











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
La biodiversidad es una de las palabras idóneas para definir a los ecosistemas acuáticos tropicales, debido a sus innumerables cuerpos hídricos que incluyen desde pequeños riachos a grandes ríos con diferentes tipos de agua y con diferentes características biológicas, químicas y físicas (Val et al., 2005). Un único cuerpo de agua puede ser susceptible a diversos cambios durante todo el año, incluso desapareciendo, en algunos casos, durante la estación seca (Junk et al., 1989; Almeida-Val et al., 1999) tal como ocurre en lagos de poca profundidad en el Amazonas (Sioli, 1984; Junk et al., 1989; Val y Almeida-Val, 1995). Geográficamente, la cuenca Amazónica cubre 7.05 millones de km2, ocupando aproximadamente el 39% de Sudamérica y alrededor del 72% se encuentra en Brasil (5.1 millones de km2) (Webb, 1995; Rossetti y Toledo, 1998). Posee una gran variedad de ambientes acuáticos, entre los cuales se destacan playas, riachuelos, cascadas y ríos de gran porte, bien sea de aguas claras, negras y blancas (Sioli, 1967; Lowe-McConnel, 1987).
Sus planicies de inundación y afluentes forman una red compleja de canales que a su vez se alteran conforme a la dinámica ambiental (Junk et al., 1989) promoviendo cambios limnológicos reflejados en los niveles de oxígeno, temperatura, disponibilidad alimenticia, refugio y áreas de cría para los peces (Junk et al., 1989; Nolan et al., 2009). Alteraciones en los parámetros ambientales y su interacción con factores endógenos en los peces, funcionan como un gatillo para eventos fisiológicos y comportamentales, desencadenando una serie de sucesos que intervienen en los procesos de migración, maduración de gametos y desove (Lowe-McConnel, 1987; Nahagama et al., 1995, 2008).
De acuerdo al proceso migratorio, se generaliza que está presente para todo el reino animal, pero en peces, dicho acontecimiento permite aprovechar la variabilidad de nuevos hábitats y evitar condiciones adversas (Dingle y Drake, 2007; Binder et al., 2011). Según Mehner y Kasprzak (2011) el término migración, se refiere los movimientos definidos por individuos o poblaciones (o parte de poblaciones) entre dos hábitats, que hasta cierto punto, es predecible temporalmente y tiene una periodicidad regular. Por lo tanto, la migración difiere de la dispersión en que los individuos hacen un viaje de regreso al hábitat inicial. Este tipo de desplazamientos inician con base a una respuesta que tuvo la especie ante un estímulo ambiental y finalizan en cuanto dicho estímulo ya no está presente (Zapata y Usma, 2013).
Publicaciones referentes al estudio de la ecología de peces amazónicos como los realizados por Lowe-McConnel (1987) y Barthem y Fabré (2004) se han enfocado en proponer categorías y estrategias particulares de manejo de los peces de acuerdo a las características ambientales donde viven y el tipo de movimiento ejercido (longitudinal y lateral) en el ecosistema acuático, dando a conocer tres tipos: especies sedentarias, especies migratorias que hacen uso las planicies inundables y especies migratorias que llevan a cabo movimientos de grandes distancias. Los peces son el grupo taxonómico de mayor importancia en la región amazónica, pero sus patrones ecológicos no se conocen completamente (Granado-Lorencio et al., 2005; Pazin et al., 2006).
Numerosas especies son capturadas con fines ornamentales y de consumo en el ríos amazónicos (Bayley y Petrere Jr, 1989; Batista y Petrere Jr, 2003) sin embargo, órdenes como los Characiformes y Siluriformes se han establecido como los de mayor prevalencia en las pesquerías y en las investigaciones de carácter ecológico, donde los aportes científicos sobre composición de la estructura trófica en lagos y canales principales de la cuenca han sido de gran relevancia (Junk et al., 1983; Soares et al., 1986; Petry et al., 2003; Cox-Fernandes et al., 2004). La riqueza de los Siluriformes representa hasta el 39% de las especies de la fauna Neotropical y la familia Pimelodidae es una de las más representativas, dentro de ella, algunas especies de los géneros Brachyplatystoma y Pseudoplatystoma son los que se destacan como migradores (Lowe-McConnel, 1987; Galvis et al., 1997; Barthem y Goulding, 1997).
Finalmente, aunque existe un interrogante cuando se intentan clasificar los diferentes tipos de migración en peces, algunos autores, hacen uso de ciertos términos para describirlos, entre estos: oceanódromos, potádromos y diádromos (Myers, 1949; McDowall, 1988). El objetivo de esta revisión ofrece algunas consideraciones generales sobre la migración de algunas especies de importancia en la familia Pimelodidae, nativos de la cuenca Amazónica, estrictamente considerados como potamodromos y su relación con los ciclos hidrológicos.
Importancia de la actividad pesquera en la Amazonia de Colombia y Brasil
La pesca hace parte intrínseca de la cultura ribereña, adaptándose a condiciones adversas en invierno y verano durante la subienda y bajanza de las aguas, pero en busca de satisfacer las necesidades de seguridad alimentaria, el pescador artesanal hace uso del producto de su trabajo para su subsistencia (Gualdrón, 2002; Diegues, 2004). En la Amazonia colombiana, esta labor es destacada por ser una de las actividades de impacto económico y cultural, que además de generar ingresos por la venta de pescado de consumo, también ha permitido incursionar en el mercado ornamental (Barthem et al., 1995; Fabré y Alonso, 1998; Agudelo et al., 2006).
Ante el anterior panorama, se desarrolla una recopilación de datos del comercio pesquero, cuya actividad es destacada en Leticia, Florencia, Puerto Leguízamo, Puerto Nariño y Mitú, pero es el municipio de Leticia que se ha caracterizado por ser uno de los principales centros de acopio de peces de cuero, teniendo también incidencia en parte del territorio peruano (Agudelo et al., 2006; De la Hoz y Manjarrés-Martínez, 2016). En los desembarcos de los Pimelodidae registrados por De la Hoz y Manjarrés-Martínez (2016) se encontraron especies como pintadillo tigre (Pseudoplatystoma tigrinum), dorado (Brachyplatystoma rousseauxii), amarillo (Zungaro zungaro), cajaro (Phractocephalus hemioliopterus), jugador (Brachyplatystoma juruense), pirabutón (Brachyplatystoma vaillantii), simi (Calophysus macropterus), barbiancho (Pinirampus pinirampus) y valentón (Brachyplatystoma filamentosum). Aun así, existe una problemática que se cierne en la Amazonia colombiana, como la carencia de lineamientos específicos que contribuyan al ordenamiento de la pesca artesanal (Agudelo, 2015) ya que tan sólo se han logrado estandarizar medidas enfocadas a las vedas, regulación de artes de pesca, áreas de pesca y tallas mínimas de captura, esto mismo acontece en lugares fronterizos de Perú y Brasil (Agudelo et al., 2009). Adicionalmente, una de las regulaciones implementadas entre Colombia y Brasil, fue la prohibición de la pesca y el comercio de C. macropterus, como consecuencia del uso indebido de carnadas de especies en peligro de extinción para su captura, como lo son los delfines Inia geoffrensis y Sotalia fluviatilis y los caimanes Melanosuchus niger y Caiman crocodilus, así, con esta medida se podría contribuir a la conservación de las especies, haciendo que C. macropterus fuera excluida dentro del mercado de bagres en la Amazonia brasilera (Iriarte y Marmontel, 2013; Brum et al., 2015).
Por otra parte, la pesca en la Amazonia de Brasil se encuentra desarrollada y posee intereses económicos, siendo el dorado (Brachyplatystoma rousseauxii) y el pirabutón (Brachyplatystoma vaillantii) los de mayor importancia y son las ciudades de Belén, Macapá, Santarém, Porto Velho, Lábrea, Tabatinga y Manaos, las zonas más sobresalientes (Batista et al., 2005; Barthem, 2013). En vista de esto, en Manaos y a través de ProVárzea se ha hecho el esfuerzo por evaluar el stock pesquero, permitiendo que últimamente sea un centro de acopio de importancia en la Amazonia (Almeida, 2006) debido a que la perspectiva de pesca pasó de ser artesanal a industrial. Sumado a esto, se han establecido áreas específicas para la delimitación de la actividad pesquera de grandes bagres, siendo desde Manaos (Brasil) hasta Iquitos (Perú) respectivamente (Barthem y Goulding, 1997). Aunque geográficamente, estos lugares han sido estratégicos para su comercialización, ecológicamente las distancias recorridas por los grandes bagres migradores indican que estas áreas están influenciadas por las cuatro fases hidrológicas del río Amazonas, permitiendo que se lleve a cabo su alimentación y reproducción (Rodríguez, 1991; Salinas, 1994).
Aproximación a la migración potamodroma
Anteriormente se pensaba que la migración en peces dulceacuícolas era relativamente extraña, ya que la mayoría de las especies se consideraban en gran parte sedentarias y mostraban movimientos locales limitados (Lucas y Baras, 2001). Aunque se conoce que las migraciones están basadas ecológicamente en su relación directa con los límites de aguas continentales o salobres, la migración potamodroma ocurre enteramente dentro de ecosistemas acuáticos continentales, abarcando hábitats lacustres como fluviales (Binder et al., 2011; Zapata y Usma, 2013; Obande et al., 2014). Excepciones notables a esta visión histórica son los grandes bagres migradores, como por ejemplo B. rousseauxii y B. vaillantii, las más importantes especies comerciales y las más estudiadas si de migraciones se refiere, caracterizados por su dependencia del estuario del río Amazonas como su vivero y ríos interiores como áreas de alimentación y desove (Barthem et al., 2017).
Evidencias significativas han mostrado que los peces de casi todos los hábitats dulceacuícolas y de todos los nichos ecológicos hacen movimientos migratorios en un rango de escalas espaciales y temporales. La capacidad mejorada para recopilar datos detallados sobre los movimientos de los peces también ha llevado a un cambio de los estudios descriptivos a investigaciones más basadas en hipótesis dirigidas a abordar algunas de las causas y consecuencias subyacentes de la migración (Olsson et al., 2006; Brodersen et al., 2008; Chapman et al., 2011; Skov et al., 2011). Generalmente, las especies enmarcadas en la migración potamodroma requieren de desplazamientos direccionados durante su fase reproductiva, incluyendo desoves estrictamente estacionales y relacionados con el pulso de inundación en el río (Petrere, 1985; Lowe-McConnell, 1987; Carolsfeld et al., 2004). De la misma manera, la influencia del aumento del caudal del agua juega un papel fundamental, pues los factores físicos y químicos inciden sobre la maduración y culminación del proceso reproductivo (Vazzoler, 1996; Jiménez-Segura et al., 2010).
Movimientos migratorios en la Amazonia
Desde hace algunas décadas, las investigaciones se han centrado en migraciones de larga distancia, no porque las migraciones a corta distancia sean ecológicamente o evolutivamente menos importantes, sino porque las migraciones en peces que recorren largas distancias imponen típicamente mayores desafíos comportamentales y fisiológicos que a los peces que realizan migraciones cortas (Binder et al., 2011; Duponchelle et al., 2016). Por tal motivo, el orden de los Siluriformes y su familia Pimelodidae ha tenido una implicación relevante en aspectos ecológicos y pesqueros, pues adicionalmente de que se encuentran en la mayoría de las cuencas de América del Sur, también han correspondido al segundo orden con mayor abundancia en especies para el Neotrópico, presentando mayores adaptaciones anatómicas y morfológicas (Salinas y Agudelo, 2000; Carolsfeld et al., 2004; Lasso et al., 2004, 2011).
De acuerdo a lo anterior, Lowe-McConnell (1987) logra recopilar información de acuerdo a la clasificación de los movimientos migratorios en los peces amazónicos, que están directamente relacionados con la variación de elevación y descenso del nivel de las aguas en los ríos: (Goulding, 1979; 1980; 1981; Ribeiro, 1985; Junk, 1984; Cox-Fernandes, 1988):
Migraciones motivadas por el cambio del nivel de las aguas: Estas migraciones son generalmente movimientos cortos en un espacio de centenas de metros a pocos kilómetros, siendo realizadas por las especies que colonizan las planicies de inundación. Son ocasionadas por los cambios del nivel de las aguas que permiten y facilitan el acceso a lugares de alimentación y de protección en el periodo de inundación y la salida de estos lugares durante el descenso, por lo general, están relacionadas con que han sido consideradas sedentarias.
Migraciones motivadas por fines reproductivos: Estas migraciones son realizadas por muchos Siluriformes y Characiformes, y ocurren generalmente en el inicio de la inundación. En algunos casos, las migraciones para desove se presentan solamente entre las planicies de inundación y los ríos, pero pueden ser realizadas río arriba hasta las cabeceras o más comúnmente, río abajo, siendo éste último caso, en el sentido de los ríos de aguas claras y negras para los de aguas blancas.
Migraciones motivadas por fines tróficos: Prácticamente todas las especies que realizan migraciones reproductivas también efectúan migraciones tróficas, en general, ellas son direccionadas río arriba, tal vez para compensar el desplazamiento de los ovocitos, de las larvas y de los juveniles, ocasionadas por el movimiento río abajo.
Ciclo hidrológico y pulso de inundación
El régimen de hidrológico durante el flujo de lluvias y posteriormente en el periodo de verano sobre la cuenca hidrográfica, se ha constituido como el factor ambiental de mayor importancia en la estructuración de las comunidades y en el funcionamiento de los sistemas de ríos y planicies de inundación (Lowe-McConnell, 1987; Junk et al., 1989) reflejándose sobre la integridad biótica y dinámica de los ensamblajes (Thomaz et al., 1997; Agostinho et al., 2001).
El ciclo hidrológico puede ser resumido en cuatro fases: aguas subiendo (inundación) aguas altas (punto máximo del caudal de agua), aguas bajando (descenso) y aguas bajas (sequía) (Lowe-McConnell, 1987). El pulso de inundación es el factor principal y responsable por la existencia de la productividad e interacción de la biota que vive en áreas periódicamente inundadas (Junk et al., 1989; Camargo y Esteves, 1996). Las precipitaciones intensas pueden inundar inmediatamente los ríos y arroyos de las cabeceras, pero los grandes ríos aguas abajo son menos influenciados por las lluvias locales y más por la precipitación total en las regiones aguas arriba que descienden en forma de una ola de inundación, ocasionando el desbordamiento de los ríos (Welcomme, 1985).
Esto podría ser un factor determinante en los cambios físicos y químicos del ambiente y como consecuencia las especies responden con una serie de adaptaciones morfológicas y fisiológicas (Junk et al., 1997) por ejemplo, en la incidencia reproductiva de individuos con diferentes patrones de desplazamiento, así lo describe Agostinho (2004) las especies de carácter sedentario con cuidado parental o fecundación interna parecen reproducirse independientemente del régimen de lluvias, salvo que las migradoras de corta distancia muestran tendencias intermedias. Pese a esto, hay una gran diferencia si se compara con los grandes migradores, puesto que presentan un desove total y tendencias a tener un elevado flujo genético como se ha estudiado en B. rousseauxii y B. vaillantii (Barthem et al., 1997; Batista, 2001) además, los peces evitan pérdidas de energía para realizar el desove en las regiones altas de la cuenca y así utilizar áreas inundables en las fases iniciales del desarrollo (Nakatani et al., 1997; Baumgartner et al., 2004; Agostinho et al., 2004).
La inundación es caracterizada por el aumento del nivel del río e igualmente por la acentuada expansión de los ambientes acuáticos en la planicie inundable, aunque la disponibilidad y la cantidad de nutrientes en zonas inundables están determinadas por el mismo ciclo (Goulding, 1980; Lowe-McConnell, 1987; Biodamaz, 2001). La importancia del pulso de inundación radica en consideración a su relación directa como uno de los factores principales que condicionan la biología y ecología de los ecosistemas amazónicos, ya que existe una dinámica en el intercambio de nutrientes y energía entre el componente acuático y terrestre (Junk et al., 1989). Esto sucede cuando el agua desborda el canal principal de los ríos hacia las zonas adyacentes y es en el inicio de este periodo que se presenta el desove de varias especies migratorias, para dispersar sus ovocitos en áreas recién inundadas (Lowe-McConnell, 1987; Montreuil et al., 2001) (Fig.1).
Luego de que la inundación cobije áreas específicas, como la vegetación baja y selva arbórea, los peces encuentran un ambiente ideal, bien sea por refugio y disponibilidad de alimento, algas planctónicas y perifíticas, que crecen en los ambientes lacustres y áreas inundadas menos sombreadas, así como también materia orgánica en descomposición, producto de algunas macrófitas acuáticas y restos de animales (Goulding, 1980; Lowe-McConnell, 1987; Goulding et al., 1988; Araújo-Lima et al., 1995; Junk et al., 1997). Durante este tiempo, los peces se encuentran muy dispersos, alimentándose intensamente y se caracteriza por ser el periodo de mayor tasa de crecimiento para las especies asociadas a las áreas inundables (Lowe-McConnel 1987; Junk, 1985; Oliveira, 1996; Fabré y Saint-Paul, 1998; Vieira, 1999) (Fig.1).
Durante el pico de aguas altas, el caudal del agua alcanza su punto máximo, es caracterizado por su corta duración y por un dominio total del ambiente acuático en las planicies inundables (Ribeiro y Petrere, 1990; Barthem y Fabré, 2004). En el descenso, los peces dan inicio a una acción de agrupamiento, las especies migradoras empiezan a formar cardúmenes y a iniciar su periodo de migración y dispersión (Fig.1) que durará todo el periodo de aguas bajas hasta el inicio de la inundación (Ribeiro y Petrere, 1990) no obstante, estas especies al dirigirse al canal principal del río o para los lagos, reducen su actividad alimentar, esta etapa es denominada como “invierno fisiológico” (Lowe-McConnel, 1987).
El periodo de aguas bajas es caracterizado por tener cierto grado de complejidad ecológica para la mayoría de las especies, pues el ambiente acuático es muy reducido, conllevando a una disminución del alimento y refugio (Fig.1). Estas áreas, que en algún momento estuvieron inundadas, forman un ambiente poco habitable e hipoxémico, como respuesta a la intensa descomposición de materia orgánica que consume el oxígeno disuelto en el agua y va liberando el gas sulfídrico, provocando una mortalidad de los peces que llegan a estos lugares (Santos, 1979; Lowe-McConnel, 1987). Estas variaciones en el ambiente, alteran la composición de las comunidades de peces en sistemas de tipo lacustre y de áreas de inundación, pero al mismo tiempo genera resiliencia en algunas especies para afrontar disturbios ocasionados por el ciclo hidrológico (Junk et al., 1989). Según Putman (1996) la resiliencia hace parte de un componente ecológico que ofrece una respuesta sobre cómo un disturbio puede modificar el comportamiento de organismos en su respectivo ambiente. Investigaciones realizadas por Bayley y Osborne (1993) y Humphries y Baldwin (2003) indican que ante condiciones adversas, las poblaciones de organismos aparentemente tienden a recuperarse y se reestablecen con rapidez después de ciertos acontecimientos naturales.
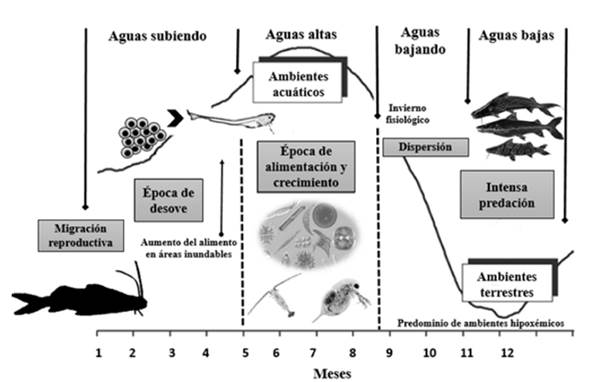
Figura 1 Dinámica del ciclo hidrológico sobre la migración en bagres (Adaptado de Lowe-McConnel, 1987; Barthem y Fabré 2003).
La fase de aguas bajas es un periodo mucho más favorable para los depredadores, que van en busca de cardúmenes de Characiformes y otros Siluriformes de menor talla, que en ese preciso instante estarán migrando en el canal del río o confinados en los lagos (Lowe-McConnel, 1987; Barthem y Goulding, 1997). A pesar de lo anterior, los peces desarrollaron durante el proceso evolutivo adaptaciones comportamentales, las cuales se modifican acuerdo a las condiciones del ambiente (Val y Almeida-Val, 1995). Con relación a las actividades de predación, que se presentan en aguas bajas por parte de los grandes bagres, Reid (1986) observó que los juveniles del cucharo (Sorubim lima) un pimelódidos de tamaño medio posee una coloración críptica, característica que permite que la coloración del pez se confunda con la del medio que lo rodea, haciendo que sea menos vulnerable en las planicies de inundación, es posible que sea una adaptación necesaria para los pimelódidos que usan las planicies de inundación y el canal principal del río en el inicio de sus vidas (Lowe-McConnel, 1987).
Clasificación de bagres migratorios
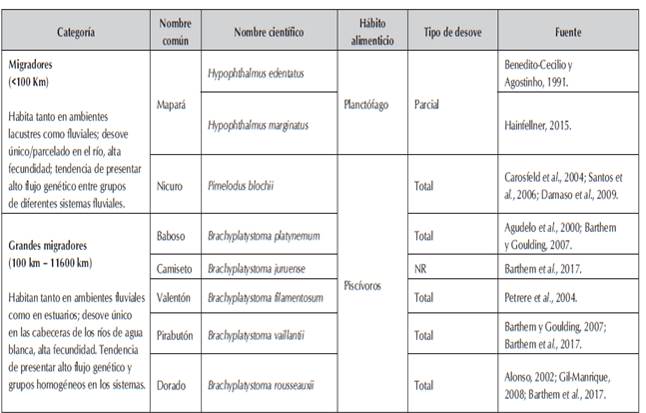
Las características del ambiente y el comportamiento de los peces, permite diferenciar dos grupos de especies, migradores y grandes migradores (Tabla 1) los migradores que hacen uso de los ambientes lacustres y fluviales, y los grandes migradores, relacionadas principalmente con el canal de los ríos y en algunos casos el uso de estuarios (Barthem et al., 1997; Batista, 2000).
Acciones antrópicas
La elevada capturabilidad en determinadas fases migratorias, explican el gran interés de la pesca comercial sobre las poblaciones que presentan comportamientos migratorios, no obstante, las actividades antrópicas mediante la construcción de barreras que irrumpen las rutas migratorias de los peces, pueden perjudicar la abundancia del stock explorado, la geomorfología fluvial que a su vez puede alterar la erosión, el transporte y la deposición de sedimentos (Carvalho y Merona, 1986; Schilt, 2007).
La construcción de represas, además de modificar las características físicas y químicas del agua y controlar su flujo (Agostinho et al., 2007) impide el libre tránsito de los peces por el río, haciendo que muchas especies no puedan efectuar su reproducción, constituyéndose como uno de los mayores peligros para la ictiofauna, eliminando bien sea especies diádromas y potamodromas de diferentes cuencas fluviales (Agostinho et al., 2005; Williams et al., 2012). De tal manera que, las estrategias desarrolladas por los peces muchas veces son afectadas por alteraciones en el ambiente, tales como las barreras que transforman ambientes lóticos en lénticos (Giamas et al., 2004) obligando a que las comunidades bióticas de su área de influencia deban adaptarse a nuevas condiciones para no ser eliminadas del sistema (Braga, 2001) además, como lo afirman Fearnside et al., (2014) las modificaciones en el hábitat principalmente por medio de represamientos, disminuye la variabilidad genética de las especies, reduciendo las tasas de crecimiento poblacional y aumentando en gran medida el riesgo de extinción.
Con el objetivo de mitigar el impacto negativo de las barreras sobre la migración de los peces, fueron construidas en Brasil, a partir del inicio del siglo XX, innumerables escaleras de peces, a fin de posibilitar la libre migración, río arriba y en la época reproductiva, pero a pesar de esto, su implementación ha sido realizada sin criterios técnicos, trayendo como consecuencia riesgos directos para la conservación de la ictiofauna local (Silva et al., 2012). Según Santos et al., (2007) el sistema de transposición de peces es de tipo escalera con escalón, este sistema parece ser mucho más simple si es comparado con otros modelos. Se constituye en una serie de depósitos o tanques escalonados secuencialmente, en donde los depósitos se separan por escalones. En la migración, los peces poco a poco van atravesando los escalones, pasando entre los depósitos o tanques durante la piracema, nadando por la lámina de agua bajando o saltando (Santos et al., 2007). Los escalones tienen la finalidad de controlar la permanencia de niveles de agua y esparcir la energía, haciendo que ésta sea disipada para favorecer la ascensión de los peces sin causar su cansancio (Makrakis et al., 2011) pero se requiere que las técnicas empleadas incluyan monitorización de la recolección cualitativa de informaciones biológicas que indiquen cuantos y de qué forma los peces transponen el obstáculo (Pompeu et al., 2012).
Conclusión
La cuenca Amazónica ha sido una de los sistemas más preservados del mundo y es caracterizado por presentar alto endemismo y diversidad de especies de peces con diferentes aspectos reproductivos y comportamentales, pero infortunadamente, los elevados costos de este tipo de investigaciones implican desconocimiento de sus rutas migratorias. Igualmente, estudios recientes han indicado que las pesquerías podrían estar disminuyendo debido a la sobrepesca y que las barreras impuestas por la acción antrópica en los cursos de los ríos aguas arriba amenazan constantemente las migraciones reproductivas. Por consiguiente, en la última década, una serie de reportes genético-geográficos y microquímicos han surgido para dar respuesta a estos sucesos migratorios y de alta dispersión a lo largo del ciclo de vida de poblaciones de bagres pimelódidos en los ecosistemas acuáticos de la cuenca amazónica. Entre ellos, se puede mencionar lo realizado por Telles et al., (2014), quienes a través de análisis espaciales para investigar la variación genética molecular (utilizando 7 marcadores de microsatélites) de Pseudoplatystoma punctifer, hicieron uso de muestras obtenidas de 15 localidades a lo largo del río Madeira y Solimões, obteniendo como resultado una disminución en la similitud genética, la cual varía entre la distancia de las poblaciones, así, se concluyó, que el patrón revelado tiene implicaciones importantes para establecer estrategias para mantener la diversidad genética en la especie, especialmente teniendo en cuenta las amenazas por los impactos causados por la construcción de grandes presas en este sistema fluvial. Luego Hegg et al., (2015), mediante microquímica de otolitos, describen los comportamientos migratorios de las tres especies de bagres migratorias más populares y comercialmente importantes, dorado (B. rousseauxii), pirabutón (B. vaillantii) y valentón (B. filamentosum). Las colectas se realizaron en la desembocadura del río Amazonas y la Amazonia central y se usaron firmas de isótopos de estroncio (87Sr/ 86Sr) registradas en los otolitos para determinar la ubicación de la cría temprana y posterior. Los resultados indicaron que los juveniles exhiben diversas estrategias de crianza, tanto los que se desarrollan en ambientes río arriba y en el estuario. Esto contrasta con el entendimiento prevaleciente de que los juveniles se crían en el estuario antes de migrar río arriba, sin embargo, cuenta con el respaldo de algunos datos de pesquerías que han indicado la presencia de historias de vida de desove y cría alternativos. Todo lo anterior, conduce a formular investigaciones encaminadas a fortalecer el conocimiento sobre comportamiento migratorio de las especies, ya que a partir de ellas se podrían identificar zonas que no han sido exploradas y que funcionan como áreas de llegada para el desove y alimentación de especies con importancia comercial para las pesquerías, permitiendo identificar aún más sus estrategias reproductivas.

