










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkVitae
Print version ISSN 0121-4004
Vitae vol.19 no.1 Medellín Jan./Apr. 2012
BIOTECHNOLOGY
ANALYSIS OF THE EFFECT OF THE INTERACTIONS AMONG THREE PROCESSING VARIABLES FOR THE PRODUCTION OF EXOPOLYSACCHARIDES IN THE MICROALGAE Scenedesmus obliquus (UTEX 393)
ANÁLISIS DEL EFECTO DE LAS INTERACCIONES DE TRES VARIABLES DE PROCESO PARA LA PRODUCCIÓN DE EXOPOLISACÁRIDOS EN LA MICROALGA Scenedesmus obliquus (UTEX 393)
Nancy M. CÓRDOBA-CASTRO1, Andrés M. MONTENEGRO-JARAMILLO1, Rosa E. PRIETO1, Gloria E. GONZÁLEZ-MARIÑO1*
1 Universidad de La Sabana. Facultad de Ingeniería. Campus Universitario Puente del Común. Km. 7. Autopista Norte de Bogotá Chía, Colombia.
* Corresponding author: gloria.gonzalez@unisabana.edu.co.
Received: 19 October 2010
Accepted: 22 November 2011
ABSTRACT
Microalgae are capable of producing biomolecules that have a wide variety of applications in agriculture, food industry, and medicine. In this study, three process variables are evaluated in order to determine its incidence on biomass and exopolysaccharides production. The effect of light intensity, agitation and carbon concentration on Scenedesmus obliquus (UTEX 393) growth and expolysaccaharides production is evaluated using 23 factorial design through the screening methodology. The simultaneous effect of level variation for three different experimental variables is examined in the present study in three levels for each parameter (Light intensity: 80, 130, 180 μE m-2 s-1, Agitation: 0, 600, 1200 rpm, carbon concentration 0, 2, 4% v/v Air-CO2). Specific growth rate and the exopolysaccharides concentration are the selected response variables. Results show that the optimal conditions for the two response variables correspond to the maximum levels of the three experimental variables (180 μE m-2 s-1, 4% air-CO2, and 1200 rpm), obtaining a specific growth rate of 0.64 d-1 and a exopolysaccharides concentration of 24.7 mg L-1. A significant interaction between the variables is observed, which has direct effects on cellular growth and exopolysaccharides production. The EPS production is facilitated by the turbulent flow (agitation maximum level), which is associated with a higher availability and better distribution of energy sources (light) and carbon dioxide. The validation of polynomials models verifies the relevance of the analysis performed.
Keywords: Algae, Scenedesmus, exopolysaccharides, process parameters, experimental design.
RESUMEN
Las microalgas son capaces de producir biomoléculas que poseen diversidad de aplicaciones relacionadas con la agricultura, alimentos y medicina. En la presente investigación se evalúan tres variables de proceso para determinar su incidencia e interacciones en la producción de biomasa y exopolisacáridos. El efecto de la intensidad de la luz, la agitación y la concentración de carbono sobre el crecimiento y la producción de exopolisacáridos en Scenedesmus obliquus (UTEX 393) es evaluada aplicando un diseño experimental factorial 23 por metodología de Screening. Se evalúa el efecto en simultáneo de la variación de tres variables experimentales en tres niveles (Intensidad de luz: 80, 130, 180 μE m-2 s-1, Agitación: 0, 600, 1200 rpm, concentración de carbono: 0, 2, 4% v/v Air-CO2). El coeficiente específico de crecimiento y la concentración de exopolisacáridos son las variables de respuesta seleccionadas. Los resultados muestran condiciones óptimas para las dos variables de respuesta en los niveles máximos de las variables experimentales (180 μE m-2 s-1, 4% mezcla CO2-aire y 1200 rpm), obteniendo una coeficiente específico de crecimiento 0.64 d-1 y una concentración de exopolisacáridos de 24,7 mg L-1. Se muestra una marcada influencia de la interacción entre variables que repercuten directamente sobre el crecimiento celular y la producción de exopolisacáridos, está ultima favorecida por los regímenes de flujo turbulentos (máximo nivel de agitación) que se asocia a una mayor disponibilidad y mejor distribución de la fuente de energía (luz) y carbono. La validación de los modelos polinómicos en montajes adicionales, certifica la representatividad de los análisis realizados.
Palabras clave: algas, Scenedesmus, exopolisacáridos, parámetros de proceso, diseño experimental.
INTRODUCTION
In recent years, there have been a large number of reports on the potential of microalgae as a valuable source of products such as phycobiliproteins, polysaccharides, proteins, fatty acids, and other secondary metabolites (1-3). Currently, the range of applications for biomolecules isolated from the diverse classes of algae is enormous; with great potential for the development of applications in human and animal nutrition, cosmetics, fatty acids, and pigments (4-5).
The polysaccharides of microalgae have potential applications in many industrial sectors due to their diverse physical and chemical interesting properties, such as stabilizing, suspending, thickening, gelling, and water-retention capability. They are used in textiles, adhesives, paints, food, and beverage industries; as well as in pharmaceuticals, in oil and metal recovery from ore, and industrial wastes (6).
The cell growth and accumulation of metabolites in microalgae culture is affected by many factors, these include medium components such as phosphate and nitrogen sources, besides culture conditions, namely, temperature, light intensity, aeration rate, and initial pH (7). Therefore, for commercial applications is relevant to determine the significant variables that affect production and yield of biomass and biomolecules, in order to generate reproducible and cost effective processes at industrial level.
Polysaccharide biosynthesis and accumulation generally take place after the growth phase of the microorganism. The polysaccharides produced by microorganisms can be classified into three main groups according to their location in the cell: (a) cytosolic polysaccharides, which provide carbon and energy source for the cell; (b) polysaccharides that make-up the cell wall, including peptidoglycans and lipopolysaccharides and (c) polysaccharides that are exuded into the extracellular environment in the form of capsules or biofilm, known as exopolysaccharides (8).
Several authors have documented the potential of microalgae, like the Chlorophyta division. Specifically, these authors have shown that the genera Scenedesmus sp. and Chlorella sp. have the potential to produce polymeric substances and growth promoters for diverse industrial and agro-industrial applications (9-10). The genera Scenedesmus, which can be found in all kinds of freshwater, are widely used for biomass production and provide appropriate experimental materials for the study of photosynthesis and other fundamental problems in biochemistry and physiology, which can impact production systems (11).
This article analyzes the effect of the interactions of three process variables through a factorial experimental design using the screening methodology, in the production of exopolysaccharide-rich extracts from the microalgae Scenedesmus obliquus (UTEX 393) that have the potential to be used in innovative agricultural products.
MATERIALS AND METHODS
Organisms and growing conditions
The present study used the green microalga Scenedesmus obliquus (UTEX 393) as the reference microorganism. The microalga was obtained from UTEX culture collection (University of Texas, Austin, TX, USA). The green microalga, was cultivated in batch culture under sterile conditions in Bristol liquid medium (UTEX), which was composed of (per liter) 0.25 g NaNO3, 0.025 g CaCl2.2H20, 0.075 g MgSO4.7H2O, 0.075 g K2HPO4, 0.175 g KH2PO4, 0.025 g NaCl, 6ml PVI Metal solution, 1ml of thiamine and 1ml of biotin. All experiments were adjusted to an initial pH of 6.8. The cultivation was carried out in photobioreactors which are glass vessels of 500ml. Each culture was inoculated with an initial S. obliquus cell concentration of 1x104 cell/ml.
The cultures were stirred using a magnetic stirrer base. The cultivation vessels were kept at a constant temperature of 27 ± 2°C. The agitation was performed with a drive magnetic base (figure 1), allowing to change the shaking rate (Rf). The light intensity (μE m-2 s-1) was measured at the geometric center for each container with a Biospherical Instrument Inc. QSFL 2101 light meter and the intensity was adjusted by modifying the capacity (Watt) of the source of light. Lamps42watt, initially located 10 cm from the bottles, were used for growth. The carbon concentration in the mixture, C * (%air-CO2 v/v) was controlled by pressure adjustments in the discharge system.

Monitoring biomass concentration
Biomass values (Yc, cells ml-1) were quantified using a particle counter (Beckman®, USA) with duplicate readings. In addition, the cell concentration measurements were validated by measuring the optical density of the culture at 560 nm for S. obliquus (12). For the optical density measurements of the culture, glass cuvettes of 1 cm3 were used, and absorbance measurements were performed on a Varian-Cary 100 spectrophotometer equipped with lamps for visible and UV light. Biomass values (Yc) and exponential regressions were used to calculate the maximum specific growth rate (μmax, d-1) using equation 1 (13), during the exponential growth phase in the culture.

where: μ = specific growth rate; Nt = population size at the end of the time interval; N0 = population size at the beginning of the time interval; Δt = time interval.
Total carbohydrates and exopolysaccharides
Two independent procedures (described below) were standardized for the quantification of intracellular and extracellular carbohydrates.
Analysis of total intracellular carbohydrates
The concentration of intracellular carbohydrates was performed by the anthrone-sulfuric acid method. The extraction of microalgae biomass carbohydrates was performed taking into account the methodology used by Yemm and Willis (14) and modified by Band (15). In this protocol, the cells undergo a process of hydrolysis for 24 hours in a chloroform-methanol mixture, after which several washes are performed, and the mixture is centrifuged at 4000 rpm for 5 minutes. Then, HCl is added, and the mixture is boiled for one hour. After this period of time, one final centrifugation is performed with the conditions previously described, and the obtained supernatant is analyzed using the anthrone reagent with a previously prepared calibration curve.
Analysis of total extracellular carbohydrates or exopolysaccharides
Analysis of total extracellular carbohydrates or exopolysaccharides in the culture supernatant was performed taking into account the methodology described by Lewin, 1956 (16) and modified by Yu et al., 2010 (17). In this protocol, the supernatant is clarified by filtration, and then the carbohydrates are precipitated with ethanol to be subsequently concentrated by evaporation. Free extracts of organic material are weighed to determine the amount of carbohydrates present.
Dynamic viscosity and density of the culture medium
The quantification of the dynamic viscosity (μ, mPa*s) of the culture medium during the growth of microalgae was performed using a Brookfield viscometer, LDVD-1 + T. The corresponding density (ρ, g ml-1) and viscosity measurements were performed according to methods described by Wazer, Lyons et al., 1963 (18). The final value reported is the average of three replicates. The values of μ and ρ were used to calculate the impeller Reynolds number (Rei) according to equation 2. These values were used for the analysis of the experiments, the results of the optimization and validation assays.
Rei= Ni Di2ρ / μ Equation 2.
where: Ni is the speed of the impeller, and Di is the diameter of the impeller.
Experimental design
The three studied factors were agitation (Rf, rpm), light intensity (Ia,μE m-2 s-1) and carbon concentration (C *, a mixture of air enriched with CO2 in % v/v) which were evaluated at three different levels, using a 23 screening factorial design. The choice of factors levels was based on information from literature and preliminary experiments. Nine experiments were carried out in randomized run order (9 points of factorial design and three center points to establish experimental errors) (table 1). The response variables were the specific growth rate (μ, d-1) and concentration of exopolysaccharides (EPS) (Ych, mg L-1).The analysis of responses for each test was performed using the statistical software Statgraphics plus 5.0.
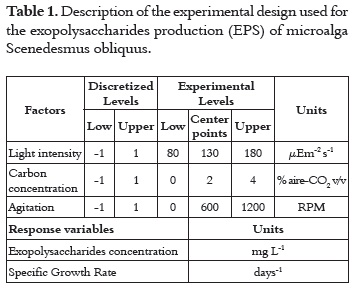
RESULTS AND DISCUSSION
To investigate the effects of light intensity, agitation and carbon concentration and their interactions on the Scenedesmus obliquus growth cell and EPS production, a batch culture was carried out in the Bristol medium for 20 days. The factors were maintained at the values shown in table 2. The cell growth and the EPS production curves of the 12 individual experiments run, showed differences among them. The day 20 was chosen for the analysis of EPS and as the end point of the culture, based on cell behavior in preliminary experimental runs.
Specific Growth Rate (μ, d-1) as a response variable
The highest growth rate (0.66 day-1) was observed for the culture grown at 180 μE m-2s-1/ 4% air-CO2 %v/v/ 1200 rpm and the l owest growth rate (0.27 day-1) was observed for the culture grown at 80 μE m-2s-1/ 0% air-CO2 %v/v/ 1200 rpm. For the specific growth rate (μ, d-1), the effects of light intensity and carbon concentration were significant because they present P values lower than 0.05 at the 95% confidence level. The effects of each factor and their interaction, as well as statistical significance, are reported in table 3.
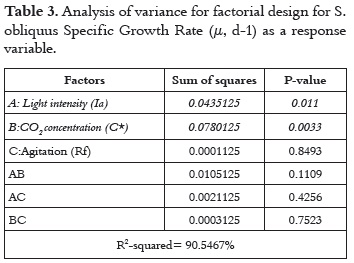
The significance of light intensity and carbon concentration on specific growth rate shows that these variables maximize the response at maximum levels. Therefore, it is clear that the dynamics of the chemical reactions in photo-biological processes are directly related to the supply of CO2 and the intensity of the light source used. In conditions of relatively constant temperature and pH, these factors would control processes or reactions related to the cell productivity of the culture. The value of the R-squared statistic supports this statement because the results obtained explain 90.55% of the variability of the specific growth rate.
The results show that agitation-Rf, rpm- (B factor) has a low effect, because it has a P-value > 0.05. This implies a low relevance of this variable in the cell growth. In consequence, this suggest that besides the broad significance of the variables A and B, all their interactions have a positive effect for maximizing the specific growth rate (μ, d-1). This high significance for variables A and B, and low for agitation (C) and other interactions between variables, can be seen most clearly in the Pareto diagram (figure 2).
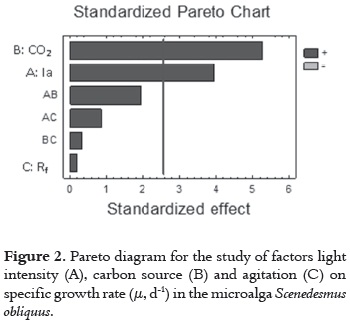
According to the interaction effects between experimental factors on the response, the AB interaction shows that maximum levels of CO2 and light intensity maximize the productivity of cells. The same effect was noticed with AC interactions (light intensity and agitation) and BC (carbon concentration and agitation), which at maximum levels also generate this effect (figure 3).
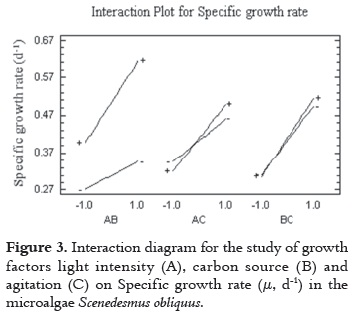
When the concentration of CO2 was found in its lowest level (0% air-CO2), the specific growth rate had the lowest value. A similar effect occurs when light intensity was located at this level. The relevance of the main effect of variable B is evident because when this variable was kept at 0% (minimum level) the lowest specific growth rate of the experiment run was obtained.
The AC interaction shows that maximum levels of light intensity and agitation are required to achieve a higher cell growth. These results show that effect achieves greater cell growth (0.66 day-1) in the experimental run when the factors are placed in these levels (180 μE m-2s-1, 4% Air-CO2 %v/v, 1200 rpm).
Much of the agitation effect (interactions AC, BC) on specific growth rate could be associated with mechanical process related to the physiology and cell stability in culture medium. However, in photoautotrophs production systems, the flow dynamics (RPM) affects the distribution of light associated with the scattering cell in the medium and the selfshading effects generated in high density cultures. Considering cultures with constant cell density, it should be noted that those systems with a turbulent shaking rate provide cells with shorter light/dark cycles (L/D) than those cultures subjected to a shaking rate of low turbulence or one that is laminar (19). As a result, the productivity (μ, d-1) in relation to the L/D cycles is affected by light intensity, such that the greater the intensity, the lower the cycle must be. In other words, the higher the intensity of light or luminous flux, the shorter the period of exposure to light should be to avoid loss of productivity. Studies by Wu, et al., 2001 (20) on processes of photo-inhibition support these assertions.
The response surface for the experimental design showed an optimal response of 0.6425 d-1 at the maximum levels of all the variables with marked decreases when the light intensity and carbon concentration are at low levels (-1). This is considering the Specific growth rate as response variable (figure 4), the analysis of main effects and their interactions.
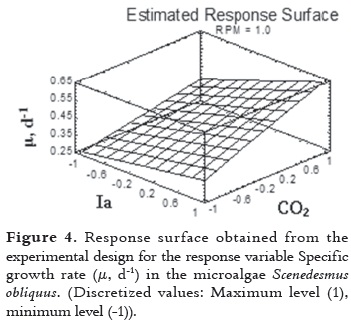
Exopolysaccharides concentration (Ych, mg L-1) as a response variable
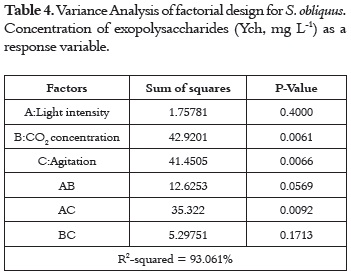
For the concentration of exopolysaccharides (Ych, mg L-1), the effects of concentration of carbon and agitation were significant because they present P values lower than 0.05 at the 95% confidence level. In addition, the light intensity-agitation (AC) interaction also had a significant effect (p < 0.05) (table 4).
The results obtained showed that the highest concentration of exopolysaccharides (24.7 mg L-1) was observed for the culture grown at 180 μE m-2s-1, 4% air/CO2, 1200 RPM and the lowest EPS concentration (8.8 mg L-1) was observed for the culture grown at 180 μE m-2s-1/ 0% air-CO2 %v/v/,0 rpm.
As observed in tables 3 and 4, the adjusted R2 of each model became larger than 0.9. This means that 90% of the variation of growth rate and EPS production under the effect of light intensity, carbon concentration and agitation can be explained by the developed models.
The results show that the main effects of CO2 concentration and agitation directly affect the production of EPS (figure 5). Thereby, indicates that EPS concentration increases within the carbon concentration range of 0 to 4%, and it reaches its maximum value (24.7 mg L-1) at 4%. Then, EPS concentration decreases progressively to reach 15.05 mg L-1 at 0% (% v/v air-CO2).
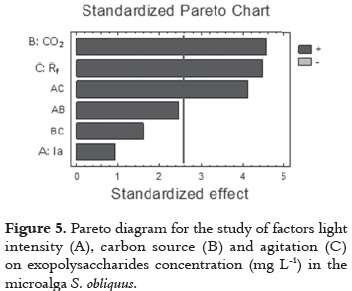
Another significant main effect on the EPS production was the agitation (p < 0.05), when this factor was fixed at the maximum level (1200 rpm), the highest EPS concentration (24.70 mgL-1) was found. However, a variation of this factor to the minimum level (0 rpm) generated a significant change in EPS production reaching a concentration of 8.80 mg L-1.
The effect of agitation (Rf) on the exopolysaccharides concentration - Ych (mg L-1) - shows a proportional relationship that can be associated to responses to the hydrodynamic stress caused by the impeller, which induces physiological and biochemical adaptations in the organism to reduce susceptibility to damage. These results on the overproduction of EPS could be associated to an environmental adaptation to reduce the shear through an increase in the viscosity of the medium (21). This observation is supported by the results obtained in this study, which show maximum production of EPS when the flow rate is more turbulent. It should be noted, however, that increased production of EPS may also be related to greater availability and better distribution of the energy source (light) and CO2. Additionally, results show that high yields of EPS are also associated with high levels of carbon in the medium and high levels of light on the cell.
By analyzing the effect of the Light intensity/ agitation interaction (AC) on the EPS concentration, we observed that the effect of agitation was strongly influenced by the effect of light intensity. Thus, at 1200 rpm and 80 μE m-2s-1(with 4% air- CO2) it was obtained value of EPS concentration of 16, 17 mg L-1, but this value improved to 24.70 mg L-1 when the light intensity increased to180 μE m-2s-1(with 4% air-CO2 and 1200 rpm).
The interaction between light intensity and carbon source, when they are kept at the maximal levels employed, also causes an increase in the production of biomass and EPS. This becomes evident considering that high availability of energy would be associated with increased generation of reducing power and ATP, which are the basic requirements for the reactions of the dark phase of photosynthesis. This would, in turn, increase the efficiency of CO2 fixation and, therefore, biomass and EPS productivity. Similar results have been found in strains of B. braunii (22), which exhibited the same effect of maximization, even under different adverse conditions.
The effect of light intensity has been reported in other studies (23) as one of the reasons for overproduction of EPS by microalgae. For practical purposes, the effect of illumination on the different responses must be considered. It can be observed that the incidence of light intensity (μE m-2s-1) on the production of EPS at its maximum level (180 μE m-2s-1) is significant when working with high light output, this is not the case when carbon availability in the medium is minimal (~0% CO2), in which case the trend in the response is negative.
This result is most likely associated with the physiological effects of adaptation for the reference microalgae, which prioritizes cell maintenance instead of synthesizing new cellular components when cells are in conditions of low carbon levels. As a result, there is a decrease in the concentration of EPS in the medium because the sugar stocks are required for other activities related to metabolic maintenance. This effect suggests that low availability of light is equivalent to low levels of carbon fixation. In addition, there is a lack of carbon in the medium, and all of these factors result in insufficient carbon supplies during the absorption phase of the carbon cycle (the dark phase of the process) to maintain the corresponding efficiency. In these conditions, the amount of fixed carbon would be minimal and would be consumed to support the development of the microalgae. Similar effects have been found in studies associated with the growth of microalgae under limiting conditions (24).
The response surface for the concentration of exopolysaccharides (Ych, mg L-1) (figure 6), confirms that the optimal levels for the production of exopolysaccharides are achieved at the maximum values of the three experimental variables considered.
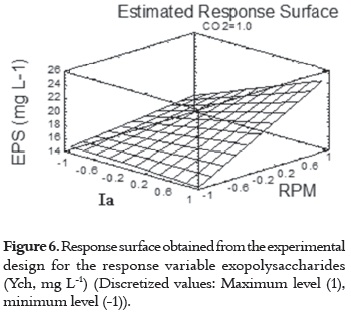
Therefore, when EPS (Ych, mg L-1) is used as the response variable, the optimal response of the microalgae Scenedesmus obliquus is expressed at an optimal value of 24.41 mg L-1. This value is exhibited when the experimental variables are all kept at the maximum levels tested. These observations are similar to the optimal response found for the specific growth rate.
From the experimental data and applying a factorial experimental design, a first-degree polynomial model can be established for the specific rate of growth and EPS as response variables. These polynomials are among the highest and lowest ranges considered for each of the experimental variables (light intensity, carbon concentration and agitation) and interactions between them. The equations obtained and their ratios are shown below for each output.
μ = 0.408+ 0.074* Ia + 0.0988*CO2+ 0.036*Ia*CO2 + 0.0163*Ia*Rf + 0.0063*CO2*Rf Equation 3.
Ych = 15.173+0.469*Ia + 2.316*CO2 + 2.276*Rf + 1.256*Ia*CO2 + 2.101*Ia*Rf + 0.814*CO2*Rf Equation 4.
where: μ = specific growth rate (d-1); Ych = concentration of exopolysaccharides in the supernatant (mg L-1); Ia = light intensity used for growing the photoautotrophic cultures (μE m-2 s-1); CO2= concentration of the carbon source supplied to the medium (air-CO2 mixture % v/v); Rf = shaking rate in the medium (rpm).
To verify the accuracy of the model obtained, a culture was carried out at the optimal levels (by triplicate), which corresponded to the experimental variables at their maximum levels, i.e. agitation (Rf) 1200 rpm, light intensity (Ia) 180μE m-2s-1, and carbon source (C*) 4% (a mixture of air enriched with CO2 in % v/v). For these assays, the specific growth rate (μ, d-1) and the concentration of exopolysaccharides‒EPS (Ych, mg L-1) were also used as response variables. Using these assays, we found that the specific growth rate, μ (d-1) exhibits a significant adjustment of its average for the assays, with a standard deviation (σ) of 0.005 and a virtually nil absolute difference (d-1) regarding to the optimum value of model. The exopolysaccharides concentration (mg L-1) showed a mean value less than projected; however, the average value of 23.79 mg L-1 is within the confidence interval (table 5).
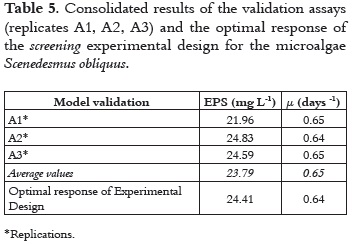
The specific growth rate (μ) and the concentration of exopolysaccharides (Ych), the two response variables under consideration, show that the process of carbon fixation that is associated with cell growth and the production of polymeric substances require the provision of an inorganic source of carbon. Therefore, the CO2 concentration can be considered to be the main factor affecting the productivity of the process. However, the CO2 supply should be considered in the context of the different interactions that affect its availability in the medium and the efficiency of its fixation in the photosynthetic process.
The increase in EPS can affect the viscosity of de culture medium and consequently the flow rate of the system. Therefore, in this research we carried out dynamic viscosity (mPa*s) measurements to determine the effect of EPS concentration in the viscosity culture medium. The increase in viscosity, associated with the production and concentration of EPS in the medium, is shown on figure 7.
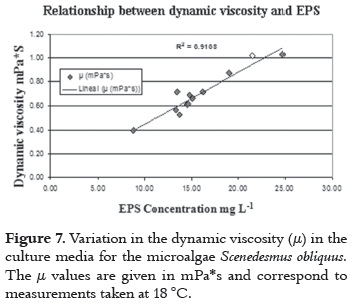
The increase in viscosity associated with the production and concentration of EPS in the medium is considered a physiological adaptation of microalgae to the environmental conditions of the production system. Specifically, this could be an adaptation against mechanical damage by trying to protect the cell wall by covering it in EPS. The results of Trujillo-Roldány et al., 2006 (25) in conditions of hydrodynamic stress in different biological models support this assertion.
Exopolysaccharides concentration in the culture medium could be related to the viscosity changes produced in it. There was an increase in the value of viscosity at the end of culture from 1.407 mPa s to 2.84 mPa s, which doubled the initial value. A direct relationship between viscosity of the culture medium and the concentration of EPS was found in the present investigation which is consistent with results by other authors (26).
CONCLUSIONS
This research showed that the model and the three-level -factorial design gives a reliable picture of the situation, and lets us know clear results for the decision making. The results indicate that the maximum productivities in biomass and exopolysaccharides production in the microalgae Scenedesmus obliquus are achieved under similar conditions, when each of the three experimental variables is at their maximum level. Thus, light intensity: 180 μE m-2s-1, carbon concentration 4% v/v, and agitation: 1200 rpm yielded a specific growth rate of 0.66 d-1 and a mean exopolysaccharide concentration of 24.1 mg L-1. The results reported in this investigation also showed that if the light intensity is at its maximum level and carbon concentration and agitation at them lowest level, the EPS productivity is negatively affected. Under these conditions it was observed that the EPS concentration was the lowest of all the experimental runs (8.8 mg L-1).
In the production process, when the experimental variables and their interactions are taken into account, the main phenomenon to consider is flow dynamics. The results of this study showed that this factor is critical to the production process because the flow rate affects the availability of light, which in turn affects the processes of carbon fixation and synthesis of macromolecules in microalgae, including the production of biomass and EPS. Therefore, new assays for the optimization of biomass and EPS production can be developed with a production system based on the optimal use of light associated with a variable shaking rate to improve the distribution of light in the culture medium, with a supply of CO2 in high concentration to maximize overall efficiency.
Studies on EPS production have been mainly conducted in Cyanobacteria, which reported a high accumulation of these substances in the culture medium. Investigations such as those by Yu et al., 2010 (7) in Nostoc flageliforme showed the presence of these substances at a concentration of 228.56 mg L-1 when the nutrient concentrations in the culture medium were modified. Studies in Arthrospira platensis, Trabelsi et al. (27) reported the presence of different concentrations of extracellular substances when changing the culture conditions such as temperature and light intensity. Although there are reports in green algae, evidence in Scenedesmus obliquus is not clearly reported.
With the results that we found in this study, we can conclude that the Scenedesmus obliquus is an appropriate microalga species for the production of exopolysaccharides (EPS). These exopolysaccharides have multiple uses, including utilization of soil improvers with the goal of developing innovative technological alternatives that are sustainable and that can present a response to this urgent need of the primary agricultural sector.
ACKNOWLEDGEMENTS
The authors acknowledge the technical and financial support provided by Colciencias, Asociación Colombiana para el Avance de la Ciencia-ACAC, Live Systems Technology S.A. and Fondo Patrimonial de la Universidad de La Sabana.
REFERENCES
1. Cohen Z, Richmond A (Editor). Handbook of microalgae mass culture. Florida: CRC Press; 1986. Chapter: Products from microalgae; p. 421-454. [ Links ]
2. Ördög V, Stirk WA, Lenobel R, Bancírová M, Strnad M et al. Screening microalgae for some potentially useful agricultural and pharmaceutical secondary metabolites. J Appl Phycol. 2004 Apr; 16 (4): 309-314. [ Links ]
3. Roeselers G, Van Loosdrecht MCM, Muyzer G. Phototrophic biofilms and their potential applications. J Appl Phycol. 2008 Jun; 20 (3): 227-235. [ Links ]
4. Cardozo KHM, Guaratini T, Barros MP, Falcao VR, Tonon AP et al. Metabolites from algae with economic impact. Comp Biochem Physiol. 2007 Jan-Feb; C146 (1-2): 60-78. [ Links ]
5. Spolaore P, Joanniss-Cassan C, Duran E, Isambert A. Commercial applications of microalgae. J Biosci Bioeng. 2006 Feb; 101 (2): 87-96. [ Links ]
6. Richert L, et al. Characterization of exopolysaccharides produced by Cyanobacteria isolated from Polynesian Microbial Mats. Curr Microbiol. 2005 Jun; 51 (6): 379-384. [ Links ]
7. Yu H, Jia S, Dai Y. Accumulation of exopolysaccharides in liquid suspension culture of Nostocflagelliforme cells. Appl Biochem Biotech. 2010 Feb; 160 (2): 552-560. [ Links ]
8. Donota F, Fontanaa A, Baccoua JC, Schorr-Galindoa S. Microbial exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction. Carbohyd Polym. 2011 Aug 31; 87 (2); 951-962. [ Links ]
9. Singh S, Arad S, Richmond A. Extracellular polysaccharide production in outdoor mass cultures of Porphyridium sp. in flat plate glass reactors. J Appl Phycol. 2000 Mar-May; 12 (3-5): 269-275. [ Links ]
10. Pulz O. Photobioreactors: production systems for phototrophic microorganisms. Appl Microbiol Biotech. 2001 Mar; 57 (3): 287-293. [ Links ]
11. Sánchez JF, Fernández JM, Acién FG, Rueda A, Pérez-Parra J, Molina E. Influence of culture conditions on the productivity and lutein content of the new strain Scenedesmus almeriensis. Proc Biochem. 2008 Apr; 43 (4): 398-405. [ Links ]
12. Martínez Sancho ME, Jiménez Castillo JM, El Yousfi F. Photoautotrophic consumption of phosphorus by Scenedesmus obliquus in a continuous culture-Influence of light intensity. Proc Biochem. 1999 Aug; 34 (8): 811-818. [ Links ]
13. Wood AM, Everroad RC, Wingard LM. Andersen RA (Editor). Algal Culturing Techniques. USA: Academic Press; 2005. Chapter: Measuring growth rates in microalgal cultures. p. 269-285. [ Links ]
14. Yemm EW, Willis JA. The estimation of carbohydrates in plant extracts by anthrone. J Biochem. 1954 Mar; 57 (3): 508-514. [ Links ]
15. Band CJ. Efecto de la composición bioquímica de microalgas sobre el valor nutritivo de dos cepas de Artemia [Tesis de Maestría]. [La Paz]: Instituto Politécnico Nacional: 1999. 69p. [ Links ]
16. Lewin RA. Extracellular polysaccharides of green algae. Can J Microbiol. 1956 Jul; 2 (7): 665-672. [ Links ]
17. Yu H, Jia S, Dai Y. Accumulation of Exopolysaccharides in liquid suspension culture of Nostoc flagelliforme cells. Appl Biochem Biotechnol. 2010 Feb; 160 (2): 552-560. [ Links ]
18. Wazer Van JR, Lyons JW, Kim KY, Colewell RE. Viscosity and flow measurements: a laboratory handbook of rheology. USA: John Wiley & Sons, Inc; 1963.406p. [ Links ]
19. Lee Y-K, Pirt SJ. Energetics of photosynthetic algal growth: influence of intermittent illumination in short (40 s) cycles. J Gen Microbiol. 1981 Jan; 124 (1): 926-935. [ Links ]
20. Wu X, Merchuk JC. A model integrating fluid dynamics in the photosynthesis and photoinhibition process. Chem Eng Sci. 2001 Nov; 56 (11): 3527-3538. [ Links ]
21. Märkl R, Bronnenmier R, Witterk B. The resistance of microorganism to hydrodynamic stress. Int Chem Eng. 1999 Feb; 31 (2): 185-197. [ Links ]
22. Dayananda C, Sarada R, Usha Rani M, Shamala TR, Ravishankar GA. Autotrophic cultivation of Botryococcus braunii for the production of hydrocarbons and exopolysaccharides in various media. Biomass Bioenerg. 2007 Jan; 31 (1): 87-93. [ Links ]
23. Rebolloso FM, García JL, Fernández JM, Acién-Fernández FG, Sánchez JA, Molina Grima E. Outdoor continuous culture of Porphyridium cruentum in a tubular photobioreactor: quantitative analysis of the daily cyclic variation of culture parameters. J. Biotechnol. 1999 Jan-Mar; 70(1-3 ): 271-288. [ Links ]
24. Grobbelaar JU. Turbulence in mass algal cultures and the role of light/dark fluctuations. J Appl Phycol. 1994 Mar; 6 (3): 331-335. [ Links ]
25. Trujillo-Roldán MA, Valdez-Cruz NA. El estrés hidrodinámico: Muerte y daño celular en cultivos agitados. Rev Latinoam Microbiol. 2006 Mar; 48 (3):269-280. [ Links ]
26. Singh S, Arad S, Richmond A. Extracellular polysaccharide production in outdoor mass cultures of Porphyridium sp. in flat plate glass reactors. J. Appl Phycol. 2000 Mar-May; 12 (3-5): 269-275. [ Links ]
27. Trabelsi L, Ben Ouada H, Hassen B, Ghoul MB. Combined effect of temperature and light intensity on growth and extracellular polymeric substance production by the cyanobacterium Arthrospiraplantensis. J Appl Phycol. 2009 Apr; 21 (4): 405-412. [ Links ]

