










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkVitae
Print version ISSN 0121-4004
Vitae vol.17 no.2 Medellín May/Aug. 2010
PRODUCTOS NATURALES
ESTUDIO DE LA COMPOSICIÓN QUÍMICA Y ACTIVIDAD ANTIFOULING DEL EXTRACTO DE LA ESPONJA MARINA Cliona delitrix
STUDY OF THE CHEMICAL COMPOSITION AND ANTIFOULING ACTIVITY OF THE MARINE SPONGE Cliona delitrix EXTRACT
Leonardo CASTELLANOS H.1; Humberto MAYORGA W. 1; Carmenza DUQUE B.1*
1Grupo de estudio y aprovechamiento de productos naturales marinos y frutas de Colombia. Departamento de Química. Facultad de Ciencias. Universidad Nacional de Colombia. A.A. 14490. Bogotá, Colombia.
RESUMEN
El extracto crudo y la fracción orgánica de la esponja marina Cliona delitrix, recolectada en San Andrés Islas (Caribe Colombiano), muestra actividad antifouling en ensayos de campo. La fracción orgánica es separada por cromatografía en columna logrando obtener fracciones enriquecidas en glicéridos, glicolípidos, fosfolípidos, ácidos grasos libres, ésteres metílicos y monohidroxiesteroles. Cada una de las fracciones es identificada de manera preliminar por TLC y técnicas de dereplicación (RMN). Los ácidos grasos obtenidos de la hidrólisis de cada una de las fracciones de glicéridos, glicolípidos y fosfolípidos son transformados en sus correspondientes ésteres metílicos, los cuales son analizados por CGAREM; luego se convierten en sus derivados pirrolidínicos y también se analizan por CG-EM. El estudio cuidadoso de los EM, tanto del éster metílico como de las pirrolidina, y los valores de ECL permiten identificar 89 ácidos grasos diferentes. Entre éstos se destacan: el ácido 7,10,12,14-icosatetraenoico, que se reporta por primera vez, y los ácidos 5,9-hexacosadienoico y hexadecanoico, por ser los más abundantes en todas las fracciones. Adicionalmente se identifican, mediante análisis de CG-EM y 1H-RMN, 19 monohidroxiesteroles. Se destacan el colestanol y el clionasterol, por ser los más abundantes, y también es de notar la presencia de estanoles (cerca del 30%) que no se habían informado antes para este género.
Palabras Clave:Cliona delitrix, lípidos, ácidos grasos, esteroles, antifouling.
ABSTRACTIn a field test for antifouling activity the raw extract and the organic fraction obtained from the marine sponge Cliona delitrix, collected at the San Andrés Islas (Colombia, Caribbean Sea), showed activity. The organic fraction was separated by column chromatography to obtain enriched fractions of glycerides, glycolipids, phospholipids, free fatty acids, fatty acids methyl esters (FAMEs) and monohydroxysterols identified by TLC and dereplication techniques (NMR). The glyceride, glycolipid, and phospholipid fractions were hydrolyzed, and the fatty acids methyl esters obtained, together with the initial fatty acids fraction were converted into their methyl esters and analyzed by HRGC-MS. In order to locate unsaturations and alkyl branches in fatty acids, their methyl esters derivates were transformed to the corresponding pyrrolidides and subsequently analyzed by HRGC-MS. The identification of the fatty acids was carried out using their ECL value as methyl esters, and particularly the study of their pyrrolidide spectra was used to locate unsaturations and alkyl branch positions. The analysis allowed the identification of 89 fatty acids. The main acids include the new 7,10,12,14-icosatetraenoic acid, as well as 5,9-hexacosadienoic acid and hexadecanoic acid. Additionally, 19 monohydroxysterols were identified by HRGC-MS and NMR 1H. Cholestanol and clionasterol were the most abundant compounds and the stanols presence (near to 30%) was not previously reported for this genus.
Keywords: Cliona delitrix, lipids, fatty acids, sterols, antifouling.
INTRODUCCIÓN
Los organismos marinos han mostrado ser una fuente de compuestos químicos de estructuras muy diversa, y en muchos casos con potente actividad biológica. Es notoria la enorme diversidad química de las esponjas, los corales, y más recientemente los microorganismos. Esta diversidad química puede obedecer a la interacción entre ellos; así, en invertebrados, la producción de compuestos químicos puede tener propósitos diversos, tales como evitar la depredación, capturar presas, combatir microorganismos, o detener el fouling, entre otros (1-2). El fouling marino se entiende como un proceso de deposición de moléculas seguido del asentamiento y desarrollo de organismos vivos sobre las superficies sumergidas. Este fenómeno obedece a la necesidad que tienen muchos organismos marinos de asentarse para su desarrollo, y hace parte de la competencia natural por colonizar y defender el espacio adquirido en el bentos, recurso que es muy limitado en el mar (3-4); así, los organismos bentónicos pueden producir sustancias químicas que los ayuden a controlar ese proceso (compuestos antifouling) (5).
Si bien el fouling es un proceso natural, y en muchos casos deseado, puede ocasionar problemas como corrosión, pérdidas económicas por mantenimiento de embarcaciones, sobrecostos en transporte marítimo etc. Para combatir este fenómeno el hombre ha desarrollado múltiples estrategias, entre ellas el empleo de recubrimientos poliméricos, pinturas con pigmentos antifouling activos, y pinturas con biocidas embebidos o enlazados a la matriz de las mismas (6). Algunos de estos biocidas han sido prohibidos por sus efectos desfavorables en muchos organismos marinos ajenos al proceso del fouling, lo que ha obligado a buscar nuevas alternativas eco amigables (biodegradables o con mayor armonía medio ambiental) para reemplazar los recubrimientos antifouling existentes (7). Una posibilidad es el uso en pinturas antifouling de las defensas químicas naturales presentes en los organismos marinos, por ejemplo, el alga roja Delisea pulchra produce algunas 2(5H)-furanonas que inhiben el asentamiento y colonización de micro y macroorganismos del fouling en su superficie. Las 3-alquilpiridinas, y sus derivados poliméricos, presentes en la esponja Reniera sarai, también previenen el biofouling microbiológico, y son poco tóxicas para el medio ambiente (4). Otros compuestos químicos con actividad antifouling comprobada son los ácidos grasos de algunos corales blandos del género Dendronephthya (8).
Por otra parte,se sabe que también hay moléculas químicas que actúan como inductores del asentamiento de larvas de los organismos que hacen parte del fouling. Algunas de las más representativas son la serotonina, que induce el asentamiento y metamorfosis de Balanus amphitrite; la histamina, involucrada en la percepción de luz/oscuridad de estos balanos, que de esta manera influye en su asentamiento (9). También se ha visto que algunos derivados bromados de la serotonina, como la barrettina y 8,9-dihidrobarrettina, aislados de la esponja Geodia barretti, compiten por los receptores de la serotonina en los cirripedios, actuando así como inhibidores del asentamiento de las larvas de cirripedios más que como inductores (3).
La esponja excavadora e incrustante Cliona delitrix Pang, 1973 (Orden: Hadromerida; familia: Clionaidae) mantiene su superficie limpia de epibiontes y es capaz de colonizar y defender su sustrato (10-11), por lo que puede ser una fuente interesante de compuestos con actividad antifouling. No obstante, sólo hay un reporte sobre la química de esta especie, recolectada en Bahía, Brasil (12). En cuanto al género, se ha encontrado que es fuente de alcaloides peptídicos (13), de aminoácidos (11), de esteroles modificados (14) y de lectinas (15). También se estudiaron los lípidos de otras especies de este género, como los esteroles de Cliona caribbaea (16), esteroles y ácidos grasos de C. celata (17-18), esteroles de C. viridis (19), fosfolípidos de C. aprica (20), y recientemente ácidos grasos de C. tenuis (21). Estos lípidos aislados de fuentes marinas han mostrado actividad citotóxica, fungicida, inmunoestimulatoria, de inhibición enzimática (2, 22) y algunos ácidos grasos tienen posibilidades de aplicación como agentes antifouling (8, 23-25), por lo que actualmente se hacen esfuerzos tendientes a obtenerlos mediante síntesis química (26-27).
Por todo lo anterior, y con el fin de contribuir al conocimiento de los lípidos del género Cliona, este estudio expone los resultados sobre la determinación de actividad antifouling del extracto crudo de Cliona delitrix y la caracterización química de la fracción lipídica responsable de dicha actividad.
MATERIALES Y MÉTODOS
Recolección de muestras
Individuos de la esponja marina C. delitrix fueron recolectados, con ayuda de cincel y martillo, en buceo autónomo en la localidad de Wildlife, Isla de San Andrés (Colombia), a una profundidad entre 6 y 10 m, en los meses de mayo de 2003 y septiembre de 2004. Los organismos congelados (-30°C) fueron enviados a Bogotá para su estudio. Ejemplares de dicho material fueron identificados por el profesor Sven Zea S., del Departamento de Biología de la Universidad Nacional de Colombia, y un espécimen fue depositado en la colección de INVEMAR con el código INV-POR 629.
Equipos de análisis y materiales
Para la CCD se emplearon cromatoplacas de gel de sílica HF254 Merck, observadas con lámpara ultravioleta a 254 y a 366 nm y reveladas con solución de ácido fosfomolíbdico (al 20% en etanol); además, para la detección de glicolípidos se utilizó H2SO4-α-naftol (al 5% en metanol), y para los fosfolípidos, azul de molibdeno (0,5% en H2SO4 diluido). Los espectros de RMN fueron registrados en un equipo Brucker-400 (13C a 100 MHz), empleando CDCl3 y CD3OD como solventes y TMS como referencia interna. Los análisis por CLAE se llevaron a cabo en un cromatógrafo Merck-Hitachi L-6000A, equipado con un detector de índice de refracción Merck LaChrom L-7490. Los análisis por cromatografía de gases de alta resolución-espectrometría de masas CGAR-EM se efectuaron en un cromatógrafo Shimadzu GC-17A acoplado a un espectrómetro de masas QP5050A (IE a 70eV), empleando una columna capilar de sílica fundida DB-1 de 25 m x 0,25 mm d. i., 0,25 μm de espesor de película y como gas de arrastre helio a 1 mL/min.
Extracción y fraccionamiento de los lípidos
La esponja marina C. delitrix congelada (1,5 Kg) se partió en finos trozos con cincel y martillo, eliminando el soporte calcáreo no excavado y otros macroorganismos. Los trozos de esponja se extrajeron primero con metanol, relación muestra:solvente 2:1, y luego con diclorometano. Los extractos fueron filtrados, concentrados en rotaevaporador y reunidos como un extracto crudo total (ECT). El ECT fue sometido a partición entre CH2Cl2 y H2O (1:1). La fase acuosa obtenida fue extraída con butanol, lo que condujo a una subfracción acuosa WW (81,6% del ECT) y subfracción butanólica WB (10,4%), siguiendo la metodología de Kupchan (28). La fase de diclorometano no polar NP (8% del ECT), posteriormente se separó entre metanol acuoso (90%) y hexano, obteniendo la subfracción metanólica FM (1,7% de NP) y la subfracción de hexano FH (5,8% de NP). La subfracción FH se separó por cromatografía en columna (CC) sobre sílica gel (0,040-0,063 mm, Merck) empleando mezclas de hexano y acetato de etilo para su elución, obteniendo fracciones enriquecidas en esteres metílicos (11% de FH), en acilglicéridos (47% de FH), ácidos libres (3.5% de FH) y monohidroxiesteroles (30% de FH). Estas fracciones fueron localizadas en el ef luente de CC por CCD comparativa usando patrones de cada tipo de compuestos, y caracterizadas de manera preliminar por RMN 1H y 13C. Las subfracciones de fosfolípidos y glicolípidos fueron obtenidas de la fracción NP por separación cromatográfica al vacío sobre sílica gel (0,040-0,063 mm, Merck), tratada previamente con hidróxido de amonio, seguida de secado a 120ºC, como lo indica la metodología usada originalmente por Privett (29). En la fracción eluida con acetona se detectó la presencia de glicolípidos (10% de NP) por análisis mediante cromatografía en capa fina, según el color violeta del revelado frente a una solución de H2SO4-α-naftol, y por el análisis de espectros de RMN 1H y 13C, debido a la presencia de señales características de residuos de azúcar. En la fracción eluida con metanol (7% de NP) se detectaron fosfolípidos, según el color azul de revelado con una solución de azul de molibdeno, y por comparación frente a patrones de fosfolípidos en CCD, además del análisis de los espectros de RMN 1H y 13C de esta fracción.
Aislamiento de compuestos polares
La subfracción butanólica WB (5 g) se pasó por una columna cromatográfica empacada con 750 g de resina XAD-4, lavando con 2 L de agua 1 mL/min. La fracción orgánica retenida se eluyó c on 1,5 L de metanol y 3 L de metanol:diclorometano 8:1. Esta fracción se separó luego con Sephadex LH-20 (3.0 cm x 85 cm), eluyendo con metanol a 1 mL/min. Mediante análisis por CCD sobre gel de sílice con la fase móvil butanol:ácido acético:agua (12:3:5), y por RMN 1H, RMN 13C, se reunieron 12 fracciones. La fracción 2 fue purificada por CLAE (30), en RP-18 (Columna Scharlau Science, Nucleosil, de 300 x 8 mm, 10 μm), usando agua:metanol (97:3) con TFA al 0,05% a 1 mL/min, y detector de índice de refracción, obteniendo así la serotonina (90% de WB). Su identificación se hizo por RMN 13C, y comparación con datos de literatura (31-32). Adicionalmente, una porción de la subfracción WW (62 g) se pasó por una columna de XAD-4 en agua, la fracción adsorbida fue luego eluida con acetona, metanol y diclorometano. Este residuo se purificó por cromatografía en Sephadex LH-20 con H2O:MeOH (9:1), obteniendo el compuesto 1H-imidazol-2(3H)-imina (169 mg) puro, cuya identificación se hizo por RMN 1H y 13C, y comparación con datos de la literatura (33).
Preparación de los ésteres metílicos de ácidos grasos y sus derivados N-acilpirrolididas
Las fracciones de ácidos libres, acilglicéridos, glicolípidos y fosfolípidos fueron convertidas a sus respectivos ésteres metílicos por reacción con hidróxido de potasio metanólico (10%) y luego con BF3-MeOH (14%). Los ésteres metílicos fueron extraídos con hexano, y el extracto orgánico, luego de ser secado sobre NaSO3 anhidro, fue concentrado. Para obtener los derivados pirrolidida, a 1 a 2 mg de los ésteres metílicos disueltos en pirrolidina (0,1 mL) se les adicionó ácido acético (0,01 mL) y la mezcla fue calentada a 110°C durante dos horas; una vez llevada a temperatura ambiente, la mezcla fue extraída con diclorometano, la fase orgánica se lavó con HCl 1M y tres veces más con agua en porciones de 1mL. Finalmente fue secada sobre NaSO4 anhidro y purificada por cromatografía en columna sobre sílica gel (34).
Análisis por CGAR-EM de los ésteres metílicos y pirrolididas de ácidos grasos
Los ésteres metílicos (muestra de 1 mg/mL) fueron analizados por CGAR-EM con el siguiente programa de temperatura: 5 min isotérmica a 100°C, incremento a 4°C/min hasta 300°C durante 40 min. La temperatura del inyector y detector fue mantenida a 300°C. Para las pirrolididas (muestra de 1 mg/mL) se usó un programa de temperatura desde 200°C durante 5 min, calentando a 4°C/min hasta 300°C durante 55 minutos. En la identificación de los ácidos grasos y sus derivados se analizaron los índices de retención ECL (longitud equivalente de cadena, por sus siglas en inglés) y los datos del espectro de masas de cada compuesto. La cuantificación se llevó a cabo por el método de normalización de áreas en CGAR.
A continuación se presenta el índice de retención ECL y los datos del espectro de masas del ácido graso nuevo.
7,10,12,14-icosatetraenoato de metilo (44): ECL 19.46; EM, m/z 318 (M+, 1%), 289 (1%), 267 (1%), 249(1%), 235 (1%), 217 (1%), 191 (2%), 161 (6%), 147 (4%), 133 (7%), 119 (10%), 105 (12%), 93 (40%), 91 (50%), 79 (100%), 67 (80%).
Pirrolidida del ácido 7,10,12,14-icosatetraenoico (44): EM, m/z 357 (M+, 5%), 342 (1%) 328 (4%), 314 (3%), 300 (1%), 286 (1%), 273 (1%), 260 (2%), 247 (1%), 234 (2%), 221 (1%), 208 (8%), 194 (2%), 181 (1%), 168 (12%), 154 (6%), 140 (12%), 126 (75%), 113 (100%), 98 (50%).
Análisis de monohidroxiesteroles
La fracción esterólica, obtenida a partir de la subfracción FH, se separó por CLAE semipreparativa y cada fracción obtenida se analizó por CGAR-EM. La CLAE se realizó con una columna ShimpackCLC-ODS (150 mm x 6 mm, 5 μm), con metanol como fase móvil (1,0 mL/min), y la CGAR-EM se hizo en la columna DB-1 mencionada antes, a una temperatura de 300ºC y un flujo de 1mL/min. Como criterio de identificación se usaron los espectros de masas y el valor del tiempo de retención relativo al colesterol. Para algunas características estructurales, tales como la estereoquímica en el C-24, se empleó la RMN 1H (400 MHz). La cuantificación de los esteroles se llevó a cabo por el método de normalización de áreas tanto en CGAR como en CLAE (35).
Ensayo en campo de actividad antifouling
Para este ensayo se utilizaron geles de PhytagelTM siguiendo la metodología usada por Henrikson and Pawlik (36), y empleando cajas de petri (90 mm de diámetro x 10 mm de altura) para verter los geles con los extractos de prueba, ECT (63 mg/mL), WW (51 mg/mL), WB (6,5 mg/mL) y NP (5 mg/mL), obtenidos de la esponja. Los geles control se prepararon mezclando la misma cantidad de Phytagel y agua, y agregando la cantidad equivalente de metanol o diclorometano empleados en el proceso de disolución de las fracciones. Las cajas fueron amarradas bocabajo, con los geles en dirección al sustrato, en dos andamios de tubos de PVC, y se sumergieron a 8 m de profundidad en Punta de Betín (Santa Marta) durante 30 días. Los experimentos tuvieron lugar en marzo de 2005 y durante marzo y abril de 2006. Luego se sacaron del mar, teniendo cuidado de mantenerlas siempre sumergidas en agua marina, y se llevaron a los laboratorios de INVEMAR donde se colocaron en acuarios con flujo continuo de agua marina filtrada. Bajo el estereoscopio se evaluó la abundancia y porcentaje de cobertura de organismos sésiles. Se hicieron comparaciones entre estas variables en extractos crudos, fracciones y el control por medio de las pruebas estadísticas Kruskal-Wallis y Dunn y ANOVA (37).
RESULTADOS Y DISCUSIÓN
Actividad antifouling de los extractos de Cliona delitrix en campo
La esponja C. delitrix enseña una superficie libre de macroorganismos, por lo cual resultó interesante valorar la capacidad antifouling del extracto crudo y hacer un monitoreo bioguiado. Con tal fin se ensayaron las fracciones y el extracto crudo disueltos en Phytagel atendiendo la metodología de Pawlik (38). Como resultado de los ensayos en campo se encontró que el ECT tiene un efecto antifouling estadísticamente significativo como se indica em la tabla 1, pues tanto la abundancia como la cobertura de organismos epibiontes disminuyeron notablemente con su uso. En el ensayo se encontró también que la fracción no polar NP presentó el menor número de organismos en la superficie del gel y el menor porcentaje de cobertura, mientras la fracción acuosa WW y la butanólica WB no mostraron actividad significativa.
Tabla 1. Ensayo de actividad antifouling: abundancia y cobertura de organismos encontrados sobre los geles después de un mes en campo. Extracto y fracciones de C. delitrix.
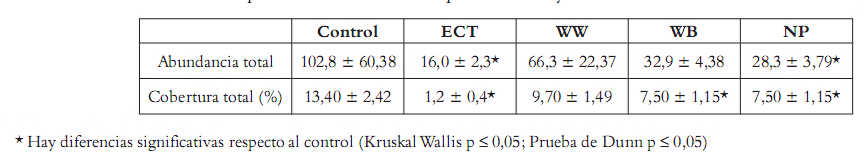
Así, se pudo establecer que en la fracción lipídica (NP) se concentró la actividad antifouling exhibida por el ECT de esta esponja.
Presencia de la serotonina en Cliona delitrix
De la subfracción butanólica WB de C. delitrix fue purificado como metabolito mayoritario la serotonina (5-hidroxitriptamina), reconocido neurotransmisor de los animales con sistema nervioso. La serotonina había sido previamente aislada como metabolito del extracto metanólico de la esponja Hyrtios reticulatus (32) y de un ejemplar de C. delitrix, recolectado en Salvador de Bahía, Brasil (12). Además, a partir de la subfracción acuosa WW, también inactiva al ensayo antifouling, se purificó otra amina heterocíclica identificada como imidazol-2(3H)-imina, la cual es parte estructural de varias sustancias farmacológicamente activas de compuestos naturales como los alcaloides marinos isonaamina, dorimidazol y preclatridina (33).
La actividad antifouling de la fracción no polar se puede considerar muy importante para la supervivencia de la esponja, porque le permite mantener su superficie libre de epibiontes, resistiendo a la ocupación de su espacio superficial por otros organismos. Se pueden considerar los compuestos responsables de esta actividad particularmente activos porque, por la gran cantidad de serotonina que contiene esta esponja, se esperaría un notable asentamiento larval (de cirripedios en particular), dadas las propiedades promotoras conocidas del asentamiento de la serotonina y sus agonistas (9, 39), situación que no se observó en nuestros ensayos de campo donde, por el contrario, la esponja presentó una superficie limpia. No obstante, es importante mencionar que no existe ninguna evidencia de que la serotonina sea liberada por este organismo hacia su superficie, y en consecuencia, las larvas de los invertebrados colonizadores no podrían detectar la presencia del inductor.
Por otra parte se sabe que a los lípidos de fuentes marinas, como es el caso de algunos esteroles (40), esfingolípidos (23), glicerolípidos y ácidos grasos
(8, 25) se les ha reconocido actividad antifouling. Esta clase de compuestos se detectaron también en la fracción bioactiva y no polar NP de C. delitrix, y por ello el estudio de la composición química de la fracción lipídica de la esponja resultó de gran interés.
Composición de ácidos grasos en la fracción lipídica
La cromatografía en columna permitió, a partir de la fracción no polar NP, la separación de fracciones enriquecidas en ésteres metílicos, glicéridos, glicolípidos, fosfolípidos y ácidos libres, como se describió en la sección experimental. Cada fracción fue identificada por su movilidad y aspecto del revelado por CCD con respecto a los patrones de los lípidos respectivos, así como por el análisis de los espectros de RMN de protónica y carbono. Los ácidos grasos componentes de cada fracción lipídica fueron luego transformados a sus ésteres metílicos y pirrolididas. La asignación estructural de los ácidos grasos se hizo mediante la comparación del parámetro cromatográfico ECL (longitud equivalente de cadena, de sus siglas en inglés) y el estudio de los espectros de masas de los ésteres metílicos, con los valores o con los espectros registrados en la literatura y los obtenidos por nosotros para muestras verdaderas (21, 41). La posición de las insaturaciones y de las ramificaciones se asignó mediante el estudio detallado del espectro de masas de las pirrolididas obtenidas (42, 43). Las técnicas aquí empleadas tienen una limitación: no es posible la asignación de la estereoquímica de los dobles enlaces si no se cuenta con los patrones de cada diastereoisómero. La cuantif icación se efectuó por el método de normalización de áreas, tanto en el TIC como en el cromatograma. En la tabla 2 pueden observarse los ésteres metílicos de los ácidos grasos identificados en la fracción de ésteres metílicos libres, y aquellos obtenidos por transformación química a partir de glicerolípidos, glicolípidos, fosfolípidos y ácidos libres, indicando su peso molecular y abundancia relativa en cada caso. Adicionalmente se indica el valor promedio de ECL para cada uno de ellos.
Tabla 2. Ácidos grasosa de las fracciones de ésteres libres, glicéridos, glicolípidos, fosfolípidos y ácidos grasos libres de la esponja marina Cliona delitrix.
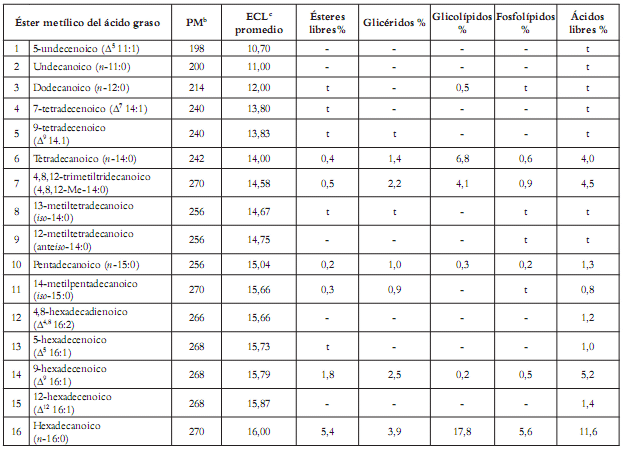
Dentro del conjunto de ácidos grasos de Cliona delitrix se destaca el ácido 7,10,12,14-icosatetraenoico (No 44), que por primera vez se reporta como un compuesto natural. Algunos otros, como el ácido 4,8-hexadecadienoico (No 12), ácido 6,9,12,14-icosatetraenoico (No 42) y el ácido 6,9,12,14,17-icosapentenoico (No 43), son muy poco frecuentes en la naturaleza, y se han hallado en otra de las especies de este género, la Cliona tenuis (21).
Con el fin de ilustrar el proceso de elucidación estructural se muestra el análisis del espectro de masas del compuesto 44: en el EM del éster metílico se observa el ión molecular m/z 318, que corresponde a la fórmula C21H34O2. La fragmentación es característica de un éster de ácido graso poliinsaturado debido a la presencia de los fragmentos característicos de la serie alquílica [CnH2n+1], la serie alquenílica [CnH2n-1], [CnH2n-3], la serie de fragmentos [CnH2n-5], que contiene el pico base m/z 79, y la serie [CnH2n-7], todos representativos de la existencia de dobles enlaces múltiples en la cadena hidrocarbonada, lo que indica que pertenecen a un ácido graso de 20 carbonos con cuatro dobles enlaces. El espectro de masas del derivado pirrolidida muestra como ión molecular m/z 357, correspondiente con la fórmula molecular C24H39NO, el ión m/z 98 [C5H8NO]+
y el ión m/z 113, resultante de una ruptura entre el C-2 y el C-3 y con transposición de hidrógeno del tipo McLafferty se indican en la figura 1.
Figura 1. Espectro de masas y los fragmentos diagnóstico de la pirrolidida del ácido 7,10,12,14-icosatetraenoico (No 44), obtenidos por espectrometría de masas en el modo de ionización electrónica.
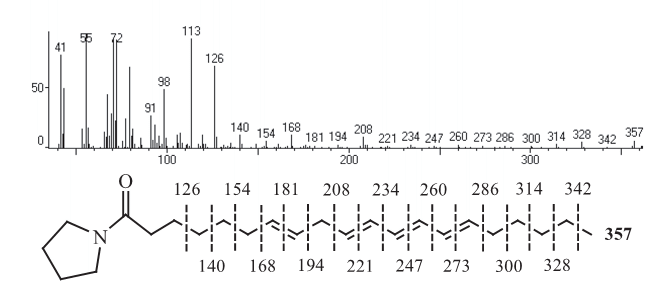
La forma general del espectro es típica de la pirrolidida de un ácido graso con cuatro dobles enlaces. A partir de los fragmentos observados se encuentra que los dobles enlaces se localizan en las posiciones C-7, C-10, C-12 y C-14, de acuerdo con la diferencia de 12 u.m.a entre los fragmentos m/z 181 y m/z 168, m/z 221 y 208, 247 y 234 y entre m/z 273 y m/z 260, así como la presencia de los fragmentos m/z 140, 154, 168, 194, 208, 234, 260, 286, 300, 314, 328 y m/z 342 (datos de ECL y de los espectros de masas se incluyen en la parte experimental). Todo lo anterior confirma la estructura del éster metílico del ácido 7,10,12,14-icosatetraenoico. El valor ECL de 19,46 de este compuesto no se encuentra reportado en la literatura para un ácido graso con las características antes mencionadas. La elucidación estructural del resto de los ésteres metílicos se hizo de la misma manera, permitiendo la identificación de 89 ácidos grasos en esta especie.
Al comparar la composición de ésteres metílicos de ácidos grasos en algunos lípidos de C. delitrix se halló que en la fracción de ésteres metílicos libres, el ácido 5,9-hexacosadienoico (No 79; 18,5%) es el más abundante, seguido del 6,9,12,14-icosatetraenoico (No 42; 8,8%), el ácido 17-hexacosenoico (No 82; 6%) y el hexadecanoico (No 16; 5,4%). En la fracción de glicéridos se pudo advertir que el ácido 5,9-hexacosadienoico (No 79; 12,6 %) es mayoritario; los ácidos restantes tienen cantidades inferiores al 10%, y se destacan el 6,9,12,14-icosatetraenoico (No 42; 6,7%) y el hexadecanoico (No 16; 3,9%). Entre los ácidos identificados en la fracción de glicolípidos se apreció que el hexadecanoico (No 16; 17,8%) prevalece, y que los restantes son inferiores al 10; son también abundantes el 5,9-hexacosadienoico (No 79; 9,3%), el tetradecanoico (No 6; 6,8%), el 5,9-octadecadienoico (No 28; 5,4%) y el octadecanoico (No 33; 5,1%). En la fracción de fosfolípidos se encontró que el ácido 5,9-hexacosadienoico (No 79; 27,7%) está en mayor cantidad. Todos los ácidos restantes de esta fracción son inferiores al 10%, como los ácidos 5,8,11,14-icosatetraenoico (No 41; 6,6%), hexadecanoico (No 16; 5,6%) y el 17-hexacosenoico (No 82; 5,4%). En la fracción de ácidos libres se encontró que los ácidos 5,9-hexacosadienoico (No 79; 24,9%) y hexadecanoico (No 16; 11,6%) se destacan. Los ácidos restantes son inferiores al 10%, entre ellos los ácidos 9-hexadecenoico (No 14; 5,2%), 4,8,12-trimetiltridecanoico (No 7; 4,5%) y el tetradecanoico (No 6; 4,0%).
Con respecto a la composición de ésteres metílicos agrupados por características estructurales en C. delitrix, se encontró que los ácidos grasos de cadena par y no ramificada predominan en todas las clases de lípidos, como se evidencia en la figura 2a y en la figura 2b, respectivamente.
Figura 2. Abundancia relativa de los ácidos grasos agrupados por características estructurales y tipos de lípidos en C. delitrix. 1. Ésteres metílicos libres, 2. Glicéridos, 3. Glicolípidos, 4. Fosfolípidos y 5. Ácidos grasos libres.
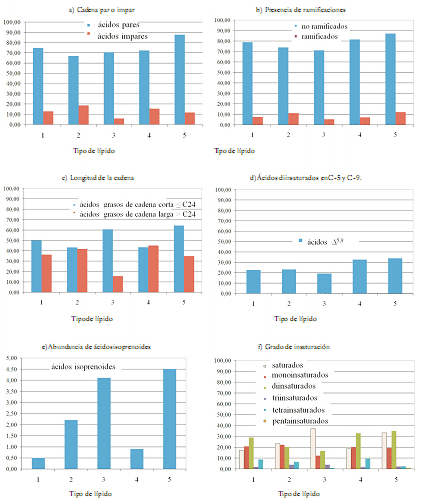
En la figura 2c, se observa que los ácidos demospóngicos, aquellos con más de 24 átomos de carbono (44), prevalecen sólo en la fracción de fosfolípidos, donde están en un 1,5% más que los de cadena más corta; en los otros casos son minoritarios; no obstante, en todas las clases de lípidos aquí estudiadas, su proporción es superior al 10%, y llega a ser casi la mitad en los glicerolípidos. Se puede apreciar que los ácidos grasos con cadena Δ5,9, indicados en la figura 2d, se distinguen en C. delitrix y que, en general, su concentración es superior al 20% en toda la variedad de lípidos estudiados, lo cual ha mostrado ser común en porífera y no en otros taxones.
En cuanto a los ácidos grasos tipo isoprenoide, de acuerdo con la figura 2e, se puede afirmar que son minoritarios, el ácido 4,8,12-trimetiltridecanoico No 7 (TMTD) es poco cuantioso, pues representa menos del 5% de la mezcla total de ácidos en todos los lípidos, corresponde a 4,5% de los ácidos totales identificados en la fracción de ácidos libres, 4,1% en la fracción de glicolípidos y 2,2% en la fracción de glicéridos.
De acuerdo con las investigaciones precedentes para esponjas del género Cliona, era de esperarse una mayor abundancia de ácidos isoprenoides porque los ácidos grasos tipo terpenoide: 15, 18, 21, 24-triacontatetraenoico y 4, 8, 12-trimetiltridecanoico han sido propuestos como quimiotrazadores para las familias Spirastrellidae y Clionidae, donde se ubica éste género (20). Pero este dato no se verifica en nuestros resultados con C. delitrix y C. tenuis (21). En otras investigaciones, a los ácidos grasos con cadena ramificada se les atribuye un origen bacteriano, en el caso específico del TMTD la transformación del fitol, que a su vez se produce por la transformación de la clorofila por cianobacterias (45). Esto concuerda con la observación en campo, en el sentido de que C. delitrix no tiene zooxantelas asociadas, en contraparte con las otras especies de Cliona presentes en el Caribe Colombiano. Es importante anotar que los ácidos grasos insaturados son elevados en todas las clases de lípidos estudiados, con una distribución heterogénea y una preponderancia de los ácidos grasos mono- y diinsaturados, como se evidencia en la figura 2f.
Otros estudios acerca de lípidos de esponjas del género Cliona en cuanto a la composición de sus ácidos grasos, muestra que en C. celata, (18), esponja recolectada en el Atlántico noroccidental, los ácidos demospóngicos son mayoritarios (79%), con gran representación de ácidos de más de 30 átomos de carbono, seguidos por los de 25 y 27 carbonos, mientras en C. aprica, esponja recolectada en el mar Caribe (20), son minoritarios (40%), pero se destaca la abundancia de los ácidos tetracosanoico (16,7%) y 5,9-hexacosadienoico (15,1%). Nuestros resultados concuerdan con los de C. aprica, ya que los ácidos desmospóngicos en C. delitrix están en una proporción cercana al 40%; sobresale también la presencia del ácido 5,9-hexacosadienoico en la fracción de fosfolípidos. No se halló correlación entre las otras clases de ácidos grasos de C. delitrix con los de estas dos especies de esponjas, en tanto que al comparar los ácidos grasos de C. delitrix con los de Cliona tenuis, se encuentra que los de cadena par y no ramificados igualmente predominaron entre todos los lípidos de C. tenuis, al igual que los ácidos grasos mono y diinsaturados, excepto en la fracción de ácidos grasos libres, donde se hallaron en abundancia inferior al 10% en C. tenuis (21). Los ácidos de cadena larga, en cambio, se observan en menor porcentaje entre los diferentes lípidos de C. tenuis y prevalecen sólo entre los glicolípidos (un 40%). La abundancia relativa de los ácidos grasos de cadena Δ5,9 fue tan baja como en C. delitrix, aunque más heterogénea, del 35,7% en la fracción de ésteres metílicos libres y tan sólo de un 2.6% en la de los ácidos libres de C. tenuis. Similar comportamiento tienen los ácidos grasos tipo isoprenoide, que representan el 3,4% de la fracción de fosfolípidos, el 2,1% de la fracción de glicolípidos y están ausentes entre los ésteres metílicos libres de C. tenuis.
Por otra parte, el ácido 5,8,11,14-icosatetraenoico, No 41 (ácido araquidónico), presente en fosfolípidos y glicolípidos de Cliona delitrix, y el ácido 9-octadecenoico No 31 (ácido palmitoleico), presente en todas las mezclas de compuestos estudiados, pueden contribuir a las propiedades antiepibióticas de la fracción lipídica, por sus reconocidas características antifouling (8, 25).
Composición de monohidroxiesteroles en la fracción lipídica
Las esponjas representan una rica fuente de esteroles con estructuras sin precedentes, cuyas combinaciones pueden caracterizar a nivel taxonómico algunos órdenes, familias y géneros del phylum. A continuación, informamos también sobre el conjunto de esteroles identificados en la fracción no polar de Cliona delitrix, datos que no han sido publicados con anterioridad.
En la tabla 3 aparece la identidad de cada esterol identificado, su tiempo de retención relativo en CGAR o en CLAE, obtenidos como se describió en la parte experimental.
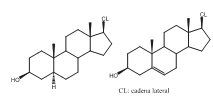
Como puede observarse en la tabla 3, se identificaron para la esponja C. delitrix 19 monohidroxiesteroles diferentes, cuyas cadenas y núcleos convencionales se indican en la figura 3 .
Tabla 3. Composición de monohidroxiesteroles de C. delitrix.
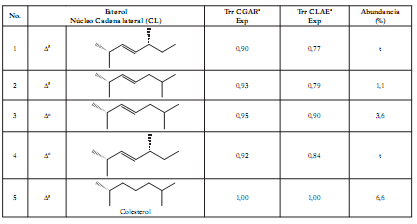
Figura 3. Estructura general de los núcleos de esteroles identificados en C. delitrix.
Son los más abundantes el colestanol (No 6; 24,8%), clionasterol (No 16; 18,1%), (22E, 24R*)24-metilcolesta-5,22-dien-3b-ol (No 8; 16,1%), sitosterol (No 17; 12%), poriferasterol (No 13; 9,1%) y el colesterol (No 5; 6,6%). Todos estos esteroles son de amplia distribución en el phylum Porifera. Los esteroles del género Cliona han sido estudiados desde el siglo pasado y, según Valentine y Bergmann (46), es el clionasterol el más sobresaliente. En otras esponjas de este género, como C. caribbaea (16), C. celata (17), C. viridis (19) y C. aprica (20), se han hallado también en gran cantidad el clionasterol y colesterol. Sin embargo, en Cliona delitrix resulta bastante notoria la abundancia de estanoles Δ0, como el colestanol (cercana al 25%), puesto que en ninguna de las otras especies del género Cliona hasta ahora estudiadas en la bibliografía, pueden observarse este tipo de esteroles.
Finalmente, aunque en la literatura no exista evidencia de esteroles como los anteriores con actividad antifouling, otros estudios han demostrado cómo algunos esteroides de esponjas sí poseen dicha bioactividad, Así, de la esponja marina Lendenfeldia chondrodes se aislaron cuatro epidioxi esteroles con actividad antifouling contra el mejillón Mytilus edulis galloprovincialis (40), y de la esponja Acanthella cavernosa se aislaron tres A-noresteroles, con actividad antifouling debida a la inhibición del asentamiento de Balanus albicostatus (47).
CONCLUSIONES
En conclusión, en un ensayo en campo se encontró que la fracción lipídica del extracto de C. delitrix era la responsable de la actividad antifouling, porque condujo al menor porcentaje de recubrimiento superficial por parte de otros organismos. Se reconoció que aunque la serotonina, inductor del asentamiento larval, es abundante en C. delitrix, su superficie se encuentra marcadamente limpia, probablemente porque este metabolito no es liberado por el organismo hacia su superficie, o porque contiene compuestos con actividad antifouling muy potente. Se determinó también que la fracción lipídica está constituida por ésteres metílicos, glicéridos, glicolípidos, fosfolípidos, ácidos libres y esteroles. En cada fracción se estableció el contenido de ácidos grasos, identificando el ácido 7,10,12,14-icosatetraenoico, no reportado antes en la literatura, y 88 ácidos conocidos, además de 19 esteroles comunes. Este trabajo es el primero acerca de la caracterización lipídica de la esponja marina Cliona delitrix.
AGRADECIMIENTOS
Agradecemos a Colciencias (COL-20101007015), Fundación para la promoción de la investigación y la tecnología (Banco de la República), y a la Universidad Nacional de Colombia (DIB), por la financiación del presente estudio. Leonardo Castellanos agradece a la Universidad Nacional de Colombia por la comisión de estudios de doctorado.
REFERENCIAS BIBLIOGRÁFICAS
1. McClintock B, Baker B. Marine chemical ecology. New York, USA: CRC press; 2001. 610 p. [ Links ]
2. Blunt JW, Copp BR, Hu WP, Munro HG, Northcote PT, Prinsep MR. Marine natural products. Nat Prod Rep. 2009 Feb; 26 (2): 170-244. [ Links ]
3. Hedner E, Sjögren M, Hodzic S, Andersson R, Göransson U, Jonsson P, et al. Antifouling activity of a dibrominated cyclopeptide from the marine sponge Geodia barretti. J Nat Prod. 2008 Mar; 71 (3): 330–333. [ Links ]
4. Fusetani N, Clare A. Antifouling compounds. Progress in molecular and subcellular biology. Subseries: marine molecular biotechnology. Berlin, Germany: Springer-Verlag; 2006. 225 p. [ Links ]
5. Cronin G. Resource allocation in seaweeds and marine invertebrates: chemical defense patterns in relation to defense theories. En: McClintock B, Baker B, editors. Marine chemical ecology. New York: CRC press; 2001. p. 325-353. [ Links ]
6. Singh N, Turner A. Leaching of copper and zinc from spent antifouling paint particles. Environ Pollut. 2009 Feb; 157 (2): 371–376. [ Links ]
7. Mol L, Raveendran T, Parameswaran P. Antifouling activity exhibited by secondary metabolites of the marine sponge, Haliclona exigua (Kirkpatrick). Int Biodeter Biodegr. 2009 Jan; 63 (1): 67–72. [ Links ]
8. Ma A, Deng Z, Ofwegen L, Bayer M. Proksch P, Lin W. Dendronpholides A-R, cembranoid diterpenes from the chinese soft coral Dendronephthya sp. J Nat Prod. 2008 Jul; 71 (7): 1152–1160. [ Links ]
9. Dahlström M, Elwing E. Adrenoceptor and other pharmacoactive compounds as putative antifoulants. En: Fusetani N, Clare A, editors. Antifouling compounds. Progress in molecular and subcellular biology. Subseries: marine molecular biotechnology. Berlin, Germany: Springer-Verlag; 2006. p. 171-202. [ Links ]
10. Chaves A, López M, Parra F, Zea S. Ecología química de las esponjas excavadoras Cliona aprica, C. caribbaea, C. delitrix y C. tenuis. Bol Inst Invest Mar Cost. 2005 Dec; 34 (1): 43-67. [ Links ]
11. Chaves A, Castellanos L, Zea S. Duque C, Rodríguez J, Jiménez C. Clionapyrrolidine A-a metabolite from the encrusting and excavating sponge Cliona tenuis that kills coral tissue upon contact. J Chem Ecol. 2008 Dec; 34 (12): 1565-1574. [ Links ]
12. Granato A, De Oliveira J, Seleghim M, Berlinck R, Macedo M, Ferreira A, et al. Produtos naturais da ascídia Botrylloides giganteum, das esponjas Verongula gigantea, Ircinia felix, Cliona delitrix e do nudibrânquio Tambja eliora, da costa do Brasil. Quim Nova. 2005 Mar; 28 (2): 192-198. [ Links ]
13. Palermo JA, Rodríguez MF, Cabezas E, Balzaretti V, Seldes AM. Celenamide E a tripeptide alkaloid from the patagonian sponge Cliona chilensis. J Nat Prod. 1998 Apr; 61 (4): 488-490. [ Links ]
14. Keyzers R, Daoust J, Davies-Coleman M, Van R, Balgi A, Donohue E, et al. Autophagy-modulating aminosteroids isolated from the sponge Cliona celata. Org Lett. 2008 Jul 17; 10 (14): 2959-2962. [ Links ]
15. Queiroz A, Silva R, Moura R, Dreyfuss J, Paredes-Gamero E, Souza A, et al. Growth inhibitory activity of a novel lectin from Cliona varians against K562 human erythroleukemia cells. Cancer Chemoth Pharm. 2009 May; 63 (6): 1023-1033. [ Links ]
16. Bergmann W, McTigue FH, Low EM, Stokes WM, Feeney RJ. Marine products. XXVI. Sterols from sponges of the family Suberitidae. J Org Chem. 1949 Jan; 15 (1): 96-105. [ Links ]
17. Erdman TR, Thomson RH. Sterols from the sponge Cliona celata Grant and Hymeniacidon perleve Montagu. Tetrahedron. 1972 Jun; 28 (20): 5163-5173. [ Links ]
18. Litchfield C, Tyszkiewicz J, Marcantonio EE, Noto G. 15, 18, 21, 24-triacontatetraenoic and 15, 18, 21, 24, 27-triacontapentaenoic acids: new C30 fatty acids from the marine sponge Cliona celata. Lipids. 1979 Jul; 14 (7): 619-622. [ Links ]
19. Sica D, De Simone F, Ramundo E, Zollo F. Sterols from some sponges. Biochem Syst Ecol. 1978; 6 (1): 77-79. [ Links ]
20. Carballeira NM, Maldonado ME, Rivera E, Porras B. The fatty acid 4, 8, 12-trimethyltridecanoic as a common constituent of the phospholipids of the sponge families Spirastrellidae and Clionidae. Biochem Syst Ecol. 1989 Jul 28; 17 (4): 311-314. [ Links ]
21. Castellanos L, Duque C. Composición química y actividad antifouling de la fracción lipídica de la esponja marina Cliona tenuis (Clionidae). Rev Col Quim. 2008 Sep-Dec; 37 (3): 259-274. [ Links ]
22. Costantino V, Fattorusso E, Imperatore C, Mangoni A, Freigang S, Teyton L. Corrugoside, a new immunostimulatory alpha-galactoglycosphingolipid from the marine sponge Axinella corrugata. Bioorgan Med Chem. 2008 Feb 15; 16 (4): 2077–2085. [ Links ]
23. Hattori T, Adachi K, Shizuri Y. New ceramide from marine sponge Haliclona koremella and related compounds as antifouling substances against macroalgae. J Nat Prod. 1998 May 9; 61 (6): 823-826. [ Links ]
24. Carballeira NM, Pagan M. Identification and total synthesis of a novel dimethylated fatty acid from the Caribbean sponge Calyx podatypa. J Nat Prod. 2000 Mar 25; 63 (5): 666-669. [ Links ]
25. Omae I. General aspects of tin-free antifouling paints. Chem Rev. 2003 Aug 14; 103 (9): 3431-3448. [ Links ]
26. Carballeira NM, ONeill R, Parang K. Synthesis and antifungal properties of alpha-methoxy and alpha-hydroxyl substituted 4-thiatetradecanoic acids. Chem Phys Lipids. 2007 Nov; 150 (1): 82-88. [ Links ]
27. Carballeira NM, ONeill R, Silva D. First total synthesis of (5Z,9Z)-(±)-2-methoxy-5,9-octadecadienoic acid, a marine derived methoxylated analog of taxoleic acid. Chem Phys Lipids. 2008 Nov; 156 (1-2): 41-44. [ Links ]
28. Kupchan SM, Britton RW, Lacadie JA, Ziegler MF, Sigel CW. The Isolation and structural elucidation of bruceantin and bruceantinol, new potent antileukemic quassinoids from Brucea antidysenterica. J Org Chem. 1975 Mar 7; 40 (5): 648–654. [ Links ]
29. Privett OS, Dougherty KA, Erdahl WL, Stolyhwo A. Studies on the lipid composition of developing soybeans. J Am Oil Chem Soc. 1973 Dec; 50 (12): 516-520. [ Links ]
30. Wood AT, Hall MR. Reversed-phase high-performance liquid chromatography of catecholamines and indoleamines using a simple gradient solvent system and native f luorescence detection. J Chromatogr B. 2000 Jul 7; 744 (1): 221-225. [ Links ]
31. Wrona M Z, Dryhurst G. Oxydation chemistry of 5-hydroxytryptamine I. Mechanism and product formed at micromolar concentratios. J Org Chem. 1987 Jun; 52 (13): 2817-2825. [ Links ]
32. Salmoun M, Devijver C, Daloze D, Braekman JC, Van Soest RW. 5-Hydroxytryptamine-derived alkaloids from two marine sponges of the genus Hyrtios. J Nat Prod. 2002 Jun 28; 65 (8): 1173-1176. [ Links ]
33. Weinmann H, Harre M, Koenig K, Merten E, Tilstam U. Efficient and environmentally friendly synthesis of 2-aminoimidazole. Tetrahedron Lett. 2002 Jan 21; 43 (4): 593-595. [ Links ]
34. Morrison WR, Smith LM. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron f luoride-methanol. J Lipid Res. 1964 Oct; 5: 600-608. [ Links ]
35. Castellanos L, Zea S, Osorno O, Duque C. Phylogenetic analysis of the order Halichondrida (Porifera, Demospongiae), using 3β-hydroxisterols as chemical characters. Biochem Syst Ecol. 2003 Oct; 31 (10): 1163-1183. [ Links ]
36. Henrikson, A, Pawlik J. Seasonal variation in biofouling of gels containing extracts of marine organisms. Biofouling. 1998 Jan 1; 12 (1-3): 245-255. [ Links ]
37. Siegel S, Castellan NJ. Nonparametric statistics for the behavioral sciences. New York, USA: McGraw-Hill; 1988. 399 p. [ Links ]
38. Pawlik JR, Steindler L, Henkel TP, Beer V, Ilan M. Chemical warfare on coral reefs: Sponge metabolites differentially affect coral symbiosis in situ. Limnol Oceanogr. 2007; 52 (2): 907–911. [ Links ]
39. Yamamoto H, Shimizu K, Tachibana A, Fusetani N. Roles of dopamine and serotonin in larval attachment of the barnacle, Balanus amphitrite. J Exp Zool. 1999 Dec 1; 284 (7): 746-758. [ Links ]
40. Sera Y, Adachi K, Shizuri Y. A New epidioxy sterol as an antifouling substance from a Palauan marine sponge, Lendenfeldia chondrodes. J Nat Prod. 1999 Oct 31; 62 (1): 152-154. [ Links ]
41. Rodriguez WP. Composición de ácidos grasos ligados a fosfolípidos en esponjas marinas del orden Halichondrida (Polifera, Demospongiae), del Caribe colombiano y su significancia como carácter químico de clasificación. [Tesis de maestría]. Bogotá: Universidad Nacional de Colombia; 2002. [ Links ]
42. Andersson BA. Mass spectrometry of fatty acid pyrrolidides. Prog Chem Fats other Lipids. 1978; 16 (1): 279-308. [ Links ]
43. Christie W. The lipid library [Internet]. United kingdom: The American Oil Chemists Society. 2010. [updated 2010 Apr 22 cited 2010 May 26]; Available from: http://www.lipidlibrary. co.uk. Consultado 6 de agosto de 2009. [ Links ]
44. Lawson MP, Thompson JE, Djerassi C. Cell membrane localization of long chain C24-C30 fatty acids in two marine demosponges. Lipids. 1988 Aug; 23 (8): 741-749. [ Links ]
45. Gillan FT, Stoilov IL, Thompson JE, Hogg RW, Wilkinson CR, Djerassi C. Fatty acids as biological markers for bacterial symbionts in sponges. Lipids. 1988 Dec; 23 (12): 1139-1145. [ Links ]
46. Valentine FR, Bergmann W. Contributions to the study of marine products. VIII. The sterol of sponges: Clionasterol and poriferasterol. J Org Chem. 1941 May; 6 (3): 452-461. [ Links ]
47. Qiu Y, Wei Z, Xu M, Li Q, Han W. New A-nor steroids and their antifouling activity from the Chinese marine sponge Acanthella cavernosa. Steroids. 2008 Dec 22; 73 (14): 1500-1504. [ Links ]
Recibido: Octubre 29 de 2009; Aceptado: Junio 01 de 2010
* Autor a quien debe dirigir la correspondencia: cduqueb@unal.edu.co

