











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
La generación de empleo en Colombia en el área rural en gran medida esta soportada en sector agrícola, siendo el cultivo de maíz de gran importancia para la economía campesina. Para el 2019 la producción nacional de maíz blanco y amarillo correspondió a 1.604.792 toneladas sembradas en 386.440 hectáreas. No obstante, la producción nacional no alcanza a suplir la demanda interna del grano; por lo tanto, las importaciones de este cereal suplen el consumo nacional. En 2019 las importaciones de maíz superaron los 5.6 millones de toneladas, las cuales corresponden a maíz de grano blanco y grano amarillo. (FENALCE, 2019). La misma fuente reporta que las importaciones del cereal, en 2019 el 66% provinieron de Estados Unidos. El restante fue aportado por otros países. Es por esta razón que en Colombia la industria de alimentos balanceados destinados para consumo animal principalmente, consume la mayoría del grano proveniente de las importaciones y una menor parte de la producción nacional.
A nivel nacional, los rendimientos por hectárea de maíz tecnificado alcanzaron un promedio de 5.8 toneladas por hectárea en 2019, que comparados con los rendimientos de otros países de zonas tropicales son bastante bajos. No obstante, las producciones de esas regiones permiten inferir que Colombia puede llegar a tener rendimientos por hectárea de más de siete toneladas. Estos rendimientos en grano, en la actualidad ya son obtenidos por productores tecnificados que hacen uso de semillas importadas producidas por empresas extrajeras (observaciones de campo realizadas durante 5 años de trabajo en el Departamento del Valle del Cauca). Es por esta razón que a nivel local en la medida que se logre la identificación de nuevas líneas y la generación de nuevos híbridos con alto potencial de rendimiento, se estima se puede llegar a sustituir parte del grano que ingresa al país a través de las importaciones. Sin embargo, para lograr reducir las importaciones se debe tener en cuenta que el éxito de cualquier programa de mejoramiento genético con énfasis en el desarrollo de líneas endocríadas para la formación de híbridos dependerá de la elección del germoplasma base (De León et al., 2005), del mismo modo que los fitomejoradores requieren de un amplio conocimiento del tipo de acción génica que actúa sobre la expresión de las características de los genotipos en estudio, con el fin de poder implementar los mejores esquemas en los programa de mejoramiento que permitan aprovechar la varianza genética para incrementar o fijar las frecuencias génicas que le sean favorables a las poblaciones.
Es por esto que al estimar los efectos de habilidad combinatoria general (H.C.G) y específica (H.C.E) permite conocer los tipos de acción génica que controlan los diferentes caracteres (Halluer y Miranda, 1988). Las variaciones de habilidad combinatoria general y habilidad combinatoria específica proporcionan una estimación acciones de genes aditivos y no aditivos respectivamente Machikowa et al., (2011). La habilidad combinatoria general (H.C.G) según Sprague y Tatum (1942), define el término como el comportamiento promedio de una línea en combinación híbrida, y Pohelmann (1974), la define como el comportamiento de una determinada línea en una serie de combinaciones híbridas. Por otra parte, la habilidad combinatoria específica (H.C.E), hace referencia a aquellos casos en los que ciertas combinaciones son relativamente mejores o peores de lo que se espera sobre la base del comportamiento medio de las líneas progenitoras involucradas según Sprague y Tatum (1942), De la Rosa et al., (2000), afirman que el conocimiento de la habilidad combinatoria, diversidad genética y heterosis del germoplasma de un programa de mejoramiento es esencial para desarrollar híbridos y variedades.
El análisis de la habilidad combinatoria general (H.C.G), permite identificar adecuadamente los progenitores con capacidad para transmitir sus caracteres deseables a la descendencia, y la habilidad combinatoria específica (H.C.E) posibilita conocer aquellas combinaciones híbridas F1 sobresalientes, originadas de cruzamientos entre variedades, línea o líneas por variedad. Igualmente, este tipo de análisis facilita información sobre el tipo de acción génica que condiciona la expresión de un carácter, lo cual es básico en la escogencia del método de mejoramiento a seguir (Espitia et al., 2006). Mediante la aptitud combinatoria de los progenitores, el mejorador logra mayor eficiencia en su programa de mejoramiento, pues le permite seleccionar progenitores con un comportamiento promedio aceptable en una serie de cruzamientos e identificar combinaciones específicas con un comportamiento superior a lo esperado, con base en el promedio de los progenitores que intervienen en el cruzamiento (De la Cruz et al., 2009).
El presente trabajo se llevó a cabo mediante la formación de todas las combinaciones híbridas posibles bajo la metodología del dialelo entre cinco líneas endocríadas seleccionadas. Los cruzamientos dialélicos son utilizados para estimar los efectos genéticos de las poblaciones en mejoramiento y la información analizada críticamente es valiosa para definir patrones heteróticos, los cuales constituyen una fuente de germoplasma para la generación de líneas élite de gran utilidad en un programa de mejoramiento dinámico (González et al., 1997). El análisis se realizó siguiendo la metodología propuesta por Hallauer y Miranda (1981). Esta metodología permite descomponer las fuentes de variación entre padres, contraste de padres y cruzamientos, análisis de los cruzamientos únicamente y estos a su vez son descompuestos en habilidad combinatoria general (H.C.G) y habilidad combinatoria especifica (H.C.E) con el fin de poder predecir las mejores combinaciones híbridas y que a futuro puedan tener la posibilidad de ser aprovechados desde el punto de vista comercial.
Materiales y métodos
Localización y Establecimiento
El trabajo se realizó en dos etapas, la primera consistió en el aumento de las líneas y la formación de los cruzamientos respectivamente. La segunda etapa estuvo relacionada con la evaluación en campo de las diferentes familias que conformaron el dialélo. La primera etapa llevó a cabo en el primer semestre de 2013 y la segunda en el segundo semestre de 2013 en la finca experimental Algarrobo, ubicada en el corregimiento de Rozo, Municipio de Palmira en el Valle del Cauca - Colombia.
El establecimiento en campo se realizó empleando un diseño de bloques completos al azar con 4 repeticiones. Cada unidad experimental estuvo constituida por cuatro surcos de cinco metros cada uno, espaciados a 0,80 metros entre ellos, se usó una distancia entre plantas de 0,20 metros, dejando una planta por sitio. El total de cada unidad experimental 16 metros cuadrados y 100 plantas en cada una de ellas. La parcela útil estuvo constituida por los dos surcos centrales de cada unidad experimental.
Características Climáticas
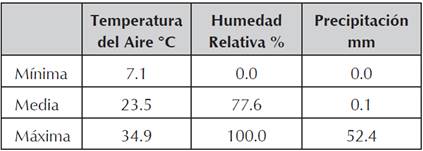
Las características climáticas de la zona donde se realizó la evaluación fueron tomadas de consultas a la base de datos de la Red Meteorológica Automatizada - RMA de Cenicaña (Cenicaña, 2014) estación meteorológica de referencia Rozo. Se tomaron como referentes los datos registrados por la estación desde el año 1993 hasta 2013.
Tabla 1 Datos climatológicos zona de influencia estación meteorológica Rozo. Red Meteorológica Automatizada - RMA del Centro de Investigación de la Caña de Azúcar de Colombia (Cenicaña, 2014).
Manejo Agronómico
La preparación del terreno se realizó con maquinaria agrícola y el establecimiento del ensayo se realizó forma manual, la aplicación de fertilizantes se llevó a cabo de forma fraccionada en dos aplicaciones, la primera transcurridos los primeros 15 días después de germinado y la segunda un mes después de la primera aplicación. La cantidad de fertilizante aplicado fue el indicado según los requerimientos de la zona para el cultivo (García Montealegre, 2008) y el análisis químico de suelo. El requerimiento hídrico fue suplido mediante la aplicación de riego por gravedad según la necesidad del cultivo, de igual modo los controles fitosanitarios para evitar pérdida de población debido a insectos propios del cultivo. Se realizó control y manejo de malezas durante los dos primeros meses de edad, hasta que las calles cerraron y evitaron el crecimiento de nuevas arvenses.
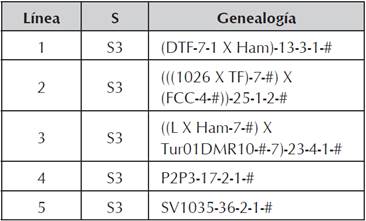
Material Genético
Los genotipos evaluados, estuvieron conformados por líneas en estado S3 de endocría y sus respectivos híbridos sencillos formados mediante la metodología de dialelo. Se evaluaron cinco (5) líneas de grano amarillo y los híbridos directos, conformados por estas. En total se evaluaron 15 genotipos (5 líneas y 10 híbridos).
Variables Evaluadas
DÍAS A FLORACIÓN MASCULINA Y FEMENINA (DFM - DFF). Se tomaron diez plantas al azar y se midió los días transcurridos desde la emergencia en campo hasta la aparición de las inflorescencias masculina y femenina. Este dato se tomó cuando el 50% de las plantas seleccionadas llegaron al periodo de antesis. ALTURA DE PLANTA (Alt. Planta). Esta variable se tomó a las diez plantas seleccionadas, se midió en metros, desde el suelo hasta la punta de la inflorescencia masculina. ALTURA DE MAZORCA (Alt. Mzca). Se registró en las diez plantas tomadas al azar en cada unidad experimental, se midió desde la base de la planta al nudo con la mazorca más alta. La medida se tomó dos semanas antes de la cosecha. La unidad de medida fue metros. LARGO DE MAZORCA (Larg. Mzca). Se tomó el dato de las diez plantas tomadas al azar en cada unidad experimental. La medida se obtuvo al momento de la cosecha. La unidad de medida fue centímetros. DIÁMETROS (Diam. Mzca y Diam. Tusa). Se tomaron las mazorcas de las diez plantas seleccionadas y se midieron sus diámetros antes de desgranar y después de desgranadas. La medida empleada fue centímetros. EVALUACIÓN POR ENFERMEDADES (Fus. Mzca=Fusarium en Mazorca, Phys=Physoderma maydis). Se evaluó la presencia de enfermedades mediante escalas de evaluación establecidas por CIMMYT (CIMMYT, 2004). NÚMERO DE PLANTAS COSECHADAS. Se contó el número de plantas cosechadas por parcela útil. NÚMERO DE MAZORCAS COSECHADAS. Se contó el número de mazorcas cosechadas de cada parcela útil en cada parcela experimental. PUDRICIÓN DE MAZORCAS. Con contó el número de mazorcas que presentaban pudriciones y se tuvo como referente las escalas de evaluación establecidas por CIMMYT (CIMMYT, 2004). NÚMERO DE HILERAS POR MAZORCA (Num. Hill). Se contó el número de hileras con granos presentes en cada mazorca cosechada de las diez plantas seleccionadas inicialmente. NÚMERO DE GRANOS POR HILERA (Grn. Hill). De las mismas diez plantas tomadas al azar se realizó el conteo de los granos de las mazorcas para sacar el promedio general. PESO DE CAMPO. En cada parcela y sin desgranar, se tomó el peso de las mazorcas de cada parcela útil. RELACIÓN GRANO/TUSA (Grn/Ts). Las mazorcas por parcela experimental útil se desgranaron y pesaron separadamente grano y tusa para determinar su relación. PESO DE GRANO. Se tomó el peso del grano de las mazorcas pesadas anteriormente. PESO DE 1000 GRANOS (P. 1000 Grn). Mediante el uso de una balanza de precisión, se cuantificó el peso promedio de 1000 semillas de los genotipos evaluados. RENDIMIENTO POR PARCELA (Rend). Estuvo determinado por el peso de los granos provenientes de cada parcela útil, Los pesos fueron corregidos por población y uniformizados al 15% de humedad y convertidos a kilogramos por hectárea mediante al formula:
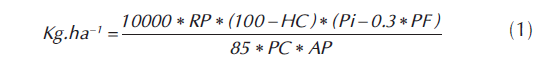
Análisis de Datos
Los datos generados se analizaron usando el programa estadístico SAS (SAS Institute Inc., 1997) y GENES (Cruz, 1998), siguiendo las recomendaciones de Vallejo et al, (2005). En el análisis o comparación de medias, sólo se usaron aquellas variables en las cuales el análisis de varianza mostró diferencias estadísticamente significativas. Para la comparación de medias se usó el estadístico de LSD. (Salazar, 2007).
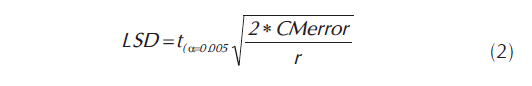
Dónde:
LSD = Diferencia Mínima Significativa
CMerror = Cuadrado Medio del Error
t(α=0.05) = valor de probabilidad de t, a un nivel de significancia del 5%.
r = número de repeticiones o número de datos que dieron origen a la media
El análisis genético se realizó usando la metodología propuesta por Hallauer y Miranda (1981), la cual es una combinación de las ventajas de las metodologías de Gardner y Eberhart (1966) con las metodologías de Griffing (1956), donde la suma de cuadrados de las entradas se divide en tres componentes, uno debido a padres, otro a cruzas y el tercero al contraste de padres vs cruzamientos, éste último componente es similar a la heterosis media del método de Gardner y Eberhart (1966). La suma de cuadrados debida a la variación de las cruzas se analizó de manera similar a la metodología 4 de Griffing (1956), en donde sólo se usó para el análisis las cruzas F1’s sin tener en cuenta los padres. El modelo estadístico para el análisis de varianza fue:
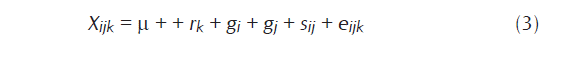
Donde:
X ij = Es el comportamiento de cualquier observación.
µ = Es la media general.
r k = Es el efecto de la replicación k.
S ij = Es el efecto de H.C.E.
e ijk = Es el error experimental para las Xijk observaciones, con K=1,2, … r repeticiones
i = j 1, 2, … n padres
La heterosis y heterobeltiosis, expresada en porcentaje se cuantificó a través del software GENES (Cruz, 1998), sin embargo, el procedimiento para su cálculo se puede definir como:
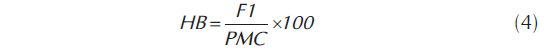
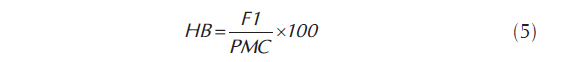
Resultados
Las diferencias estadísticamente significativas entre genotipos radican en el contraste que hay entre los distintos genotipos homocigotos de los progenitores que corresponden a líneas endogámicas (S3) y los genotipos híbridos generados a través de los cruzamientos. Estas diferencias de expresiones fenotípicas, también pueden ser explicadas por la heterosis que se genera en los híbridos.
Se encontraron diferencias estadísticamente significativas (Tabla 3), entre genotipos parentales para algunos de los caracteres evaluados (P<0,05). Sin embargo, para la variable rendimiento (Rend) no se hallaron diferencias estadísticas significativas, atribuido posiblemente al nivel de endocría de las líneas homocigotas y su depresión en el carácter medido Hallauer y Miranda (1988), García et al., (2004). No obstante, caracteres relacionados a la fenología de la planta como días a floración femenina (DFF) y días a floración masculina (DFM), sus cuadrados medios si presentaron diferencias estadísticas significativas (P<0,05). De igual modo, para los componentes relacionados con el rendimiento/planta como son; granos por hilera (Grn.Hill), número de hileras (Num.Hill) y peso de 1000 granos (P.1000Grn), se encontró que existen diferencias entre los padres (Tabla 4). Para estos mismos componentes del rendimiento, el análisis en el contraste de padres versus cruzamientos (P vs C) presentan diferencias estadísticas (P<0,05) y que da respuesta a la manifestación de la heterosis como expresión del vigor hibrido de los cruzamientos entre progenitores comparado con los genotipos parentales y que son líneas avanzadas en el proceso de endocría.
Tabla 3 Cuadrados medios y significancia estadística de variables de líneas de maíz amarillo y sus respectivos cruzamientos
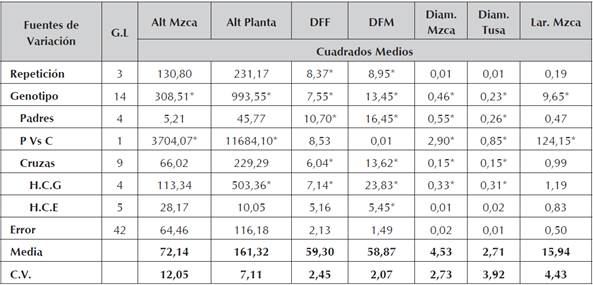
* Significativo al nivel de probabilidad de 0.05.
Tabla 4 Cuadrados medios y significancia estadística de variables del componente rendimiento y enfermedades de líneas de maíz amarillo y sus respectivos cruzamientos
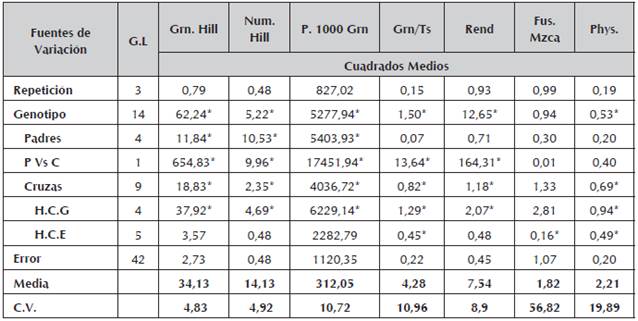
*Significativo al nivel de probabilidad de 0.05.
Resultados de Habilidad Combinatoria General (H.C.G)
En el análisis de los efectos aditivos, sólo unas pocas de las variables analizadas mostraron efecto significativo (p<0.05) inherente a la habilidad combinatoria general (H.C.G.). Para los caracteres en evaluación, altura de mazorca (Alt.Mzca) y altura de planta (Alt.Planta) el progenitor número uno (1), mostró que es capaz de transferir la característica a su descendencia y los efectos fueron estadísticamente significativos a un nivel de probabilidad de p<0.05 (Tabla 5).
Tabla 5 Efectos de Habilidad Combinatoria General (H.C.G) en las cinco líneas de endospermo amarillo evaluadas a un nivel de significancia del 0.05.
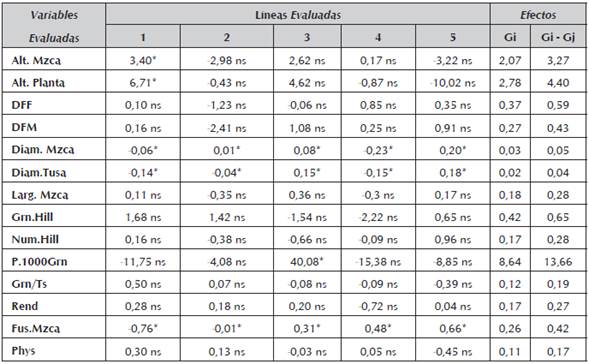
* Significativo al nivel de probabilidad de 0.05, ns= no significativo
Según Montenegro Torres et al., (2002), los valores de H.C.G ayudan a identificar los genotipos que en promedio combinan mejor con otros. Sin embargo los valores de los efectos de habilidad combinatoria general (H.C.G) (Tabla 5), se puede ver que la mayoría de los valores fueron no significativos, sólo se presentaron valores significativos para el carácter diámetro de mazorca (Diam.Mzca), diámetro de tusa (Diam,Tusa) y pudrición de la mazorca por fusarium sp. No obstante, la no significancia de los valores para los efectos de habilidad combinatoria general y los valores de varianza bajos para estos efectos puede ser un indicio de que las líneas evaluadas están emparentadas genéticamente y por lo tanto su distancia y variabilidad genética es mínima.
Bajo el supuesto de emparentamiento de las líneas al hacerse un cruzamiento entre ellas se logra un efecto de endogamia y es posiblemente la razón por la cual los valores mostrados son no significativos. Otra posible causa de los valores bajos para los efectos de habilidad combinatoria general (H.C.G), puede ser el bajo nivel de endocría de las líneas o de homocigocis para los caracteres evaluados que no permitió el re-establecimiento de la heterosis en las combinaciones híbridas resultantes Hallauer y Miranda (1988).
Resultados de Habilidad Combinatoria Específica (H.C.E)
Mediante los efectos de habilidad combinatoria específica se pueden determinar dos grupos heteróticos de líneas “A” y “B” que forman el patrón heterótico de las líneas bajo análisis. En este trabajo, se encontró que la mayoría de los efectos de habilidad combinatoria específica (H.C.E) fueron no significativos estadísticamente a un nivel de probabilidad (p<0.05).
La escasa manifestación de valores significativos en los valores para H.C.E (Tabla 6), se puede corroborar con los obtenidos en la expresión de la heterosis, y aunque arrojaron algunos datos con valores estadísticamente significativos no son generalizados ni representativos en la expresión del carácter en estudio. Es por ello que, la escasa significancia de efectos de habilidad combinatoria específica indica la escasa capitalización de los efectos genéticos no aditivos o dominantes en el proceso de selección genealógica y el beneficio en la heterosis de las combinaciones híbridas resultantes.
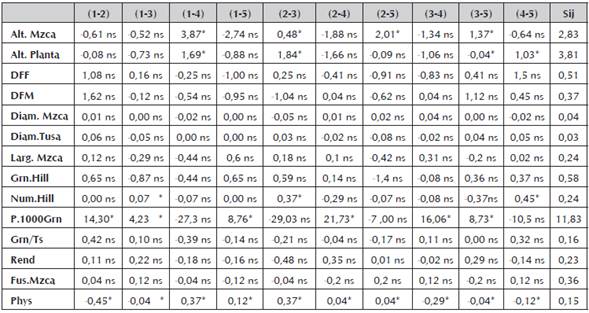
Resultados de la Heterosis y Heterobeltiosis
La heterosis se origina en la hibridación, pero no es un fenómeno universal ni fácilmente observable. Se presenta cuando se cruzan padres genéticamente diferentes. El efecto principal de la heterosis, en general, está relacionado con un aumento significativo de la productividad, y un gran número de caracteres agronómicamente importantes, también han sido mejorados a través de la heterosis (Vallejo et al., 2010).
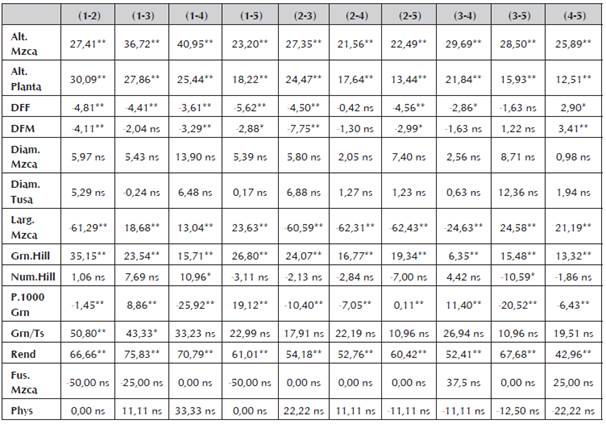
Aunque en los cruzamientos producto de las líneas bajo estudio en este trabajo presentaron para algunos caracteres heterosis positiva y estadísticamente significativa (Tabla 7), sus valores no son tan altos como se esperaría para una muy buena combinación hibrida y que a futuro pueda ser de interés comercial. Estos resultados son congruentes con los encontrados en los valores para los efectos de habilidad combinatoria especifica (H.C.E), dónde los valores fueron estadísticamente no diferentes de cero, en su gran mayoría. Este comportamiento en los valores obtenidos es atribuido posiblemente a la presencia de líneas emparentadas, genéticamente similares o a su bajo nivel de endocría lo que concuerda con enunciado por Vallejo et al., (2002).
Tabla 7 Heterosis en términos de porcentaje (%) para cada una de las variables respuesta evaluadas en cada uno de los cruzamientos de endospermo amarillo.
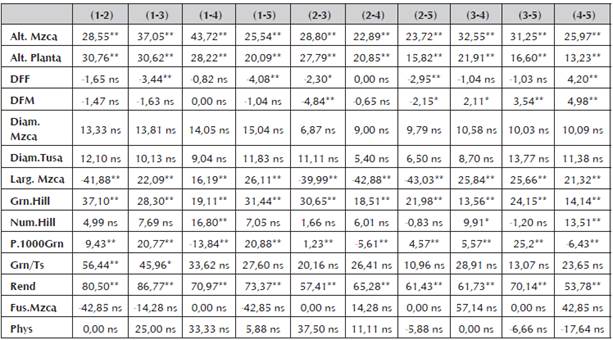
“*” y “**” Significativo al nivel de probabilidad de 0.05 y 0.01 respectivamente, “ns” No significativo.
El carácter altura de planta (Alt.Planta) y altura de mazorca (Alt.Mzca), en todas las combinaciones híbridas evaluadas, presentó heterosis altamente significativa, sin embargo, los valores no superan el 50%.
De igual modo, para los demás valores evaluados se presenta heterosis de tipo positivo o trans-heterosis y valores de tipo negativo o cis-heterosis. Y, aunque se presentó heterosis en los caracteres evaluados, sea de tipo positivo o tipo negativo y con significancia estadística, todos los valores se encuentran por debajo del 50% en su expresión, a excepción del carácter rendimiento que obtuvo valores muy superiores y estadísticamente significativos (p<0.05).
La heterosis basada en el promedio de los progenitores tiene su importancia desde el punto de vista biológico y genético en razón a que considera el aporte genético de los dos progenitores. En el mismo sentido que la heterosis, la heterobeltiosis se calculó respecto al progenitor superior presentando datos altamente significativos y presentan similitud a los encontrados respecto a la heterosis. Para las variables altura de mazorca y altura de planta, se presentan los valores más altos respecto a la expresión en porcentaje de la heterobeltiosis (Tabla 8). Los valores para el carácter rendimiento en las cruzas evaluadas poseen una alta expresión respecto a sus progenitores, dando como resultado valores de heterobeltiosis altos y altamente significativos.
Discusión
El valor de los cuadrados medios de la habilidad combinatoria general (H.C.G) en todas las variables analizadas fue mayor que los de habilidad combinatoria específica (H.C.E), lo que indica que la variación genética del tipo aditivo está gobernando en mayor proporción la repuesta de las variables analizadas (Salazar, 2007). La suma de cuadrados para habilidad combinatoria general (H.C.G), la cual está relacionada con el componente genético aditivo presentó significancia en las variables evaluadas exceptuando altura de mazorca (Alt. Mzca) y largo de la misma (Larg.Mzca) (Tabla 5). En este mismo componente se presentaron diferencias significativas en la variable días a floración femenina, rendimiento y altura de la planta. Estos resultados son congruentes con los reportados por San Vicente et al., (2001), quienes analizaron la aptitud combinatoria y patrones heteróticos entre poblaciones tropicales de maíz de endospermo amarillo, encontrando que los efectos aditivos contribuyen a la expresión de los caracteres días a floración femenina, la altura de la planta y el rendimiento.
No obstante, para la respuesta al rendimiento, los resultados son disimiles a los encontrados por De la Rosa et al., (2000), quienes en su estudio hallaron que la contribución a la varianza del rendimiento atribuible a los cruzamientos está constituida por un mayor porcentaje a los efectos no aditivos (H.C.E) que a los efectos aditivos (H.C.G). Para este estudio en particular se encontró que para la característica expresión del rendimiento se observaron mayores efectos del tipo aditivo (H.C.G), con una participación del 77% del total del componente y una participación del 23% debido a los efectos de tipo no aditivo o dominante. Estos resultados se deben a que en este carácter no se presentó como se esperaba un alto beneficio y manifestación de la heterosis. Estos resultados son similares a los hallados por Ortiz et al., (1993), quienes atribuyen la importancia de partición de la variación entre las cruzas y que la partición revela que los cuadrados medios de la habilidad combinatoria general (H.C.G) y específica (H.C.E) son significativamente diferentes. En este contexto, se encontró que el valor de los cuadrados medios de la H.C.G fue más del doble que los de la H.C.E, indicando que la variación genética de tipo aditivo está gobernando la repuesta para el carácter rendimiento
Para el caso de los efectos genéticos no aditivos o los relacionados con la habilidad combinatoria especifica (H.C.E) (Tabla 6), se encontró que el cuadrado medio de este componente es significativo para la variable días de floración masculina (DFM), pudrición por fusarium en la mazorca (Fus.Mzca) y enfermedades de campo como Physoderma sp. (Phys). El componente aditivo para la variable (Fus.Mzca), tiene una participación de 93%, el 7% restante es atribuible al componente no aditivo, este es un resultado favorable ya que permite hacer inferencia que no hay participación de la heterosis o que su efecto es mínimo en la expresión de la pudrición de la mazorca atribuida a Fusarium sp. Estos resultados, aunque no son iguales, se asemejan a los reportados por Reid, Zhu et al., (2009), quienes encontraron que la pudrición por Fusarium sp., estaba asociada a genes de tipo aditivo con una participación de 66% y un porcentaje menor (34%, a los de tipo dominante. Para la expresión de la enfermedad Physoderma maydis (Phys), al igual que para (Fus.Mzca) los mayores efectos son de tipo aditivo, 61% de la varianza es atribuible los efectos de tipo aditivo y el restante 39% de participación pertenece a los efectos de tipo dominante.
Los resultados obtenidos para habilidad combinatoria especifica (H.C.E), son congruentes con lo hallado en los estimativos para habilidad combinatoria general (H.C.G), donde la mayoría de los valores fueron no significativos solo en casos específicos se mostró significancia. González et al., (1997), afirman que cruzas con efectos de habilidad combinatoria específica (H.C.E) negativos significa que las líneas pertenecen a un mismo grupo heterótico y cruzas con efectos de habilidad combinatoria específica (H.C.E) positivos significa que pertenecen a grupos heteróticos opuestos. Por lo tanto, teniendo en cuenta los resultados obtenidos por este autor, los datos registrados en la tabla 6., se puede deducir que no existe patrón heterótico entre las líneas evaluadas. Por lo tanto, la manifestación de valores positivos o significativos en los efectos para de habilidad combinatoria específica (H.C.E) son escasos. Es decir, que probablemente las líneas evaluadas pertenecen al mismo grupo heterótico.
La relación entre los valores de heterosis (Tabla 8) y habilidad combinatoria específica (H.C.E) para rendimiento, concuerdan con los resultados expuestos por De la Cruz et al., (2010). Estos autores mencionan que los mayores valores de heterosis, corresponden a las mayores H.C.E. Adicionalmente, afirman que la estimación de la heterosis de las cruzas puede estar favorecida por los bajos rendimientos de ambos progenitores, cosa que no se presenta tan clara en el presente estudio, ya que los progenitores evaluados presentaron medias de rendimiento muy similares. Se infiere que el grado de expresión de la heterosis está relacionado con el nivel de endocría de las líneas parentales, menor nivel de endocría generara menor expresión en el porcentaje de la heterosis para el carácter rendimiento.
Los resultados obtenidos en este trabajo, se convierten en una herramienta para avanzar en los procesos de mejoramiento genético de las poblaciones de dónde fueron obtenidas las líneas evaluadas. No obstante, se debe articular con estudios de diversidad genética a través de métodos moleculares con el fin de poder conocer de manera más profunda las distancias genéticas presentes entre los distintos grupos de poblaciones para la obtención de líneas destinadas a programas de hibridación.

