










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkORINOQUIA
On-line version ISSN 0121-3709
Orinoquia vol.18 supl.1 Meta Dec. 2014
ARTÍCULO ORIGINAL/ORIGINAL ARTICLE
Desempeño productivo del coporo con dietas isotópicas
Productive performance of coporo with isotopic diets
Desempenho produtivo do coporo com rações isotópicas
Johana P. Ortega1*
Martha I. Yossa2*
1Zootecnista, MSc, Dr
2MVZ, Esp. MSc, Dr
* Grupo de investigación en alimentación y nutrición de organismos acuáticos-GRANAC, Instituto de Acuicultura de los Llanos. Universidad de los Llanos, km 12 vía Puerto López, Villavicencio, Meta-Colombia Email: granac.iall@gmail.com
Recibido: octubre 30 de 2014. Aceptado: noviembre 28 de 2014
Resumen
Los peces detritívoros son la base de la cadena alimentaria y están ampliamente distribuidos en las cuencas neotropicales, los Prochilodus son representativos de este grupo y el bocachico llanero o coporo (Prochilodus mariae) es de interés comercial en acuicultura. Para evaluar el desempeño productivo en alevinos de coporo con dietas isotópicas, fueron conformados dos tratamientos: dieta comercial (δ13C -20.93±0.34) y dieta experimental (δ13C -24.6±0.9) suministradas durante sesenta días. Parámetros de peso medio, longitud media estándar, ganancia de peso (GP), tasa de crecimiento específico (TCE) y factor de conversión alimenticia (FCA) fueron determinados. La contribución y fraccionamiento de la dieta fueron estimados a partir de las lecturas isotópicas en muestras de músculo. El peso y longitud estándar del coporo fue superior en el tratamiento de la dieta comercial (DC). En los parámetros GP y TCE hubo diferencia significativa (P<0.05) entre tratamientos, siendo el tratamiento DC siempre superior frente a la dieta experimental (DE). La contribución relativa de la DC y DE fue de 59.2 y 53.4% respectivamente, en cuanto que el mayor fraccionamiento de la dieta en carbono y nitrógeno fue de la DE en 1.97 y 1.14%0 respectivamente. La relación entre la contribución de la dieta y la ganancia de peso fue más eficiente para DC en términos de carbono, contrariamente, la eficiencia del nitrógeno fue con DE. Aunque la contribución de las dietas fue inferior a 60%, ambas contribuyeron a mejorar el desempeño del coporo.
Palabras clave: Fraccionamiento, Isotopos estables, Prochilodus mariae, TCE.
Abstract
The detritivorous fish are the base of the food chain and are widely distributed in neotropical basins, Prochilodus are representative of this group and bocachico llanero or coporo (Prochilodus mariae) is of commercial interest in aquaculture. This study aimed to evaluate the productive performance in fingerlings coporo with isotopic diets were comprised two treatments: commercial diet (δ 13C -20.93±0.34) and experimental diet (δ 13C -24.6±0.9) supplied for sixty days. Mean weight, mean standard length, weight gain (WG), specific growth rate (SGR) and feed conversion ratio (FCR) were determined. The contribution and fractionation of the diet were estimated from isotopic values muscle samples. The weight and length standard of coporo was more than in the treatment of the commercial diet (CD). In the WG and SGR parameters was significant difference (P< 0.05) between treatments, with the CD treatment better compared to the experimental diet (ED). The relative contribution of CD and DE were 59.2 and 53.4% respectively, in isotopic fractionation of carbon and nitrogen diet of ED was 1.97 and 1.14%0 respectively. The relation between the contribution of diet and weight gain was most efficient CD in terms of carbon contrary, nitrogen efficiency was with ED. Although the contribution of the diets was less than 60%, both contributed to improving the performance of coporo.
Key Words: Isotopic fractionation, Prochilodus mariae, SGR, Stables isotopes.
Resumo
Os peixes detritívoros são a base da cadeia alimentar e são amplamente distribuídos nas bacias neotropicais, os Prochilodus são representativos deste grupo e o bocachico llanero ou coporo (Prochilodus mariae) é de interesse comercial na aquicultura. A fim de avaliar o desempenho produtivo nos alevinos de coporo com dietas isotópicas, foram constituídos dois tratamentos: racão comercial (δ13C -20.93±0.34) e racão experimental (δ13C -24.6±0.9) fornecidas durante 60 dias. parâmetros médios, a média de comprimento padrão, ganho de peso (GP), taxa de crescimento específico (TCE) e conversão alimentar (CA) foram determinados. A contribuição do fracionamento da dieta foram estimados com as leituras de isótopos em amostras de músculo. O peso e comprimento padrão do coporo foi maior no tratamento da dieta comercial (DC). Nos parâmetros GP e TCE houve diferença significativa (P<0,05) entre os tratamentos, o tratamento DC sempre foi maior que o dieta experimental (DE). A contribuição relativa de DC e DE foram na ordem de 59.2 e 53.4%, o fracionamento da dieta no carbono e nitrogênio da ED foi de 1.97 e 1.14%0 respectivamente. A relação entre a contribuição da dieta e ganho de peso foi mais eficiente em termos de carbono na DC mas a eficiência de nitrogênio foi maior na DE. Embora a contribuição das dietas foi menor do 60%, as duas contribuíram para melhorar o desempenho do coporo.
Palavras chave: Isotopos estaveis, Fracionamento, Prochilodus mariae, TCE.
Introducción
Los peces detritívoros son la base de la cadena alimentaria y están ampliamente distribuidos en las cuencas neotropicales (Barthem y Fabre, 2003). Los Prochilodus son representativos de este grupo y, durante la última década, en Colombia la pesca comercial ha reportado grandes volúmenes que llegan a representar el 30, 54 y 31% en las cuencas del Amazonas, Magdalena y Orinoco respectivamente. En la cuenca del Orinoco se ha presentado un incremento del 47% en los desembarques de P. mariae, comúnmente conocido como bocachico llanero o coporo, siendo la segunda especie de mayor importancia con un aporte de 153.4 toneladas (MADR y CCI, 2010).
Los Prochilodus, poseen un cuerpo fusiforme, alargado y comprimido lateralmente, boca protráctil que le permite succionar el detrito y realizar raspado a sustratos (Bowen, 1984). Ésta estructura sumada a las característica fisiológicas del sistema digestivo integrado por un estómago dividido en dos partes (cardíaco y pilórico): el primero, sirve de reserva y está cubierto por un epitelio superficial y glandular, que lo protege de la acción mecánica y secreciones de ácido clorhídrico y pepsinógeno; el segundo, es rígido, en él y en el intestino se realiza la digestión con ayuda de la acción enzimática que permite reducir las partículas ingeridas. El esófago tiene la función de predigestión mecánica y química del alimento (Bowen, 1984; Chaves y Vazzoler, 1984).
Esta compleja estructura le permite el aprovechamiento del detrito formado a partir de microalgas, zooplancton y materia orgánica en descomposición, con bajos niveles de proteína y energía (Yossa y AraújoLima, 1998; Yossa, 2002). Estos hábitos podrían estar relacionados con el bajo desempeño productivo de los Prochilodus en cultivo (Urbano et al., 2009).
Para mejorar el desempeño zootécnico de los Prochilodus en cultivo, ha habido diversos experimentos, utilizando dietas purificadas, semipurificadas, convencionales y con abonos orgánicos, con alto contenido de proteína y energía bruta, isotópicas y otros con alimentos simulando ambiente natural (Caicedo et al., 2002; González y Wills 2003; González y Wills 2005; González y Wills, 2009; Yossa y Vásquez-Torres,2012; Sacramento, 2014). El interés en el cultivo de estas especies radica, entre otros aspectos, en el valor agregado que representa como limpiador del estanque, además de ser una especie de alto consumo nacional y optima especie para policultivos, sistema en el puede alcanzar 500 gramos en 12 meses aproximadamente (MADR y CCI, 2010).
Ya en ambiente natural las investigaciones se han enfocado en la identificación de la fuente de carbono y explicar la asimilación del detrito utilizando los análisis isotópicos, como una herramientas eficaz, aplicada a la nutrición de peces (Fosrberg 1993; Faria y Benedito, 2011).
Los análisis isotópicos se han utilizado en estudios ambientales y ecológicos, para establecer la posición trófica, la incorporación y asimilación de nutrientes para el crecimiento de especies marinas y dulceacuícolas. Por ejemplo, Lopes et al., (2007), determinaron a través de isotopos de carbono que la dieta asimilada por Prochilodus lineatus en ambientes naturales provenía de fuentes de carbono C3. Y con el isótopo de nitrógeno, el cual actúa como indicador del nivel trófico de las especies, se ha demostrado la influencia de la edad de los organismos en la variación isotópica (Overman y Parrish, 2001). A partir de las lecturas isotópicas de C y N, es posible establecer el fraccionamiento de la dieta (Van-Zander y Rasmussen, 2001) que es la diferencia entre el depredador y su presa o fuente (dieta) y son los carnívoros los que presentan mayores valores en el fraccionamiento.
Las plantas C3 y C4, difieren en las rutas metabólicas que usan en la fijación del carbono durante el proceso de fotosíntesis, el producto derivado de este proceso tiene valores isotópicos de carbono 13 (δ13C) que son expresados en partes por mil (%0). Las plantas C3 presentan valores isotópicos entre -20 y -35 %0; las C4 poseen valores de -9 a -17%0 (Santiago et al., 2005), en estos términos las plantas C4 son más enriquecidas que las C3.
El grupo de plantas C3 está integrado entre otras por el fitoplancton, cianobacterias, arroz, soya, papa, trigo, centeno, cebada, hortalizas; mientras que en el grupo C4 se encuentran las plantas vasculares como el maíz, caña de azúcar, sorgo, mijo y perla.
En ambiente natural, la dieta que soporta la abundancia de los Prochilodus es con base en detrito con fuente de carbono C3 (Araujo-Lima et al.,1986; Benedito et al., 2000) la estructura digestiva de esta especie, garantiza el aprovechamiento del detrito y otros ítems alimentarios de bajo valor proteico, lo que podría a su vez, ser el impedimento para el aprovechamiento de alimentos enriquecidos, como los suministrados en los sistemas de producción piscícola. Estos inconvenientes incentivan la búsqueda de alternativas nutricionales para mejorar su desempeño. Así, el objetivo de este trabajo fue evaluar el desempeño zootécnico en alevinos de coporo con dietas isotópicas.
Metodologia
Localización
El experimento se desarrolló en el Instituto de acuicultura de la Universidad de los Llanos-IALL (4°09´09"N y 73°46´22 W), en Villavicencio- Meta, a una altura de 418 msnm, temperatura promedio de 26 °C, humedad relativa del 75% y precipitación pluvial de 4.050 mm.
Dietas
Se utilizaron dos dietas, una comercial (DC) que venía siendo suministrada a los alevinos y otra experimental (DE), ambas con contenido proteico de 24%, como usado por González y Wills (2009) en la especie Prochilodus magdalenae. La formulación de la DE (Tabla 1) fue con base en materias primas de carbono tres (C3) utilizadas convencionalmente en los alimentos concentrados para peces, y elaborada según los protocolos del Laboratorio Experimental de Alimentación y Nutrición de Peces (LEANP) del IALL. Las materias primas fueron molidas finamente en micromolino (Scienceware, USA), pesadas y homogenizadas en seco, mezcladas con agua, aceite, alfacelulosa, vitaminas y minerales y sometidas a proceso de extrusión a 123 °C (micro-extrusora Exteec®, Riberão Preto -Brasil) para formar gránulos compactos, flotantes de 0.3 mm diámetro. La dieta fue secada en horno a 60 °C, manteniendo peso constante y almacenada en bolsas de cierre hermético a 4 °C hasta su uso. Para determinar la calidad de las dietas (DE y DC) fue realizado un análisis proximal (Tabla 1) de acuerdo con los métodos de la AOAC (2005) para determinación de: materia seca, proteína y ceniza; para energía fue usada la bomba calorimétrica (Parr121 EA USA).
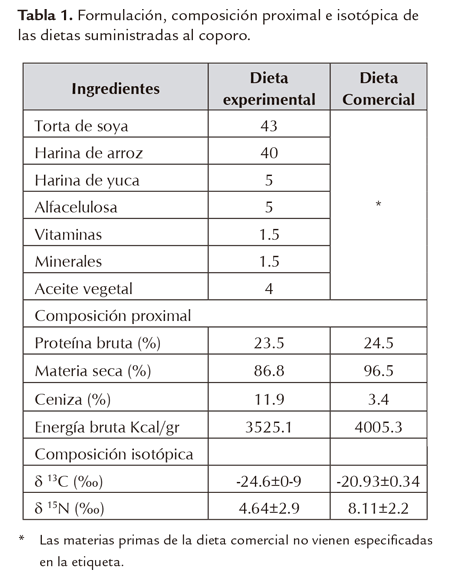
Material biológico
Alevinos (150) de coporo de 35 días de eclosionados y de peso promedio de 0.76±0.2g, donados por una estación piscícola, fueron trasladados en bolsas plásticas con agua y oxígeno en proporción 1:3, al laboratorio de bioensayos en el IALL. Un total 130 alevinos fueron distribuidos equitativamente en dos acuarios de 50l c/u, que contenían agua madurada durante una semana, aireación constante y termostatos que mantuvieron la temperatura en 25°C±0.02. Durante el periodo de aclimatación que fue de 8 días como recomendado por González y Wills (2003) se suministró hasta aparente saciedad la dieta (DC y DE) correspondiente a cada acuario. Los 20 alevinos restantes fueron sometidos a choque térmico, eviscerados y congelados para análisis posteriores.
Finalizado el periodo de aclimatación, los alevinos de cada acuario (DC y DE) fueron anestesiados con 75 mg/l de eugenol diluido en alcohol etílico al 92% (Vidal et al., 2008), para pesarlos en gramera electrónica (Metter Toledo AB204-S) con sensibilidad de ±0.1mg, medirlos y conformar por cada acuario, cuatro grupos de diez peces lo más homogéneo posible.
Unidades experimentales
Para iniciar el experimento cada grupo de peces fue trasladado a un acuario de 30l, con agua igualmente madurada durante una semana, con pH de 6.69±0.02 y temperatura de 26.3± 0.9 oC elevada y controlada con termostato para garantizar la zona de confort. Desde el inicio y hasta el final del experimento, las ocho unidades experimentales permanecieron en un área cerrada con condiciones controladas y monitoreo diario (T°C, pH, O2), con aireación y termostato para constante. Recambio de agua del 10% y sifoneo realizado todos los días para retirar el alimento no ingerido; las heces y desechos metabólicos, evitando así alteraciones en la calidad del agua que fue monitoreada con equipo multiparamétrico YSI Pofessional plus.
La ubicación de unidades experimentales fue completamente al azar. A cada grupo de peces en su unidad experimental se le suministró dieta a razón del 15% de la biomasa inicial, en dos raciones diarias (9:00 y 16:00 horas) de acuerdo con González y Wills (2005). Durante los sesenta días del experimento se realizaron ajustes al suministro de alimento de acuerdo a la mortalidad y a la observación de consumo de la dieta.
Parámetros zootécnicos
Para estimar los parámetros: peso medio, longitud media estándar, ganancia de peso (GP), tasa de crecimiento especifico (TCE) y factor de conversión alimenticia (FCA); se usaron las siguientes fórmulas:

Para determinar el peso final se siguió la rutina de manipulación descrita al iniciar el experimento.
Fraccionamiento de la dieta
Para estimar la contribución de cada dieta, fueron tomados los veinte alevinos reservados a priori a la fase de aclimatación y el total de cada unidad experimental al final del ensayo, para obtener muestras de músculo tipo filete. Las muestras de músculo (inicial y final) y de dietas (comercial y experimental) se secaron en estufa a 60°C por 72 horas, luego se maceraron finamente de cada unidad experimental se conformó un pool-, se empacaron (~6mg) en tubos eppendorf y fueron enviadas para análisis de isotopos a la Universidad de Georgia, EE.UU (Institute of Ecology Analytical Chemistry Laboratory). Los análisis de carbono y nitrógeno se realizaron por triplicado, sometiendo 1mg de muestra a combustión en un analizador elemental de gases (Thermo Finnigan Delta Plus) conectado a un espectrómetro de masa, los valores isotópicos son expresados con la notación δ en partes por mil (‰) relativos al estándar internacional Pee Dee Belemnite (PDB) para el δ13C y nitrógeno atmosférico para δ15N.
El fraccionamiento (Δ) fue determinado a partir de la ecuación de Michener y Schell (1994)
ΔC = δ13C músculo - δ13C dieta
ΔC = δ15N músculo - δ15N dieta
Diseño experimental
Se estableció un diseño experimental completamente al azar, con dos tratamientos: dieta comercial (DC) y dieta experimental (DE); con cuatro repeticiones cada uno y cada repetición con 10 alevinos.
Análisis estadístico
Los parámetros evaluados (peso medio, longitud estándar, ganancia de peso, tasa de crecimiento específica y tasa de conversión alimenticia) son presentados como la media ± desvío estándar y fueron transformados por el método arco-seno para realizar ANDEVA. En las tres últimas variables se realizó ANDEVA de tratamientos des-balanceados porque ocho días antes, una réplica fue perdida en su totalidad. Para los valores significativos (P<0.05) se realizaron comparaciones de medias utilizando el test de Tukey a un nivel de significancia del 5%. Para estimar la contribución del carbono de la dieta, se trabajaron las lecturas isotópicas de músculo final que evidenciaron cambios frente al músculo inicial y los resultados de las lecturas isotópicas se analizaron a través de gráficos del tipo box-plot. Para la relación entre la contribución de la dieta y ganancia de peso se hizo un análisis de regresión simple. Los datos se analizaron con el programa Systat for Windows (v13.0, SPSS Inc., USA). Para establecer el fraccionamiento de las dietas se tomaron las medias de las lecturas isotópicas de carbono y nitrógeno y se aplicó la respectiva fórmula.
Resultados Parámetros Zootécnicos
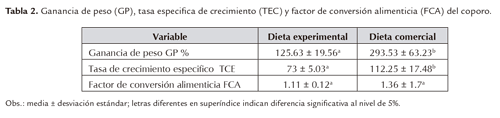
Al final del experimento en el tratamiento con dieta comercial (DC) se registró el mayor peso y longitud (3.54± 2.71 y 5.32±1.16) respectivamente, mientras que en el tratamiento con dieta experimental (DE) estas variables fueron inferiores (2.30±0.78 y 4.56±0.58). La ganancia de peso (GP), la tasa específica de crecimiento (TEC) y el factor de conversión alimenticia (FCA), son presentados en la tabla 2. En los parámetros GP y TEC hubo diferencia significativa (P<0.05) entre tratamientos, siendo el tratamiento DC siempre superior frente a DE.
Composición isotópica de la dieta y el músculo
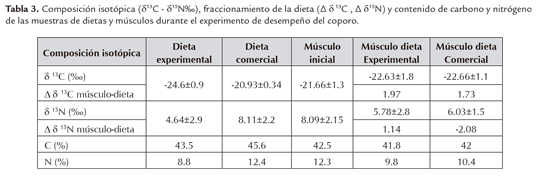
En la tabla 3, se registra la composición δ13C (‰), δ15N (‰) y los porcentajes totales de carbono y nitrógeno para las muestras de las dietas (DE, DC) y de músculo (inicial y final) en los tratamientos. Se denota que ambas dietas son similares isotópicamente ya que las lecturas δ13C están en el rango de las fuentes C3. Se evidencia también la memoria isotópica de la dieta comercial a través de las lecturas δ13C (‰) y δ15N (‰) en el músculo inicial (Tabla 3).
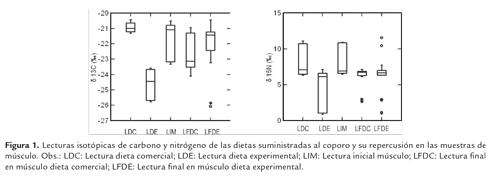
La comparación entre las lecturas de δ13C y δ15N de las muestras de músculo inicial y final frente a la dieta suministrada, se observa en figura 1. Entre tratamientos no hubo diferencias significativas (P>0.05), sin embargo, en el tratamiento DC se observó menor enriquecimiento δ13C referente al valor de la dieta y al músculo inicial. En el tratamiento DE el valor δ13C del músculo permaneció más próximo a su valor inicial. Los valores medios del δ15N permanecieron en un rango estrecho en todas las muestras analizadas.
Fraccionamiento de la dieta y relación entre la contribución de carbono y nitrógeno con la ganancia de peso del coporo
El fraccionamiento para ambas dietas se describe en la tabla 3. En la relación entre GP y contribución del carbono (Figura 2) el mejor ajuste en el tratamiento DC fue con el modelo polinomial. La ganancia de peso en relación a la incorporación del carbono total, tuvo una relación positiva R = 0.71, mientras que en el tratamiento DE el mejor ajuste fue con el modelo lineal y la relación aunque positiva fue menor R= 0.59.
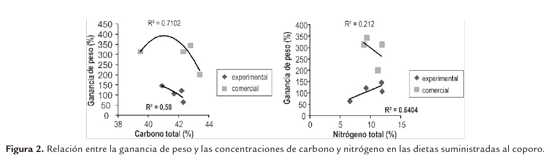
Para la relación entre nitrógeno total (%) y GP (Figura 2), con un modelo lineal se realizó el mejor ajuste. El tratamiento DC tuvo una relación débil estadísticamente R= 0.21 y se puede atribuir a la variación inter-especifica en la especie. El aporte de nitrógeno al tratamiento DE fue mejor aprovechado (R= 0.64). Fue así como a medida que aumentó la incorporación del N (%) en el músculo la ganancia de peso se incrementó. Demostrando una correlación directa entre la ganancia de peso y el aporte de N (%) total.
Discusión Parámetros zootécnicos
Considerando que los parámetros de producción están relacionados con el tipo de dieta suministrada, el estado fisiológico del pez, la calidad de agua, la edad, el sexo, el estrés (Martínez-Porchas et al., 2009) y factores externos, este estudio se realizó en un área cerrada, en condiciones controladas, con alevinos de un mismo desove y aclimatados bajo las mismas prácticas, a fin de minimizar posibles efectos sobre el desempeño de P. mariae en cada tratamiento.
Los Prochilodus en ambiente natural consumen detrito con contenido proteico del 20% (Yossa y Araújo-Lima, 1998; Yossa, 2002), en este estudio con el suministro de dietas con 24% de proteína, los alevinos de coporo tuvieron una ganancia de peso superior al 100% y de 70% en la tasa de crecimiento especifico (Tabla 2). Sin embargo para P. affins con una dieta de 26% de proteína Bomfim et al., (2005) reportó una GP y TCE de apenas 60 y 46% respectivamente, entretanto González y Wills (2003) para P. magdalenae conuna dieta de 28% de proteína obtuvieron una GP de 98%. A pesar de ser dietas diferentes, la respuesta de los Prochilodus a bajas concentraciones proteicas estaría relacionada con su ecología trófica. En este contexto, el coporo fue más eficiente con la dieta experimental (FCA=1.1, Tabla 2) que es empobrecida con respecto a la dieta comercial en términos de materia orgánica (Tabla 3).
Composición isotópica de la dieta y en el músculo
Las lecturas isotópicas de carbono en las dietas DE (-24.60‰) y DC (-20.93‰) fueron similares, esto permitió verificar que la fuente de carbono de la dieta comercial fue C3 (Figura 1 y 2; Tabla 3). El valor medio de δ13C en el músculo del tratamiento DE y DC fue de -22.63±1.8 y -22.66±1.1‰ respectivamente, estos cambios con relación a la lectura inicial del músculo (Tabla 3), reflejan la renovación del tejido de acuerdo a la dieta consumida. Estos valores fueron inferiores al reportado por Yossa y Vásquez-Torres (2012) para P mariae (-21.44±0.84‰) en cultivo que reflejó un incremento con relación a la dieta consumida (-20.44±0.69‰). Similar comportamiento se observó en DC con el incremento dieta-músculo inicial (0.73‰) y músculo inicial-músculo final (1‰) (Tabla 3), sin embargo en este caso la memoria isotópica juega un papel fundamental, si se considera que esta dieta fue la suministrada durante todo el desarrollo de los alevinos. En el caso de DE al final del experimento, aún no reflejaba la δ13C de la dieta consumida y para lograr el equilibrio con δ13C del músculo inicial se requiere un incremento del 3‰ (Tabla 3). En ambiente natural se ha reportado para una misma población de P. linneatus δ13C de -24.5 y -26.8‰ (Lopes et al., 2007), con lo que se puede inferir el proceso de renovación de tejido similar al del DE. Con estos reportes se corrobora que en ambiente natural y en cultivo la fuente de energía para los Prochilodus es C3.
En términos de δ15N se pudo apreciar que el valor inicial en el músculo reflejó la composición de la dieta comercial (Tabla 3) a la que venían acostumbrados los alevinos; al finalizar el experimento la composición isotópica en el músculo vario en ambos grupos, denotando una menor asimilación posiblemente por una redistribución de nutrientes (Abimorad et al., 2014) y en el caso de DE es posible que la proteína fuese de menor calidad.
La incorporación de carbono y nitrógeno en ambos tratamientos fue superior a 40 y 9% respectivamente (Tabla 3), valores que difieren del trabajo realizado por Abimorad et al., (2014) en juveniles de Piaractus mesopotamicus alimentados con dietas suplementadas con aminoácidos, donde la incorporación fue mayor a 90% para carbono y nitrógeno. El menor porcentaje de incorporación obtenido en este ensayo podría ser explicado por la característica morfológica del tracto digestivo de los detritívoros que conlleva a un metabolismo lento (Bowen, 1984).
Fraccionamiento de las dietas
Los valores de fraccionamiento isotópico del carbono (Δ13C) de -0.6 a 2.7‰ propuesto por DeNiro y Epstein (1978) y del nitrógeno (Δ15N ) de 1.3 a 5.3‰ según Minagawa y Wada (1984) son los valores usados como referencia en estudios de ecología alimentar. Los resultados obtenidos en este experimento para Δ13C y Δ15N (Tabla 3) se mostraron fuera de estos valores de referencia, igual a los reportados por Sacramento (2014) para P. lineatus y para carpa y tilapias por Focken y Becker (1998) quienes reportaron que estos valores se deben a la gran variabilidad intra e interespecifica asociada a factores ambientales, zootécnicos y fisiológicos.
Conclusiones
Ambas dietas contribuyeron a mejorar el desempeño zootécnico de los alevinos de coporo, sin embargo en términos de ganancia de peso, tasa especifica de crecimiento y factor de conversión alimenticia fue más diciente en el tratamiento de dieta comercial. En cuanto al fraccionamiento fue en el tratamiento de dieta experimental donde se registró mayor incorporación de la dieta en el músculo. Para lograr una mayor contribución de la dieta y por ende mejorar el desempeño del coporo, se recomienda establecer experimentos de mayor duración que permitan establecer el turnover isotópico o taxa de reposición en mayor proporción.
Agradecimientos
Al grupo GRANAC por el apoyo en este trabajo a través del proyecto "Fuentes de carbono" convocatoria IIOC 2010, al Instituto de acuicultura de los Llanos-IALL por el apoyo con la infraestructura, a la Universidad Nacional Abierta y a Distancia, y a la Piscícola Aquapeces por la donación de los alevinos utilizados en este experimento. Este trabajo forma parte de los resultados de la tesis de JP Ortega.
Referencias
AOAC-Association of Official Analytical Chemists. 2005. Official methods of Analysis. Washington. D.C., USA. [ Links ]
Abimorad EG, Ducatti C, Castellani D, Jomori RK, Portella, M.C. The use of stable isotopes to investigate the effects of supplemental lysine and methionine on protein turnover and amino acid utilization in pacu, Piaractus mesopotamicus, juveniles. Aquaculture. 2014;433:119-124. [ Links ]
Araujo-Lima CA, Forsberg BR, Victoria R, Martinelli L. Energy sources for detritivorous fishes in the Amazon. Science. 1986;234:1256-1258. [ Links ]
Barthem RB, Fabre NN. 2003. Biologia e diversidade dos recursos pesqueiros da amazônia. En: Ruffino (Editores). A pesca e os recursos pesqueiros na Amazônia Brasileira. Manaus. [ Links ]
Benedito-Cecilio E, Araujo-Lima CAR, Forsberg BR, Bittencourt MM, Martinelli LC. Carbon sources of Amazonian fisheries. Fish Manag Ecol. 2000;7:305-315. [ Links ]
Bomfim MD, Lanna ET, Serafini MA, Ribeiros FB, Da Silva PK. Proteína Bruta e Energia Digestível em Dietas para Alevinos de Curimbatá (Prochilodus affins). Rev Bras Zootec. 2005;34:1795-1806. [ Links ]
Bowen SH, Bonetto AA, Ahlgren MO. Microorganisms and detritus in the diet of a typical neotropical riverine detritivore Prochilodus platensis (Pisces, Prochilodontidae). Limnol Oceanogr. 1984;29:1120-1122. [ Links ]
Caicedo J, Pérez A, Wills A. 2002. Efecto de dos presentaciones de alimento durante la fase de levante en el desarrollo del bocachico Prochilodus magdalenae (Steindachner 1878). I Congreso Internacional virtual de acuicultura CIVA. [24/03/2015] URL: http://www.revistaaquatic.com. [ Links ]
Chaves PT, Vazzoler G. Aspectos biólogicos de peixes amazônicos. III. Anatomia microscópica do esôfago, estômago e cecos pilóricos de Semaprochilodus insignis (Characiformes: Prochilodontidae). Acta Amazonica. 1984;14:343-353. [ Links ]
DeNiro MJ, Epstein S. Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Ac. 1978;42:495506. [ Links ]
Faria AC, Benedito E. Quality and digestibility of food ingested by various trophic fish groups in the Upper Paraná River floodplain. Rev Biol Trop. 2011;59:85-101. [ Links ]
Focken U, Becker K. Metabolic fractionation of stable carbon isotopes: implications of different proximate compositions for studies of the aquatic food webs using δ13C data. Oecologia. 1998;115(3):337-343. [ Links ]
Forsberg BR, Araujo-Lima CARM, Martinelli LA, Victoria RL, Bonassi JA. Autotrophic carbon sources for fish of the central Amazon. Ecology. 1993; 74:643-652. http://dx.doi.org/ 10.2307/ 1940793 [ Links ]
González RA, Wills A. Evaluación del desempeño de los alevinos del bocachico (Prochilodus magdalenae) Steindachneer 1878 sometidos a dos tipos de dieta. Rev Med Vet Zoot. 2003;50:50-56. [ Links ]
González RA, Wills FG. Relación proteína/energía digestible para alevinos de bocachico (prochilodus magdalenae), steindachner, 1878. Revista U.D.C.A. Actualidad & Divulgación Científica. 2005;8:111-118. [ Links ]
González RA, Wills GA. Evaluación de dietas isoenergéticas con varios niveles de proteína y de harina de pescado en alevinos de bocachico (Prochilodus magdalenae). Revista U.D.C.A. Actualidad y Divulgación Científica.2009;12:69-77. [ Links ]
Lopes CA, Cecilio-Benedito E, Martinelli LA. Variability in the carbon isotope signature of Prochilodus lineatus (Prochilodontidae, Characiformes) a bottom-feeding fish of the Neotropical region. J Fish Biol. 2007; 70:1649-1659. [ Links ]
Martínez-Porchas M, Martínez-Córdova LR, Ramos-Enríque R. Dinámica del crecimiento de peces y crustáceos. REDEVET. 2009;10:1-16. [ Links ]
Michener RH, and Schell DM. 1994. Stable isotope ratios as tracers in marine aquatic food webs. En: LAJTHA H, and Michener RH. (Editores). Stable isotopes in ecology and environmental science. Oxford: Blackwell Scientific. Cap.7, p. 138-157. [ Links ]
Minagawa M, Wada E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochim Cosmochim Ac. 1984;48:1135-1140. [ Links ]
Ministerio de Agricultura y Desarrollo Rural MADR y Corporación Colombiana Internacional CCI. 2010. Pesca y acuicultura Colombia. Bogotá [ Links ]. Overman NC, Parrish DL. Stable isotope composition of walleye: 15N accumulation with age and area-specific differences in δ13C. Can J Fish Aquat Sci. 2001;58:1253-1260. [ Links ]
Sacramento PA. 2014. Efeito da dieta sobre a taxa de turnover e o fator discriminante de carbono e nitrogênio em peixe detritívoro Neotropical Tese doutorado, Centro de Ciências Biológicas, Universidade Estadual de Maringá [ Links ].
Santiago LS, Silvera K, Andrade JL, Dawson TE. El uso de isotopos estables en biología tropical. Interciencia. 2005;30:536-542. [ Links ]
Urbano TG, Moreno C, Silva A, Santaime R. 2009. Engorde de coporo en lagunas de tierra, en el estado Delta Amacuro. INIA Divulga, Pesca Y Acuicultura. 7-10 p.p. [ Links ]
Van-Zander M, Rasmussen JB. Variation in δ15N and δ13C trophic fractionation: implications for aquatic food web studies. Limnol Oceanogr. 2001;46:2061-2066. [ Links ]
Yossa MI, Araujo-Lima CARM. Detritivory in two Amazonian fish species. J Fish Biol. 1998;52:1141-1153. [ Links ]
Yossa MI, 2002. Estratégia alimentar de peixes detritivoros da bacia do Amazonas e do Orinoco. Tese doutorado. Instituto Nacional do Amazônia-Inpa. Universidade do Amazonas, Manaus, Am, Brasil. [ Links ]
Yossa MI, Vasquez-Torres W. 2012. Resultados preliminares de fuentes de carbono en estanques piscícolas. Memórias XIII jornada de Acuicultura- IALL, 20-23. P.p [ Links ]
