










Services on Demand
Journal
Article
 text in
text in  Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad de Odontología Universidad de Antioquia
Print version ISSN 0121-246X
Rev Fac Odontol Univ Antioq vol.24 no.1 Medellín July/Dec. 2012
ARTÍCULOS ORIGINALES DERIVADOS DE INVESTIGACIÓN
Análisis molecular de Sonic hedgehog (Shh) en la etiología de la fisura labiopalatina no sindrómica en tríos caso-progenitores chilenos
Ramiro J. Rincón R.1; José Suazo2; Rafael Blanco C.2
1 Profesor Facultad de Odontología, Universidad de Antioquia, Medellín, Colombia
2 Profesores Facultad de Medicina, Universidad de Chile, Santiago, Chile
RECIBIDO: FEBRERO 28/2012-ACEPTADO: JUNIO 12/2012
Rincón RJ, Suazo J, Blanco R. Análisis molecular de Sonic hedgehog (Shh) en la etiología de la fisura labiopalatina no sindrómica en tríos caso-progenitores chilenos. Rev Fac Odontol Univ Antioq 2012; 24(1): 110-120.
RESUMEN
INTRODUCCIÓN: la fisura labio palatina no sindrómica, NSCLP (del inglés Nonsyndromic cleft lip and palate) es una de las malformaciones congénitas más frecuentes tanto en Chile como en el resto del mundo. Presenta un modo de herencia multifactorial, en la que interactúan varios genes y el medio ambiente. Evidencias experimentales han demostrado la participación de Sonic hedgedhog (Shh) en la migración de las células de la cresta neural, en la transformación epitelio-mesénquima y en la formación de las estructuras medias craneofaciales durante el desarrollo embrionario, es probable una asociación entre variantes de Shh y la NSCLP.
MÉTODOS: el objetivo de este trabajo fue evaluar las regiones exónicas e intrónicas adyacentes de Shh, en una muestra de 150 tríos caso-progenitores para hallar la asociación con NSCLP. Se utilizó el método PCR-RFLP para determinar la presencia de heterodúplex. Luego, se utilizó la técnica de Conformation Sensitive Gel Electrophoresis (CSGE) para ver la distorsión del ADN en los heterodúplex. Como método alternativo, se hizo un análisis de polimorfismos de un solo nucleótido (del inglés single-nucleotide polymorphism SNP) para determinar asociación entre NSCLP y Shh, para lo cual se utilizaron los SNP: rs1233555 y rs1233556, ubicados en el primer intrón de Shh.
RESULTADOS: el objetivo de este trabajo fue evaluar las regiones exónicas e intrónicas adyacentes de Shh, en una muestra de 150 tríos caso-progenitores para hallar la asociación con NSCLP. Se utilizó el método PCR-RFLP para determinar la presencia de heterodúplex. Luego, se utilizó la técnica de Conformation Sensitive Gel Electrophoresis (CSGE) para ver la distorsión del ADN en los heterodúplex. Como método alternativo, se hizo un análisis de polimorfismos de un solo nucleótido (del inglés single-nucleotide polymorphism SNP) para determinar asociación entre NSCLP y Shh, para lo cual se utilizaron los SNP: rs1233555 y rs1233556, ubicados en el primer intrón de Shh.
CONCLUSIÓN: la no asociación puede deberse a que la frecuencia de distribución de los SNP en la población chilena es diferente a la de las poblaciones referidas, o a que el número de SNP analizados fue insuficiente, o la no inclusión para el análisis de otras regiones de Shh.
Palabras clave: labio y paladar hendido no sindrómico, Sonic hedgedhog, SNP.
INTRODUCCIÓN
La fisura labiopalatina es una malformación congénita producida por la falta de fusión entre las estructuras que dan origen al labio superior y al paladar óseo secundario.1-3 Aproximadamente a las cuatro semanas en mamíferos, la cara está formada por el proceso frontal rodeado por los primeros y segundos arcos branquiales. Posteriormente, el proceso frontal da origen a los procesos frontonasales mediales y laterales, y el primer arco branquial origina los procesos maxilares y mandibulares. Los procesos nasomediales crecen más que los laterales, fusionándose a los procesos maxilares para formar el labio superior y el paladar primario.4 En la par te interna de la boca primitiva, las láminas palatinas del proceso maxilar se elevan y se fusionan en la línea media con el septum nasal para formar el paladar óseo secundario.5, 6 Fallas en la fusión de alguna de estas estructuras embrionarias generan fisuras.7, 8 Esta patología es clasificada en fisura labiopalatina sindrómica7 y no sindrómica (NSCLP, OMIM 119530).
La NSCLP presenta características de una enfermedad de herencia compleja, generado por factores ambientales y genéticos, los cuales producen variabilidad en la expresión fenotípica.9 La NSCLP es la más frecuente en el mundo. La etnia amerindia y asiática son las que presentan mayores tasas NSCLP.10, 11 En Chile, la NSCLP presenta tasa promedio de 1,5 por 1.000 recién nacidos vivos (RNV), afectando más a los hombres.12 La población chilena de estrato socioeconómico bajo presenta mayor composición de etnia amerindia y altas tasas de NSCLP. 13, 14 Esta anomalía se convier te en Chile en un problema de salud pública por su impacto social y económico, al disminuir los costos en su atención y rehabilitación.
Durante el desarrollo del labio superior y el paladar están involucrados tres procesos: la migración de las células de la cresta neural (del inglés Cranial Neural Crest CNC)15, 16 la transformación epitelio-mesénquima (del inglés: Epithelial-Mesenchymal transformation)17 y la formación de las estructuras medias craneofaciales.18-20 Estos procesos están regulados por la expresión de factores de transcripción y moléculas de secreción o de superficie celular,2, 9, 17, 21-25 Sonic hedgedhog (Shh), es una vía de señalización que probablemente se encuentra relacionada con la formación del labio superior y el paladar.9, 21, 23, 24, 26, 27
Evidencias en peces19, 27 y ratones,28 demuestran que alteraciones o deficiencias en Shh, interfieren en la migración de las CNC generando fenotipos similares a la NSCLP en humanos. En ratones se observó que Shh está relacionado con la transformación epitelio-mesénquima, generando fisuras palatinas cuando Shh es mutado.23 En pollos5, 29 y peces30 alteraciones de Shh generan estructuras mediales craneofaciales deficientes, fenotipos compatibles con el síndrome de la holoprosencefalia,31, 32 acompañado de fisura labiopalatina.
El gen Shh se ubica, en humanos, sobre la región del cromosoma 7q-36,3, está constituido por tres exones y dos intrones y tiene longitud de 29,4 kb*. Molecularmente, la proteína Sonic hedgehog (SHH),33 se comporta como una molécula de señalización intercelular, la cual se sintetiza por un precursor que sufre clivaje autocatalítico34 y el colesterol y ácido palmítico se unen covalentemente a SHH.34-38 Se cree que la adición de lípidos, colesterol y ácido palmítico restringe la movilidad de SHH en el ambiente extracelular.39, 40 Sonic hedgehog, es esencial para el desarrollo normal de muchos órganos y es un factor causal de la holoprosencefalia.34, 41 Shh se expresa en el epitelio de los arcos branquiales,42, 43 cuando se presentan alteraciones de Shh en el ratón conducen a fenotipos similares a la NSCLP.8, 23
Estos antecedentes conducen a la hipótesis que Shh estaría asociado al fenotipo NSCLP en humanos. Para contrastar esta hipótesis se utilizó una muestra de tríos caso-progenitores para identificar alelos, genotipos y haplotipos que fueron transmitidos de los progenitores a los descendientes y así determinar la asociación de Shh y NSCLP
MATERIALES Y MÉTODOS
Muestra de pacientes. Luego de la firma voluntaria del consentimiento informado por todos los individuos del estudio, el cual fue previamente aprobado por el Comité de ética de la Facultad de Medicina de la Universidad de Chile, de muestras de sangre periférica se extrajo el ADN total a 150 pacientes masculinos que presentaban la NSCLP y sus respectivos progenitores utilizando el protocolo modificado descrito por Maniatis.44 La NSCLP fue diagnosticada por un médico genetista.
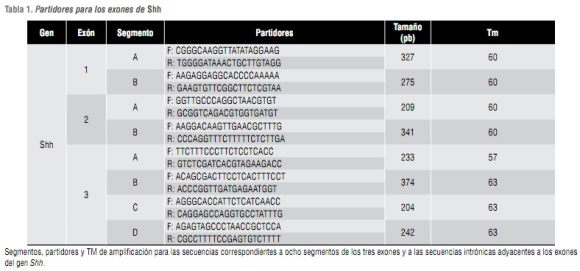
Se diseñaron cebadores o par tidores para amplificar ocho segmentos entre 200 y 400 pb (tabla 1) de las regiones exónicas e intrónicas adyacentes a los exones de Shh. Se amplificaron estos segmentos utilizando PCR. Para el diseño de los par tidores se utilizó el programa Primer 3.
Las muestras amplificadas se desnaturalizaron a 94 °C y renaturalizaron a 64, las posibles mutaciones presentes en la renaturalización conforman apareamientos de bases no complementarias, generando una deformación en la conformación de los amplificados los cuales son detectados en geles sensibles a estas deformaciones, geles MDE-CSG.
En el caso de encontrarse heterodúplex se procedió a la secuenciación del amplificado respectivo.
Análisis de SNP. El análisis de SNP fue alternativo al análisis de heterodúplex. Se analizaron dos SNP y para su selección se utilizó la base de datos NCBI y Proyecto Internacional HapMap, teniendo en cuenta las frecuencias de estos SNP en la población asiática y europea, dada la composición étnica de la población chilena. Se utilizó el programa DNA for Windows, para hallar las enzimas de restricción de los dos SNP. Además, se diseñaron los cuatro par tidores para amplificar estos dos SNP. Para determinar la presencia de los SNP en las muestras de los tríos caso-progenitores, se utilizaron las endonucleasas de restricción NlaIII para rs1233 555 y Sau3A1 para rs 1233556 (tabla 2) .

Análisis estadístico. Se hizo un análisis de alelos, genotipos y haplotipos para determinar la asociación de Shh y la FLPNS utilizando los test de desequilibrio de ligamiento (del inglés transmission-disequilibrium Test TDT), odd ratio y prueba de razón de verosimilitud. Para estos análisis se utilizó el programa UNPHASED.
RESULTADOS
Para determinar la asociación entre Shh y FLPNS se hizo un análisis de tríos caso-progenitores para determinar los alelos, haplotipos y genotipos transmitidos de padres a hijos mediante la técnica de heterodúplex. En la muestra de 150 tríos caso-progenitores, el análisis de los geles MDE-CSG no mostraron bandas retardadas en su migración.
La ausencia de bandas retardadas es sinónimo de ausencia de heterodúplex, lo cual indica que no se detectaron apareamientos erróneos de bases (mismatches) para los diferentes segmentos amplificados de las tres regiones exónicas e intrónicas adyacentes de Shh analizadas (figura 1) .
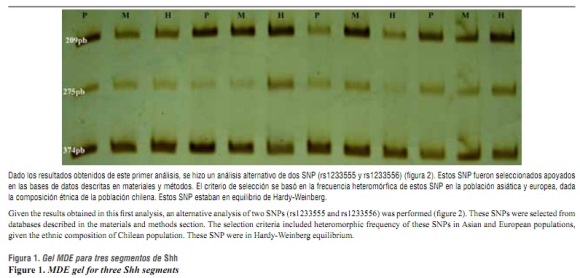
Estos dos SNP se encuentran en el intrón 1 (el primer recuadro de color fucsia de izquierda a derecha en la figura 2 de Shh, fueron seleccionados por su mayor frecuencia en la población asiática y europea utilizando las bases de datos de la National Center for Biotechnology Information (NCBI) y Proyecto Internacional HapMap. Shh: Sonic hedgedhog.

Los alelos de los SNP rs1233555 y rs1233556 fueron C > T y C > T, respectivamente. En las figuras 3A y 3B, se observan los resultados de la endonucleasa de restricción NlaIII para nueve amplificados del SNP rs1233555 y Sau3A1 para ocho amplificados del SNP rs 1233556.
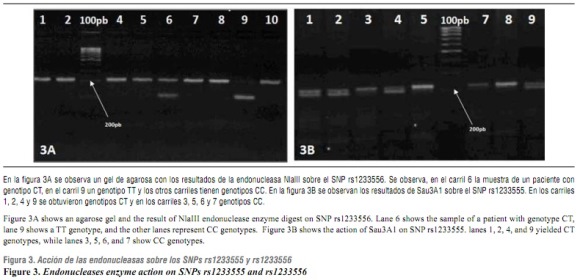
En el gel de agarosa, de la figura 3A, se observa el resultado de Nla III sobre amplificados del SNP rs1233556, en donde se obtuvieron los genotipos CT, TT y CC. En el gel de agarosa, de la figura 3B , se observa el resultado de Sau3A1 sobre amplificados del SNP rs1233555, obteniéndose los genotipos CT y CC. La combinación de los genotipos de los dos SNP conformó los haplotipos CC, CT, TC y TT. Estos haplotipos no mostraron resultados significativos de asociación con la NSCLP (tabla 3).
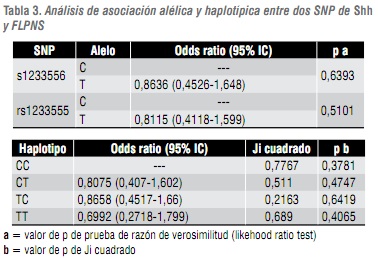
El análisis estadístico de alelos, genotipos y haplotipos no mostraron una transmisión preferencial de los mismos desde los progenitores a la progenie afectada. Estos resultados indican que no se detectó desequilibrio de ligamiento para los dos SNP analizados y por lo tanto no se detectó asociación entre la NSCLP y los marcadores utilizados (tabla 3).
DISCUSIÓN
El propósito de este trabajo fue demostrar que Shh está asociado a NSCLP. Al inicio se utilizaron geles MDE-SGS en donde no se observaron bandas retardadas en la migración del ADN de ocho segmentos de Shh de 150 tríos. Lo que condujo a un análisis de dos SNP en el primer intrón de este gen, el cual presentó evidencias experimentales en el desarrollo del labio superior y paladar. El análisis de SNP no mostró ninguna asociación entre Shh y NSCLP.
La no asociación entre el gen Shh y la NSCLP entre alelos, genotipo y haplotipo pudo deberse a que las regiones analizadas no están involucradas en esta asociación, pero es posible que las regiones regulatorias corriente arriba o abajo no analizadas tengan alguna injerencia directa o indirecta en la asociación de Shh y NSCLP. Esta injerencia fue sugerida por las evidencias en estudios experimentales que han demostrado que Shh está involucrado en los eventos de la migración de las células de la cresta neural craneales (CNCC –del inglés Cranial Neural Crest Cells), en la formación de las estructuras medial craneales y en la transformación epitelio-mesénquima. Estos eventos intervienen en la diferenciación y formación de las estructuras embrionarias que dan origen al labio superior y paladar.
Mutaciones y haploinsuficiencias de este gen en peces, pollos y ratas alteran la migración de las células de la cresta neural craneal o la inducción posicional de las células ectomesenquimales craneales, lo cual afecta los mecanismos de inducción, iniciación y ejecución de los programas específicos de diferenciación entre las CNCC, generando fenotipos similares a NSCLP en humanos.16, 23, 27 Pero no solo mutaciones de Shh afectan estos eventos, también estarían modulados por reguladores transcripcionales que activan varios genes, los cuales interaccionan con Shh.
Por ejemplo, en una etapa posterior a la migración e inducción de las CNCC, la interacción de Shh con el factor de crecimiento fibroblástico 10 (FGF10) afecta la transformación epitelio-mesénquima, proceso clave en la producción de la fisura labiopalatina.7 Otro ejemplo es el caso de la proteína morfogenética ósea 4 (BMP4), que induce la expresión de Shh a nivel del epitelio del borde medial de las láminas palatinas, permitiendo su crecimiento y fusión.21 Por lo tanto, mutaciones o deficiencias funcionales de estos genes que interactúan con Shh, podrían alterar dichas relaciones y producir el fenotipo NSCLP, sin que esté afectado Shh. También es probable que modificaciones leves de Shh influyan drásticamente en la regulación de otros genes que intervienen en la etiología de la NSCLP. Estas regulaciones por otros genes sobre Shh o viceversa, insinúan una relación epistática entre esos genes en la etiología de NSCLP.45, 46 También, otro aspecto impor tante de considerar, es la presencia de este gen en el síndrome de la holoprosencefalia en humanos, interpretándose que Shh podría ser clave en el desarrollo craneofacial dependiendo del tiempo y ambiente embrionario. Mutaciones de Shh severas o en estadios embrionarios tempranos podrían ser conducidas hacia síndromes acompañados de fisuras labiopalatinas, mientras que mutaciones o interacciones débiles con otros genes en estadios embrionarios tardíos, se derivarían en solo NSCLP. Podríamos especular entonces que las mutaciones o haploinsuficiencia en Shh no es un requisito único para producir NSCLP. Sin embargo, a diferencia de este estudio, Orioli y colaboradores encontraron mutaciones y polimorfismos de Shh relacionados con NSCLP.47 Por otra par te, es posible que el tamaño muestral y la rigurosidad en el control de algunos factores inherentes a la población chilena produzca las diferencias en los resultados de asociación de Shh y NSCLP entre las dos poblaciones utilizadas en los dos estudios.
Por otro lado, el número de SNP utilizados pudo ser insuficiente, o faltó incluir otros ubicados en regiones regulatorias de Shh para determinar la asociación de este gen con NSCLP, o que la distribución de estos SNP en la población chilena presentan una frecuencia diferente a las poblaciones referidas en este estudio, lo que no permitió detectar fehacientemente un posible rol de Shh en la NSCLP.
No obstante, se considera necesario analizar las regiones reguladoras cis y promotora de Shh, donde también podrían ubicarse variantes que influyan en el fenotipo de la NSCLP.
Podemos concluir que la falta de asociación puede deberse a que la frecuencia de distribución de los SNP en la población chilena se diferencia de las poblaciones refereridas, o que el número de SNP analizados fue insuficiente, o que faltó incluir las regiones no analizadas de Shh.
REFERENCIAS
1. Murray JC. Gene/environment causes of cleft lip and/or palate. Clin Genet 2002; 61(4): 248-256. [ Links ]
2. Helms JA, Cordero D, Tapadia MD. New insights into craniofacial morphogenesis. Development 2005; 132: 851-861. [ Links ]
3. Chai Y, Maxson RE. Recent advances in craniofacial morphogenesis. Dev Dyn 2006; 235: 2353-2375. [ Links ]
4. Radlanski RJ, Renz H. Genes, forces, and forms: mechanical aspects of prenatal craniofacial development. Dev Dyn 2006; 235(5): 1219-1229. [ Links ]
5. Hu D, Marcucio RS, Helms JA. A zone of frontonasal ectoderm regulates patterning and growth in the face. Development 2003; 130(9): 1749-1758. [ Links ]
6. Vasiri Sani F, Hallberg K, Harfe BD, McMahonc AP, Lindea A, Gritli-Linde A. Fate-mapping of the epithelial seam during palatal fusion rules out epithelial-mesenchymal transformation. Dev Biol 2005; 285(2): 490-495. [ Links ]
7. Murray JC, Schutte BC. Cleft palate: players, pathways, and pursuits. J Clin Invest 2004; 113(12): 1676-1678. [ Links ]
8. Rice R, Spencer-Dene B, Connor EC, Gritli-Linde A, McMahon AP, Dickson C et al. Disruption of Fgf10/Fgfr2bcoordinated epithelial-mesenchymal interactions causes cleft palate. J Clin Invest 2004; 113(12): 1692-1700. [ Links ]
9. Gritli-Linde A. Molecular control of secondary palate development. Dev Biol 2007; 301(2): 309-326. [ Links ]
10. Tolarová MM, Cervenka J. Classification and birth prevalence of orofacial clefts. Am J Med Genet 1998; 75(2): 126-137. [ Links ]
11. Vanderas AP. Incidence of cleft lip, cleft palate, and cleft lip and palate among races: a review. Cleft Palate J 1987; 24(3): 216-225. [ Links ]
12. Nazer J, Aravena T, Cifuentes L. Malformaciones congénitas en Chile: un problema emergente (periodo 1995-1999). Rev Med Chile 2001; 129(8): 895-904. [ Links ]
13. Palomino HM, Palomino H, Cauvi D, Barton SA, Chakraborty R. Facial clefting and Amerindian admixture in populations of Santiago, Chile. Am J Hum Biol 1997; 9(2): 225-232(a). [ Links ]
14. Palomino H, Cerda-Flores RM, Blanco R, Palomino HM, Barton SA, De Andrade M et al. Complex segregation analysis of facial clefting in Chile. J Craniofac Genet Dev Biol 1997; 17(2): 57-64(b). [ Links ]
15. Santagati F, Rijli FM. Cranial neural crest and the building of the vertebrate head. Nat Rev Neurosci 2003; 4(10): 806-818. [ Links ]
16. Jeong J, Mao J, Tenzen T, Kottmann AH, McMahon AP. Hedgehog signaling in the neural crest cells regulates the patterning and growth of facial primordial. Genes Dev 2004; 18(8): 937-951. [ Links ]
17. Kang P, Svoboda KKH. Epithelial-Mesenchymal transformation during craniofacial development. J Dent Res 2005; 84(8): 678-690. [ Links ]
18. Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of embryonic development of the zebrafish. Dev Dyn 1995; 203: 253-310. [ Links ]
19. Brand M, Heisenberg C-P, Warga RM, Pelegri F, Karlstrom RO, Beuchle D et al. Mutations affecting development of the midline and general body shape during zebrafish embryogenesis. Development 1996; 123: 129-142. [ Links ]
20. Schilling TF, Kimmel CB. Musculoskeletal patterning in the pharyngeal segments of the zebrafish embryo. Development 1997; 124(15): 2945-2960. [ Links ]
21. Zhang Z, Song Y, Zhao X, Zhang X, Fermin C, Chen Y. Rescue of cleft palate in Msx1-deficient mice by transgenic Bmp4 reveals a network of BMP and SHH signaling in the regulation of mammalian palatogenesis. Development 2002; 129(17): 4135-4146. [ Links ]
22. Zucchero TM, Cooper ME, Maher BS, Daack-Hirsch S, Nepomuceno B, Ribeiro L et al. Interferon regulatory factor 6 (IRF6) gene variants and the risk of isolated cleft lip or palate. N Engl J Med 2004; 351(8): 769-780. [ Links ]
23. Rice R, Connor E, Rice DPC. Expression patterns of hedgehog signalling pathway members during mouse palate development. Gene Expr Patterns 2006; 6(2): 206-212. [ Links ]
24. Haworth KE, Wilson JM, Grevellec A, Cobourne MT, Healy C, Helms JA et al. Sonic hedgehog in the pharyngeal endoderm controls arch pattern via regulation of Fgf8 in head ectoderm. Dev Biol 2007; 303: 244-258. [ Links ]
25. Zhang W, Kang JS, Cole F, Yi MJ, Krauss RS. Cdo Functions at multiple points short article in the Sonic hedgehog pathway, and cdo-deficient mice accurately model human holoprosencephaly. Dev Cell 2006; 10(5): 657-665. [ Links ]
26. Helms JA, Kim CH, Hu D, Minkoff R, Thaller C, Eichele G et al. Sonic hedgehog participates in craniofacial morphogenesis and is down-regulated by teratogenic doses of retinoic acid. Dev Biol 1997; 187: 25-35. [ Links ]
27. Wada N, Javidan Y, Nelson S, Carney TJ, Kelsh RN, Schilling TF. Hedgehog signaling is required for cranial neural crest morphogenesis and chondrogenesis at the midline in the zebrafish skull. Development 2005; 1 32: 3977-3988. [ Links ]
28. Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H et al. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 1996; 383: 407-413. [ Links ]
29. MacDonald ME, Abbott UK, Richman JM. Upper beak truncation in chicken embryos with the cleft primary palate mutation is due to an epithelial defect in the frontonasal mass. Dev Dyn 2004; 230(2): 335-349. [ Links ]
30. Takamiya M, Campos-Ortega JA. Hedgehog signalling controls zebrafish neural keel morphogenesis via its leveldependent effects on neurogenesis. Dev Dyn 2006; 235(4): 978-997. [ Links ]
31. Cordero D, Marcucio R, Hu D, Gaffield W, Tapadia M, Helms JA. Temporal perturbations in Sonic hedgehog signaling elicit the spectrum of holoprosencephaly phenotypes. J Clin Invest 2004; 114(4): 485-494. [ Links ]
32. Maity T, Fuse N, Beachy PA. Molecular mechanisms of Sonic hedgehog mutant effects in holoprosencephaly. Proc Natl Acad Sci USA 2005; 102(47): 17026-17031. [ Links ]
33. Varjosalo M, Taipale J. Hedgehog signaling. J Cell Sci 2007; 120: 3-6. [ Links ]
34. Nanni L, Ming JE, Bocian M, Steinhaus K, Bianchi DW, Die-Smulders C et al. The mutational spectrum of the Sonic hedgehog gene in holoprocencephaly: Shh mutations cause a significant proportion of autosomal dominant holoprocencephaly. Hum Mol Genet 1999; 8(13): 2479-2488. [ Links ]
35. Chamoun Z, Mann RK, Nellen D, Von Kessler DP, Bellotto M, Beachy PA et al. Skinny hedgehog, an acyltransferase required for palmitoylation and activity of the hedgehog signal. Science 2001; 293: 2080-2084. [ Links ]
36. Lee JD, Treisman JE. Sightless has homology to transmembrane acyltransferases and is required to generate active hedgehog protein. Curr Biol 2001; 11(14): 1147-1152. [ Links ]
37. Amanai K, Jiang J. Distinct roles of central missing and dispatched in sending the hedgehog signal. Development 2001; 128: 5119-5127. [ Links ]
38. Micchelli CA, The I, Selva E, Mogila V, Perrimon N. Rasp, a putative transmembrane acyltransferase, is required for Hedgehog signaling. Development 2002; 129(4): 843-851. [ Links ]
39. Gallet A, Rodríguez R, Ruel L, Therond PP. Cholesterol modification of hedgehog is required for trafficking and movement, revealing an asymmetric cellular response to hedgehog. Dev Cell 2003; 4: 191-204. [ Links ]
40. Chen MH, Lin Y-J, Kawakami T, Xu SM, Chuang PT. 2004. Palmitoylation is required for the production of a soluble multimeric Hedgehog protein complex and long-range signaling in vertebrates. Genes Dev 2004; 18: 641-659. [ Links ]
41. Nieuwenhuis E, Hui CC. Hedgehog signaling and congenital Malformations. Clin Genet 2005; 67(3): 193-208. [ Links ]
42. Moore-Scott BA, Manley NR. Differential expression of Sonic hedgehog along the anterior-posterior axis regulates patterning of pharyngeal pouch endoderm and pharyngeal endoderm-derived organs. Dev Biol 2005; 278(2): 323-335. [ Links ]
43. Yamagishi C, Yamagishi H, Maeda J, Tsuchihashi T, Ivey K, Hu T et al. Sonic hedgehog is essential for first pharyngeal arch development. Pediatr Res 2006; 59(3): 349-354. [ Links ]
44. Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. A laboratory manual. New York: Cold Spring Harbor Laboratory Press; 1989. [ Links ]
45. Ferguson MWJ. Craniofacial malformations: towards a molecular understanding. Nature Genet 1994; 6: 329-330. [ Links ]
46. Ito Y, Yeo JY, Chytil A, Han J, Bringas P Jr, Nakajima A et al. Conditional inactivation of Tgfbr2 in cranial neural crest causes cleft palate and calvaria defects. Development 2003; 130: 5269-5280. [ Links ]
47. Orioli IM, Vieira AR, Castilla EE, Ming JE, Muenke M. Mutacional analysis of the Sonic hedgehog in 220 newborns with oral clefts in a south American (ECLAMC) population. Am J Med Genet 2002; 108(1): 12-15. [ Links ]
CORRESPONDENCIA
Ramiro J. Rincón R.
Facultad de Odontología
Universidad de Antioquia
Correo electrónico: ramirojrr@gmail.com
