forest Project Annual Report 147. (pp. 64-73). San Juan, Puerto Rico. [ Links ]
Ortega, J., Serrano, V.H., & Ramírez-Pinilla, M.P. (2005). Diet composition and microhabitat of Eleutherodactylus johnstonei in an Introduced Population at Bucaramanga City, Colombia. Herpetological Review, 36 (3), 238-241. [ Links ]
Ortega, J., Serrano, V.H., & Ramírez-Pinilla, M.P. (2005). Reproduction of an introduced population of Eleutherodactylus johnstonei at Bucaramanga. Copeia, 3, 642-648. [ Links ]
Ovaska, K. (1991). Diet of the frog Eleutherodactylus johnstonei in Barbados. West Indies. J. Herpetol, 25, 486-488. [ Links ]
Schwartz, A. (1967). Frogs of the genus Eleutherodactylus in the Lesser Antilles. Studies on the Fauna of Curacao and other Caribbean Islands, 24 (1), 1-62. [ Links ]
Toft, C. (1980). Feeding ecology of thirteen syntopic species of anurans in a seasonal tropical environment. Oecology (Berl.)., 45, 131-141. [ Links ]
Toft, C. (1981). Ecology of Panamanian Litter anurans: Patterns in Diet and Foraging Mode. Journal of Herpetology, 15(2), 139-144. [ Links ]
Turner, F.B. (1959). An analysis of the feeding habits of Rana pretiosa in Yellowstone Park (Wyoming). Amer. Midl. Nnt., 61, 403-413. [ Links ]
Urbina-Cardona, J.N., y Castro, F. (2011). Distribución del nicho ecológico actual y futuro de anfibios y reptiles invasores: Competencias en las Corporaciones Autónomas Regionales y de Desarrollo Sostenible en Colombia. 180-188. En, O. Vargas-Ríos, & S.P. Reyes (Eds). La restauración ecológica en la práctica. Memorias del I Congreso Colombiano de Restauración Ecológica & II Simposio Colombiano de Experiencias en Restauración Ecológica. Bogotá, Colombia: Grupo de Restauración Ecológica, Universidad Nacional de Colombia - GREUNAL. [ Links ]
Wang, C. (2000). Chihwang AT uclink.berkeley.edu, AmphibiaWeb. Reovered 19/03/2010 www.amphibiaweb.com [ Links ]

Revista de Ciencias por Universidad del Valle se encuentra bajo una licencia Creative Commons Reconocimiento 4.0.

