











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
La Orinoquia es una vasta región del territorio colombiano (26 millones de hectáreas) que tiene una importancia estratégica para el desarrollo del país; más del 53% de este territorio corresponde a la Altillanura bien drenada (Mejía, 1984), allí se ubica una población de 664.604 animales especialmente de cría (Compes, 2014) que produce un alto porcentaje de machos para la ceba que surten los grandes centros de consumo del país.
El sistema de cría se maneja de forma extensiva y se ve afectado entre otros factores por la estacionalidad de las lluvias, la pobre fertilidad de los suelos, el bajo contenido de materia orgánica, los altos niveles de aluminio intercambiable, lo que lleva a una escasa disponibilidad de nutrientes para los forrajes, todo lo cual, se traduce en bajos niveles productivos (Rivas et al., 2004). Después de analizar diferentes especies forrajeras en la Altillanura, se encontraron deficiencias, especialmente de Cu y Zn; estas mismas deficiencias se identificaron en los tejidos de los animales (Laredo y Gómez 1980; Gómez y Laredo 1983).
Durante la gestación en el bovino ocurren importantes cambios metabólicos, hormonales y reproductivos (Chilliard et al., 1998; Kindahl et al., 2002) que se ven influenciados por la nutrición, pero solo se reflejan al momento del parto o durante el posparto con la aparición de las enfermedades metabólicas o asociadas a la producción (Barth et al., 2011;Smith et al., 2017).
Los perfiles metabólicos (PM) se utilizaron por primera vez en ganado de leche en Inglaterra (Payne et al., 1970), desde entonces, se han constituido en una herramienta complementaria en el diagnóstico de las enfermedades asociadas a la producción (Quintella et al., 2011). El fundamento de esta técnica se basa en el análisis de ciertos componentes de la sangre como indicadores de imbalances nutricionales, lo que permite mejorar la alimentación, detectar problemas subclínicos y prevenir las enfermedades especialmente durante el periodo de transición (Butler y Smith, 1989). De forma paralela, la estimación de la condición corporal (CC) ha sido recomendada para evaluar el manejo de la nutrición (Rae et al., 1996) ya que permite medir la cantidad de energía metabólica almacenada en forma de grasa subcutánea y en el músculo (Houghton et al., 1990).
La variación en los cambios de la CC pueden reflejar el estado nutricional; en consecuencia una baja CC puede tener un valor predictivo de los trastornos metabólicos, fallas en la reproducción y la producción de leche (Edmonson et al., 1989). Por otro lado, la restricción energética durante la gestación disminuye la CC al parto y reduce el peso de los terneros al nacimiento (Freetly et al., 2000) por consiguiente el estado nutricional de la madre, es el principal factor en el desarrollo y función del sistema de órganos fetales (Caton y Hess, 2010). Igualmente, la baja ingestión de energía y proteína durante la gestación tardía, afecta el desarrollo de la placenta, que conduce al nacimiento de terneros débiles y un aumento en la retención de la placenta (Zhang et al., 2002).
El objetivo de este estudio fue conocer los valores de determinados metabolitos sanguíneos, la condición corporal y el peso vivo de vacas de cría, durante cada uno de los trimestres de gestación en una finca de nivel tecnológico medio de la Altillanura Colombiana.
Materiales y métodos
Ubicación y Manejo del Sistema productivo
El estudio se llevó a cabo en una finca ubicada en el municipio de Puerto López, Meta, Colombia (4° 6‘ 7“ latitud N y 72° 54‘ 24“ longitud Oeste), entre los meses de Enero-Diciembre de 2013. Se utilizaron 19 vacas multíparas de un grupo de 159, entre 3 a 4 partos de la raza Cebú (C) y cruces absorbentes de Cebú con Simmental (CxS) con edades entre los 5 y 6 años. Los animales seleccionados fueron sometidos a un programa de inseminación a tiempo fijo (IATF) siguiendo el protocolo sugerido por Cutaia et al., (2003) y confirmada la preñez por ultrasonido al día 50 posinseminación (SSD-500 Aloka Co, Tokyo, Japan).
Los animales durante el estudio se mantuvieron en pastoreo continuo en praderas de Brachiaria humidicola cv. dyctioneura (pasto llanero), Brachiaria brizantha cv. marandú y leguminosas nativas perennes como Desmodium spp y Centrosema spp., sal mineralizada al 8% de fósforo y agua permanente.
Toma de muestras, análisis de metabolitos, peso vivo y condición corporal
De cada vaca se tomaron 2 muestras de 5.0 ml por punción coccígea con y sin anticoagulante en tubos vacutainer estériles, transcurrido 30 minutos de su obtención, las muestras con anticoagulante fueron centrifugadas a 3.000 rpm durante 10 minutos y el suero obtenido conservado a -70°C. El muestreo se realizó a las 7:00 horas para cada uno de los trimestres de gestación T1: gestación temprana (segunda semana del mes de mayo), T2: gestación media (segunda semana de Septiembre) y T3: gestación tardía (segunda semana de Diciembre). La Glu, Col, Alb, Ptt, Hb, Ca, P, Cu y Mg se determinaron por el método colorimétrico de punto final, mientras la urea, Zn y la AST mediante método cinético - enzimático, las Glo por la diferencia entre Ptt y Alb, Hto por tubo capilar. Los reactivos usados fueron Spinreact (Spinreact, Girona, España) y la lectura de las pruebas, se realizó en un espectrofotómetro semiautomatico (Mindray Ba-88 A, Shenzhen, China). Simultáneamente al muestreo sanguíneo se realizó el pesaje de las vacas en una báscula con capacidad de 1.000kg (Prometalico) y evaluada la CC en escala de 1-9 en 1: emanciada y 9:obesa (Morrison et al., 1999)
Análisis estadístico
Los resultados fueron analizados mediante estadística descriptiva para cada una de las variables en estudio, determinando el rango, promedio y desviación estándar (Weisbrot, 1985), verificándo la distribución normal de los datos por la prueba de Kolmogorov-Smirnov,
Se determinó el rango de referencia para cada metabolito implicados en el estudio utilizando la metodología descrita por Bermes et al., (1976) y Weisbrot (1985), obteniendo los intervalos de confianza al 95%. Además, se realizaron correlaciones de Pearson y Stephens (1964), para determinar el grado de asociación entre las variables peso, condición corporal y etapas de la gestación con los valores de los metabolitos en estudio.
Resultados y discusión
Los valores promedios totales, el error estándar y los intervalos de confianza de los metabolitos analizados se observan en la Tabla 1. Los valores promedio totales de Glu (43,76 mg/dL) fueron inferiores a los reportados en razas criollas colombianas (46,98 mg/dL) (Campos et al., 2004), vacas Jersey (48,1 ng/dL) a los 8 dias PP (Roos et al., 2008), vacas rubia Gallega al primer mes posparto (57,38 mg/dL) (Quintela et al., 2011) y en general para la especie (45-75 mg/dl) (Kaneko, 1989).
Tabla 1 Concentraciones medias, error estándar e IC para los metabolitos estudiados en vacas de un sistema de producción de cría en la Altillanura colombiana.
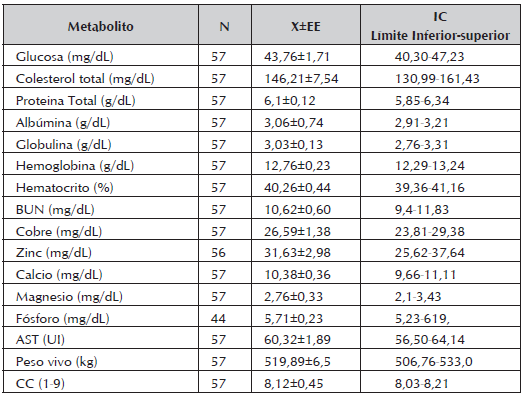
◊ Intervalo de confianza (95%).
Los valores de Glu fueron diferentes para los tres trimestres de gestación (P<0,05), Tabla 2, encontrando los mayores valores en T2 (47,43 mg/dl), estos niveles pueden ser atribuibles a una mayor demanda por parte del feto. Consecuentemente los menores valores en T1 (37,04 ng/dL) y T3 (43,63 mg/dL) podrían estar asociados en el primer caso a causas nutricionales, ya que este periodo coincidió con la época de sequía en donde las deficiencias de energía y proteína fueron marcadas debido a la escasez de nutrientes en el forraje y en el segundo caso, a la alta demanda por parte del feto en donde el crecimiento fetal es mayor. Los valores en T3 son menores a los reportados en vacas Brahman (50,81 ng/dL) durante las últimas semanas de gestación (Villa et al., 2011).
Tabla 2 Concentraciones medias, desviación estándar y rango para los metabolitos energéticos y nitrogenados durante los trimestres de la gestación en vacas de cría en la Altillanura colombiana.
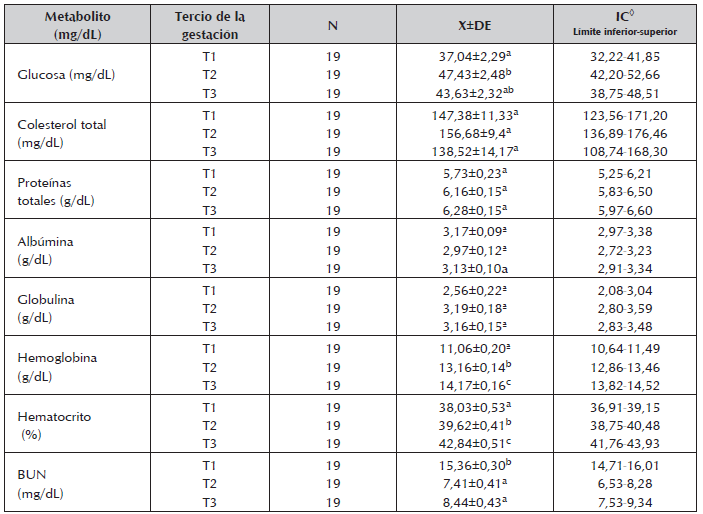
◊ Intervalo de confianza (95%).
Los niveles de Glu presentaron una correlación negativa (r=-0,388; P<0,01) con el BUN es decir, que cuando la Glu aumentaba el BUN disminuía. Obeidat et al., 2002 asociaron los elevados niveles de nitrógeno ureico en suero con un bajo consumo energético o con condiciones severas de desnutrición.
Contrariamente, la Glu plasmática presentó una correlación positiva con la AST (r=0,316; P<0,01), es decir que cuando aumentaba la Glu aumentaba la AST. Dado que la síntesis de Glu sanguínea en los rumiantes depende totalmente de la función hepática, cualquier alteración en la actividad de las enzimas indicadoras del daño hepático, tienen mayor valor diagnóstico que la propia glicemia (Álvarez, 2008).
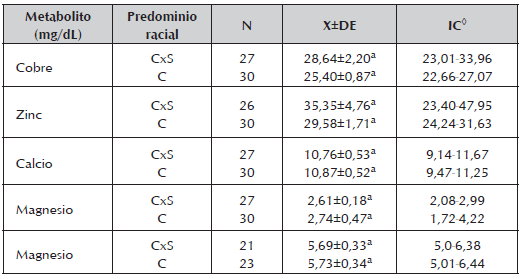
El Col no mostró diferencias (P>0,05) durante los trimestres de gestación, sin embargo en T3, los niveles fueron más bajos (Tabla 2), coincidiendo con los arreglos metabólicos previos al parto como la movilización de la grasa, crecimiento fetal y el inicio de la lactancia (Bell 1995). Una tendencia similar fue encontrada por Shope y Gowen (1928), quienes a su vez observaron una gran variación individual. Los niveles de Col en función de grupo racial presentaron diferencias estadísticas (CxS:129,56±72 mg/dL Vs C: 163,69±9,35 mg/dL P<0,05) lo cual sugiere que el ganado Cebú podrían tener mayores valores de Col en sangre, como ha sido sugerido (Álvarez, 2008).
Barrios et al., 2013 encontraron en la región centro occidental de Venezuela una media de 118±35 mg/dL para el Col, valor muy inferior al encontrado en este estudio. De igual forma, Giraldo et al., 2008 encontraron bajos niveles de Col en vacas Brahman (108,2mg/dL) cuatro semana antes del parto con una CC ≤ a 8.
Respecto a las Ptt los valores promedio se encontraron por debajo del rango reportado para la especie (6,74-74,6 ng/dL). Además no se encontró diferencias en los niveles por trimestre de gestación. En tanto que la síntesis de proteínas esta bajo control genético, se esperaría encontrar diferencias entre los dos grupos raciales, sin embargo esta situación no fue observada.
Durante la gestación el total de las Ptt disminuyen debido a una disminución de la Alb y un ligero incremento de las Glob, mas cerca al parto hay un rápido incremento en las a-globulinas y un aumento en la totalidad de las Ptt plasmáticas; en la lactación el total de las proteínas plasmáticas disminuye debido a la disminución de la Alb (Kaneko, 1989). Los niveles de Alb (3,06ng/dL) encontrados en nuestro estudio, se encontró dentro de los valores de referencia para la especie (3,03-3,55 ng/dL). Al igual que la Alb y las Glo no se encontraron diferencia entre los trimestres de gestación, ni por grupo racial. Generalmente un perfil anormal de proteínas se ha asociado con alguna enfermedad (Kaneko, 1989), situación que no fue observado en este estudio.
A medida que avanzaba la gestación se encontró un aumento en la Hem y el Hto, encontrándo diferencias (P<0,05) entre los tercios de la gestación (Tabla 2). Los valores promedios obtenidos para la Hem y Hto en T3 son mayores a los reportados por Zambrano y Marques (2009) en vacas Gyrolando, atribuidos posiblemente a una mayor demanda de aminoácidos necesarios para la síntesis de proteínas, que hace reducir la síntesis de otras proteínas (Contreras, 2000). De forma contraria se ha visto que la Hem disminuye significativamente durante la mitad de la gestación con los menores valores durante el segundo mes (Calvo et al., 1989), esta disminución ocurre por el ingreso de la Hem de la madre dentro de la circulación fetal, apoyada por la dilución de la sangre como consecuencia del aumento del volumen del plasma (Singh et al., 1991).
Los valores del Hto de este estudio fueron superiores a lo reportado en vacas Brahman y cruzadas Brahman x Sanmartinero que pastaban sabanas nativas similares a las utilizadas por los animales de este estudio (Vera y Rámirez-Restrepo, 2017). Con respecto a la Hem los valores son mayores a los encontrados en vacas Sahiwal durante los trimestres de gestación (Roy et al., 2010). De todas formas los resultados tanto de la Hem y Hto encontrados en nuestro estudio deben ser analizados con cuidado puesto que se conoce que la bioquímica sanguínea y los componentes celulares se ven influenciados por varios factores entre ellos, la raza, el sexo, la edad, el comportamiento, el manejo, los cambios fisiológicos y el periodo del dia (Taylor-Macallister, 1997). Las Ptt presentaron una correlación positiva con las Glo (r= 0,848; P<0,01), mientras se observó una correlación negativa entre la Alb y Glo (r=-0,383; P<0,01)
Se encontró una correlación positiva entre la Hem y Hto (r=0,781; P<0,01). De igual forma, estos dos metabolitos presentaron una correlación positiva con la CC y el PV (r=0,510, r=0,397; P<0,01 respectivamente). De forma contraria, el BUN presentó una correlación negativa con la Hem, Hto, CC, y PV (r=-0,711, r=-0,419, r=-0,430 y r=-0,441; P<0,01 respectivamente) y una correlación positiva con el Ca(r=0,275; P<0,05).
El BUN varió para los diferentes trimestres de gestación (P<0,05), (Tabla 2) presentando las concentraciones mas altas en T1, periodo que coincidió con la época final de la sequía, la cual es crítica para el crecimiento de las pasturas en esta región del país. Las concentraciones más bajas de BUN se observaron en T2 y T3, lo que puede estar relacionado con la baja calidad del pasto (época seca). Sin embargo, niveles altos de BUN pueden estar asociados a la oferta de Proteína cruda (PC) ofrecida durante el pastoreo, al bajo consumo de energía y/o condiciones severas de desnutrición (Obeidat et al., 2002), se puede inferir entonces, que los altos niveles de BUN durante T1 pudieron estar influenciados por la cantidad de PC presente en la pasturas y el consumo de energía (EE), descartando un estado de desnutrición ya que, los animales presentaron una buena condición corporal (7,93±0,60). Los niveles de BUN durante T1 son similares a los reportados por Tabeleão et al., 2006 en hembras mestizas (Bos taurus x Bos Indicus) en Río Grande/RS, Brasil y superiores a los reportados por Ceballos et al., 2002 en ganado lechero del trópico alto del eje cafetero colombiano e inferiores a las encontradas durante el preparto en vacas Rubia gallega (Quintela et al., 2011). Resultados similares para el BUN encontrados en este estudio han sido reportados por Romero-Treviño 2011 en vacas multíparas Pardo Suizo x Cebú en lactancia con una concentración de 10.6 mg/dL e inferior a la reportada por Barrios et al., 2013. Respecto a los niveles de acuerdo con el grupo racial no se encontró diferencia (p>0.05).
Las concentraciones séricas de hemoglobina, hematocrito y BUN, se observaron variaciones según el tercio de la gestación (Tabla 2).
Los niveles promedios de Cu y Zn encontrados en este estudio, están por debajo de los niveles críticos (Cu= < 0,57 mg/ml; Zn=< 0.6mg/ml) (Noaman et al., 2012) para todos los trimestres de gestación evaluados sin diferencias entre trimestres ni por grupo racial.
Otros estudios han encontrado bajos niveles de Cu en vacas preñadas que coinciden con nuestro estudio, lo cual se ha atribuido a que la preñez es dependiente del Cu, dadas las demandas adicionales para atender esta condición (Noaman et al., 2012). Estos resultados coinciden a la vez con los bajos niveles al final de la gestación (Slavick et al., 2006). Es conocido que el Cu es importante para diversas funciones corporales como la formación de hemoglobina, crecimiento, prevención de trastornos patológicos, actividad de las metaloenzimas esenciales en la reproducción normal, entre otras (Underwood y Suttle, 1999), por lo que las deficiencias de este mineral puede afectar estos procesos.
Estudios en EU en un sistema de producción vaca-ternero bajo condiciones similares a nuestro estudio, encontraron que los forrajes no eran normales en Zn y marginales en Cu con baja disponibilidad de Cu debido a las altas concentraciones de Fe y Mo (Corah et al., 1996; Mortimer et al., 1999). Esta situación es concordante bajo las condiciones de la Altillanura colombiana, caracterizadas por los altos niveles de Fe. De otro lado se ha asociado las deficiencias de Cu con los bajos niveles contenidos en el alimento (Ward, 1978)
Respecto al Zn, más de 200 enzimas dependen de este mineral, por lo que participa en la biosintesis de acidos nucleicos, el desarrollo esquelético, la estabilizacion del DNA, RNA y ribosomas, la replicación y diferenciación celular, el metabolismo de carbohidratos, síntesis y actividad de hormonas entre otras (Ahola, 2004). En Ganado alimentado con una dieta baja en Zn, la rata de absorción solo fue de 50% (Stake et al., 1975). Los bajos niveles de Cu y Zn podría sugerir alguna deficiencia subclínica sin que se reconocieran desórdenes clínicos en los animales analizados, tal como ha sido observado en otro estudio (Fiorentin et al., 2018). Otra consideración importante y que no fue analizada en este estudio es la variación que presentan estos minerales de acuerdo con la época. De tal forma, se han encontrado elevadas concentraciones sericas de Cu durante el invierno frente al verano (Noaman et al., 2012), lo que plantea el interrogante si se debería usar una sal mineralizada con la misma composición química todo el año.
Los niveles de Ca, Mg y P se encontraron dentro de los niveles normales sugeridos por González y Silva, 2006 y Álvarez 2008 y los encontrados por Barrios et al., 2013. Los valores de Cu (26,59 ± 1,38mg/dL) y Zn (31,63 ± 2,98mg/dL) no presentaron diferencias para el tipo racial (tabla 5), aunque se consideran como valores críticos para los bovinos.
En este caso, la aparente discrepancia entre los valores séricos bajos, niveles al límite y superior de Zn e inferior de Cu en las praderas, posiblemente se debe a las altas concentraciones de Fe en el suelo y el forraje que impiden una mejor disponibilidad de P para las plantas, de igual forma, se presenta una baja absorción de Cu y Zn en el ganado (Fordyce et al., 1996), resultados que se podrían asimilar a los encontrados en este estudio.
Tabla 3 Concentraciones medias, desviación estándar y rango para los metabolitos energéticos y nitrogenados encontrados durante la gestación en vacas cruzadas Cebú x Simmental (CxS) y Cebú (C), de un sistema de producción de cría en la Altillanura colombiana.
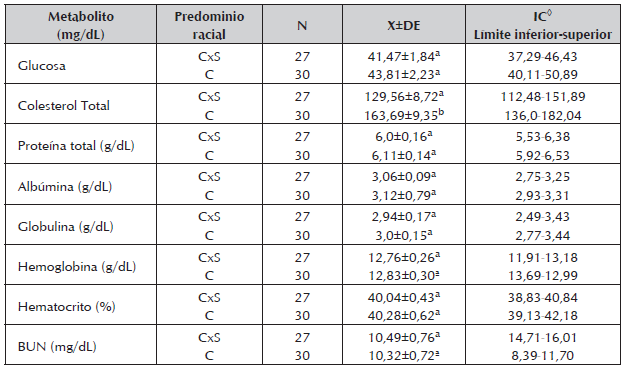
◊ Intervalo de confianza (95%).
Por otro lado, se conoce que el Fe compite por los sitios de absorción a nivel intestinal con el Cu. La presencia en este estudio de animales con niveles de Cu y Zn por debajo de sus requerimientos hace pensar que estarían predispuestos a desarrollar una carencia secundaria de cobre.
La concentración del Zn sérico presentó una correlación negativa con la AST (r= -0,299; P <0,05), lo que indica que aumentos de la AST se corresponden con disminuciones del Zn. De igual forma el Ca con el Zn (r= -0,29; P <0,05) y una correlación positiva de este último con el BUN (r=0,275; P<0,05).
La concentración total de Ca (10,3±0,3ng/dL) fue superior a los limites reportados para la especie (8,0-10,5 mg/dL) con los mayores valores en T1 (11,98 mg/dL) y T3 (11,38mg/dL) los cuales fueron diferentes a T2 (9,09mg/dL) (P<0,05) muy por debajo del límite inferior (Tabla 4), indicando tal vez una mayor demanda y consumo en este trimestre. Sin embargo, los niveles séricos de BUN y Ca durante T2 pueden estar indicando una carencia de proteína bruta y de Ca o una mayor demanda por parte del feto durante este trimestre.
Durante T1 y T3 los valores séricos del Ca siempre se mantuvieron por encima de los limites normales (8,5 - 9,0 mg/dl) (Kaneko et al., 1997). Con nivel deficiente de Ca en el suelo, un porcentaje de Ca bajo durante la época de lluvias y un porcentaje superior durante la sequía de las pasturas, los valores séricos encontrados en este estudio se deben fundamentalmente al eficiente mecanismo homeostático que mantuvo el nivel del Ca sérico dentro de límites fisiológico (Said et al., 1964.) y a la suplementación mineral en fósforo que recibieron los animales (sal mineralizada al 8%).
Se ha identificado que el exceso de Ca disminuye la absorción de Zn (Graham 1991), por lo que es posible que esta situación haya ocurrido en este estudio, sin que se observe que los niveles de Ca durante la gestación variarán significativamente (Tabla. 4) pudiendo afectar la absorción del Zn. Durante T1 (época seca), los valores séricos de Cu, Ca y P resultaron mayores (Tabla 4), y superiores a los reportados por Depablos et al., 2009. Los valores de Mg no presentaron diferencias por trimestre de gestación, ni por grupo racial y se encontraron dentro de los niveles normales para vacas de carne (1,2-3,5 ng/dL) (Church et al., 1978). Similar comportamiento presentaron los niveles de fosforo.
Tabla 4 Concentraciones medias, desviación estándar y rango para los metabolitos minerales encontrados durante los trimestres de la gestación en vacas de cría en la Altillanura Colombiana.
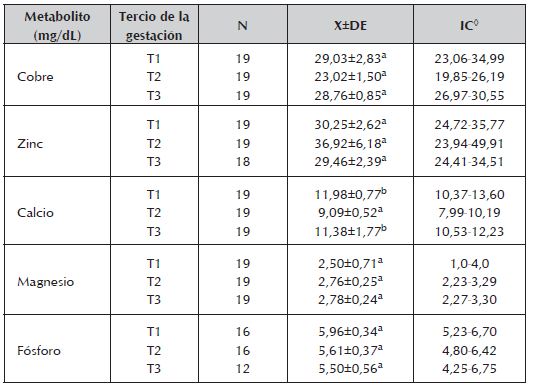
◊ Intervalo de confianza (95%).
La concentración promedio de P sérico durante la gestación y durante los trimestres de la gestación se encontraron dentro de los rangos sugeridos para ganado bovino (Kaneko et al., 1997). Con deficiencias de fósforo en suelo y en las pasturas, los valores normales séricos de fósforo se podrían explicar por suplementación mineral en fósforo recibida por los animales durante el estudio (sal mineralizada al 8%). En general se puede decir que los minerales deben ser analizados en su conjunto, debido a sus múltiples interacciones, tal como lo demostró un estudio en ratas preñadas en donde la deficiencia de Fe tuvo efectos opuestos sobre los niveles de Cu y Zn en el hígado fetal, lo cual puede afectar el metabolismo de estos nutrientes con graves consecuencias para el desarrollo fetal y el neonato (Cottin et al., 2019).
Tabla 5 Concentraciones medias, desviación estándar y rango para los metabolitos minerales encontrados durante la gestación en vacas cruzadas Cebú x Simmental (CxS) y Cebú (C), de un sistema de producción de cría en la Altillanura colombiana.
◊ Intervalo de confianza (95%).
No se encontraron diferencias significativas en la concentración sanguínea de la AST durante los trimestres de la gestación, ni por grupo racial (Tabla 6 y 7). La AST es un buen indicador del funcionamiento hepático, aumentos de la AST sérica se encuentran asociados con daño hepático, como en el caso de las hipoglicemias (González y Silva, 2006).
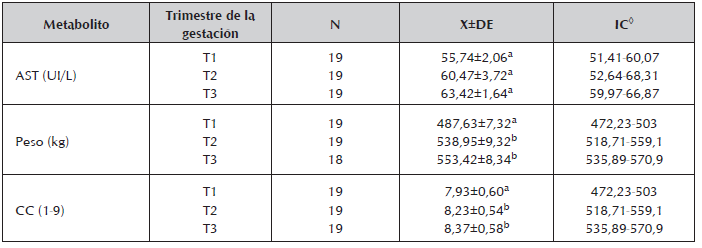
Tabla 6 Concentraciones medias, desviación estándar y rango para los metabolitos AST, peso vivo y CC durante los trimestres de la gestación en vacas de cría en la Altillanura Colombiana.
◊ Intervalo de confianza (95%).
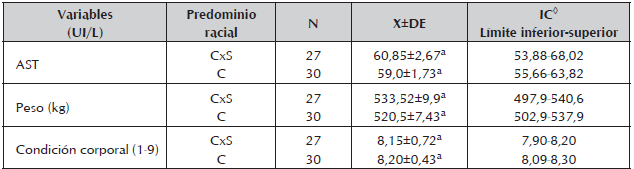
Tabla 7 Concentraciones medias, desviación estándar y rango para AST, peso vivo y condición corporal encontrados durante la gestación en vacas cruzadas Cebú x Simmental (CxS) y Cebú (C), de un sistema de producción de cría en la Altillanura colombiana.
◊ Intervalo de confianza (95%).
Se presentó un aumento de la AST a medida que avanzaba la gestación (tabla 6). La concentración sérica promedio de AST encontrada en este estudio fue superior a la reportada por Roos et al., 2008 en vacas Jersey al sur de Rio Grande do Sul e inferior a las encontradas por Quintela et al., 2011 en vacas de la raza Rubia Gallega y por Coppo et al., en 2000 en bovinos cruzados Cebú x Europeo (30UI/L). Contrariamente la disminución de la AST a través de la gestación, se ha asociado con cambios hormonales y uterinos (Roy et al., 2010).
Barini 2007, en un estudio con ganado Curraleiro, encontró que los niveles séricos de AST eran directamente proporcionales a la edad, lo que evidencia que los PM se ven influenciados por diferentes factores entre ellos la edad.
Se observaron variaciones en el peso y la CC durante los trimestres de la gestación (Tabla 6), pero no por predominio racial (Tabla 7). Los mayores pesos se observaron en las vacas con predominio racial CxS (533,52±9,90 kg) frente a las Cebú (520,5±7,43 kg), contrario a lo encontrado para la CC, en donde las vacas con predominio racial Cebú presentaron la mayor CC (8,20±0,43) que las CxS (8,15±0,72). En razón de que tanto el PV como la CC aumentaron a través del tiempo, no es fácil obtener mayor información de la relación de estas variables con los PM, por lo tanto se plantea en futuros estudios conocer esta relación en la etapa de transición y posteriormente en el posparto ya que en estos periodos, la variación de la CC y el PV son mayores, con notaria evidencia de un balance energético negativo, tal como ha sido realizado en otros estudios (Soca et al., 2013).
El PV estuvo altamente relacionado con la CC (r=0,795; P<0,01). Datos similares fueron reportados por Villa et al., 2011 durante el periparto en vacas Brahman (r=0,56; P<0,05). La promedios de la CC permite afirmar que las vacas mantuvieron sus reservas de grasa para satisfacer los requerimientos energéticos durante la gestación ya que vacas con promedio de CC superior a 6 son animales con suficientes reservas corporales que les permite una mayor resistencia a condiciones de restricción energética.
Al analizar el comportamiento PV y CC frente al resto de metabolitos, se observó una correlación negativa entre la CC con BUN (r=-0,430; P<0,01) y positiva con la Hb y el Hto (r=0,510; P<0,01; r=0,447; P<0,01 respectivamente). De igual forma, el PV se correlacionó negativamente con el BUN (r=-0,441; P<0,01) y positivamente con la Hb (r=0,397; P<0,01) y el Hto (r=0,258; P<0,05).
Se concluye finalmente que la mayor parte de los metabolitos analizados se encontraban dentro de los límites normales, excepto el Cu y Zn, sin que llegaran a observar desórdenes clínicos asociadas a esta deficiencia. Se reconoce el uso de los PM para identificar los problemas asociados a la alimentación y nutrición y se requieren nuevos estudios en una población más amplia, lo cual permita hacer más eficiente el sistema de cría propio de la Altillanura colombiana.

