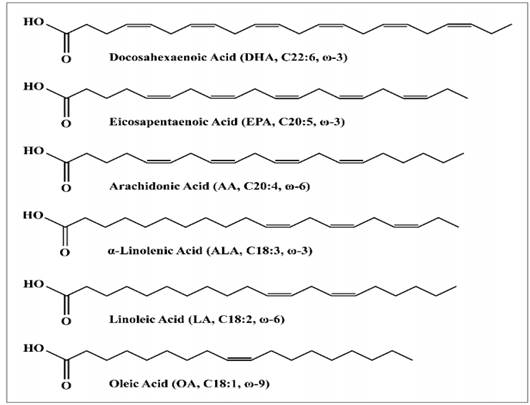
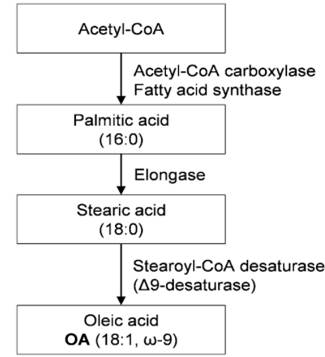
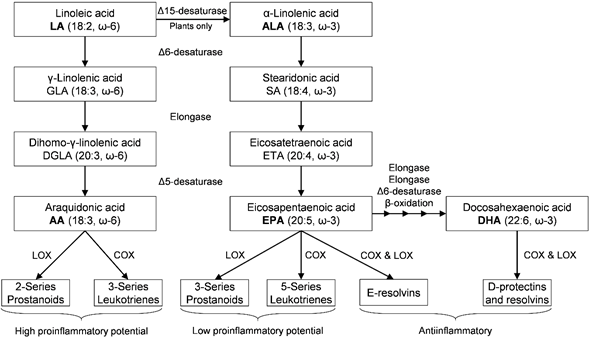
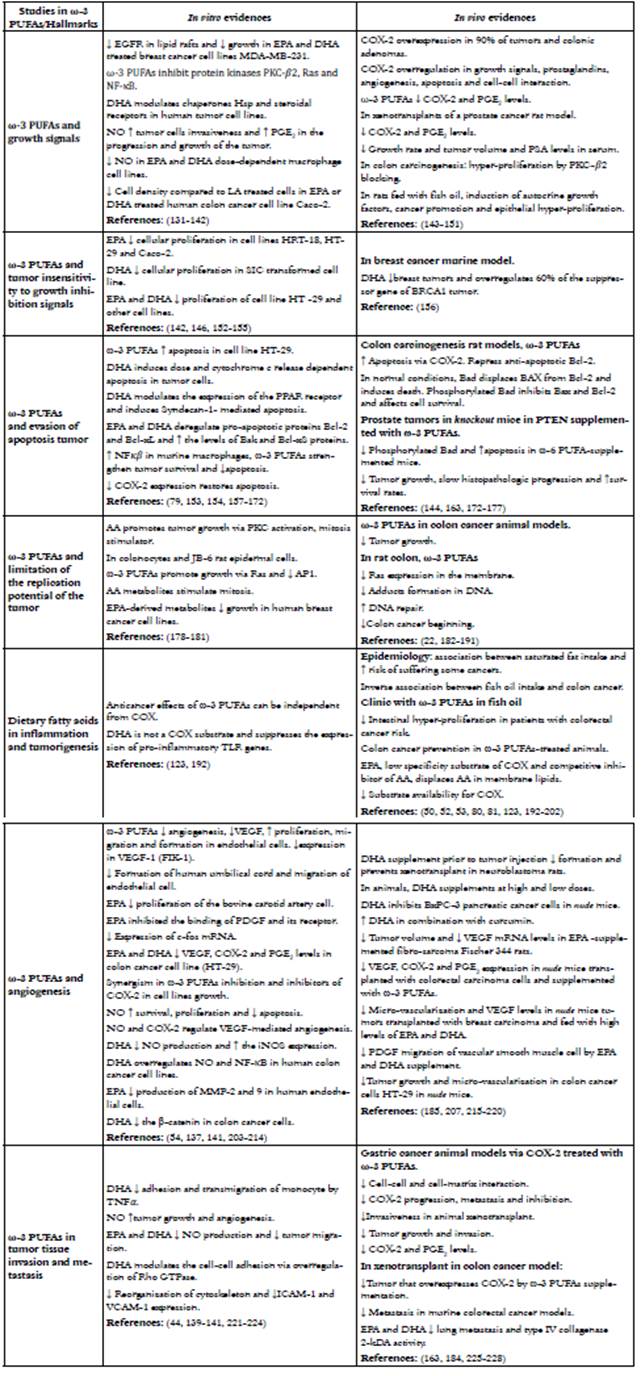
REFERENCES
1. Lee JY, Hwang DH. The modulation of inflammatory gene expression by lipids: mediation through Toll-like receptors. Mol Cells. 2006 Apr 30; 21(2):174-85.
[ Links ]
2. Lee JY, Sohn KH, Rhee SH, Hwang D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through toll-like receptor 4. J Biol Chem. 2001 May 18; 276(20):16683-9.
[ Links ]
3. Trichopoulou, Naska, Antoniou, Friel, Trygg, Turrini. Vegetable and fruit: the evidence in their favour and the public health perspective. Int J Vitam Nutr Res. 2003 Mar; 73(2):63-9.
[ Links ]
4. Chajes V, Thiebaut ACM, Rotival M, Gauthier E, Maillard V, Boutron-Ruault M-C, et al. Association between serum trans-monounsaturated fatty acids and breast cancer risk in the E3N-EPIC Study. Am J Epidemiol. 2008 Jun 1; 167(11):1312-20.
[ Links ]
5. Escrich E, Solanas M, Moral R, Grau L, Costa I, Vela E, et al. Lípidos de la dieta y cáncer de mama: evidencias científicas clínicas, anatomopatológicas y moleculares. Rev Esp Obes. 2008; 6(3):129-38.
[ Links ]
6. Martin-Moreno JM, Walter C, Gorgojo L, Banegas JR, Rodrlguez-Artalej F, Fernández-Rodríguez JC, et al. Dietary fat, olive oil intake and breast cancer risk. 1994; 780:774-80.
[ Links ]
7. Macquart-Moulin G, Riboli E, Cornée J, Charnay B, Berthezene P, Day N. Case-control study on colorectal cancer and diet in marseilles. Int J Cancer. 1986 Aug 15; 38(2):183-91.
[ Links ]
8. Stoneham M, Goldacre M, Seagroatt V, Gill L. Olive oil, diet and colorectal cancer: an ecological study and a hypothesis. J Epidemiol Community Health. 2000; 54 (10):756-60.
[ Links ]
9. Bautista D, Obrador A, Moreno V, Bautista D, Obrador A, Moreno V, et al. Ki-ras mutation modifies the protective effect of dietary monounsaturated fat and calcium on sporadic colorectal cancer. Cancer Epidemiol Biomarkers Prev. 1997; 6 (January):57-61.
[ Links ]
10. Bartoli R. Effect of olive oil on early and late events of colon carcinogenesis in rats: modulation of arachidonic acid metabolism and local prostaglandin E2 synthesis. Gut. 2000; 46(2):191-9.
[ Links ]
11. Reddy BS, Maeura Y. Tumor promotion by dietary fat in azoxymethane-induced colon carcinogenesis in female F344 rats: influence of amount and source of dietary fat. J Natl Cancer Inst. 1984; 72(3):745-50.
[ Links ]
12. Schwartz B, Birk Y, Raz A, Madar Z. Nutritional-pharmacological combinations. Eur J Nutr. 2004; 43(4):221-9.
[ Links ]
13. Navarro-Tito N, Soto-Guzmán A, Castro-Sánchez L, Martínez-Orozco R, Salazar EP. Oleic acid promotes migration on MDA-MB-231 breast cancer cells through an arachidonic acid-dependent pathway. Int J Biochem Cell Biol. 2010; 42(2):306-17.
[ Links ]
14. Nakamura MT, Nara TY. Structure, function, and dietary regulation of delta6, delta5, and delta9 desaturases. Annu Rev Nutr. 2004 Jul 14; 24(1):345-76.
[ Links ]
15. Johnson GB, Brunn GJ, Platt JL. Activation of mammalian Toll-like receptors by endogenous agonists. Crit Rev Immunol. 2003; 23(1-2):15-44.
[ Links ]
16. Xu XH, Shah PK, Faure E, Equils O, Thomas L, Fishbein MC, et al. Toll-Like receptor-4 is expressed by macrophages in murine and human lipid-rich atherosclerotic plaques and upregulated by oxidized LDL. Circulation. 2001 Dec 18; 104(25):3103-8.
[ Links ]
17. Belvin MP, Anderson K V. A conserved signaling pathway: the Drosophila Toll-dorsal pathway. Annu Rev Cell Dev Biol. 1996 Nov; 12(1):393-416.
[ Links ]
18. Hoffmann J a, Kafatos FC, Janeway C a, Ezekowitz R a. Phylogenetic perspectives in innate immunity. Science. 1999; 284(5418):1313-8.
[ Links ]
19. Lee JY, Zhao L, Hwang DH. Modulation of pattern recognition receptor-mediated inflammation and risk of chronic diseases by dietary fatty acids. 2009; 68(1):38-61.
[ Links ]
20. Pollard JW. Opinion: tumour-educated macrophages promote tumour progression and metastasis. Nat Rev Cancer. 2004 Jan; 4(1):71-8.
[ Links ]
21. Coussens LM, Werb Z. Inflammation and cancer. Nature 2002; 420(6917):860-7.
[ Links ]
22. Reddy BS, Maruyama H. Effect of dietary fish oil on azoxymethane-induced colon carcinogenesis in male F344 rats. Cancer Res. 1986; 46(7):3367-70.
[ Links ]
23. Reddy BS, Sugie S. Effect of different levels of omega-3 and omega-6 fatty acids on azoxymethane-induced colon carcinogenesis in F344 rats. Cancer Res. 1988; 48(23): 6642-7.
[ Links ]
24. Anand P, Sundaram C, Jhurani S, Kunnumakkara AB, Aggarwal BB. Curcumin and cancer: an "old-age" disease with an "age-old" solution. Cancer Lett. 2008; 267(1): 133-64.
[ Links ]
25. Wong SW, Kwon MJ, Choi AMK, Kim HP, Nakahira K, Hwang DH. Fatty acids modulate Toll-like receptor 4 activation through regulation of receptor dimerization and recruitment into lipid rafts in a reactive oxygen species-dependent manner. J Biol Chem. 2009; 284(40):27384-92.
[ Links ]
26. Jarnagin WR, Rockey DC, Koteliansky VE, Wang SS, Montgomery Bissell D. Expression of variant fibronectins in wound healing: cellular source and biological activity of the EIIIA segment in rat hepatic fibrogenesis. J Cell Biol. 1994; 127(6 II): 2037-48.
[ Links ]
27. Saito S, Yamaji N, Yasunaga K, Saito T, Matsumoto SI, Katoh M, et al. The fibronectin extra domain A activates matrix metalloproteinase gene expression by an interleukin-1-dependent mechanism. J Biol Chem. 1999; 274(43):30756-63.
[ Links ]
28. Kato M, Wang H, Kainulainen V, Fitzgerald ML, Ledbetter S, Ornitz DM, et al. Physiological degradation converts the soluble syndecan-1 ectodomain from an inhibitor to a potent activator of FGF-2. Nat Med. 1998 Jun; 4(6):691-7.
[ Links ]
29. Platt JL, Dalmasso AP, Lindman BJ, Ihrcke NS, Bach FH. The role of C5a and antibody in the release of heparan sulfate from endothelial cells. Eur J Immunol. 1991; 21(11):2887-90.
[ Links ]
30. Subramanian N, Vasudevan S, Hussain AT. Direct communication of a pulmonary artery with left atrium--report of a case with balloon occlusion studies and successful correction: a case report. Angiology. 1986 Feb; 37(2):136-41.
[ Links ]
31. Hold GL, Rabkin CS, Chow W, Smith MG, Gammon MD, Risch HA, et al. A functional polymorphism of Toll-Like receptor 4 gene increases risk of gastric carcinoma and its precursors. Gastroenterology. 2007; 132(3):905-12.
[ Links ]
32. Sun J, Wiklund F, Zheng SL, Chang B, Bälter K, Li L, et al. Sequence variants in Toll-like receptor gene cluster (TLR6-TLR1-TLR10) and prostate cancer risk. J Natl Cancer Inst. 2005; 97(7):525-32.
[ Links ]
33. Tahara T, Arisawa T, Wang F, Shibata T, Nakamura M, Sakata M, et al. Toll-like receptor 2 - 196 to 174del polymorphism influences the susceptibility of Japanese people to gastric cancer. Cancer Sci. 2007; 98(11):1790-4.
[ Links ]
34. Boraska Jelavic T, Barisic M, Drmic Hofman I, Boraska V, Vrdoljak E, Peruzovic M, et al. Microsatelite GT polymorphism in the Toll-like receptor 2 is associated with colorectal cancer. Clin Genet. 2006; 70(2):156-60.
[ Links ]
35. Nieters A, Beckmann L, Deeg E, Becker N. Gene polymorphisms in Toll-like receptors, interleukin-10, and interleukin-10 receptor alpha and lymphoma risk. Genes Immun. 2006; 7(8):615-24.
[ Links ]
36. Anderson BM, Ma DWL. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis. 2009; 8(1):33.
[ Links ]
37. Su HM, Moser AB, Moser HW, Watkins PA. Peroxisomal straight-chain acyl-CoA oxidase and D-bifunctional protein are essential for the retroconversion step in docosahexaenoic acid synthesis. J Biol Chem. 2001; 276(41):38115-20.
[ Links ]
38. Hussein N, Ah-Sing E, Wilkinson P, Leach C, Griffin BA, Millward DJ. Long-chain conversion of [13C] linoleic acid and a-linolenic acid in response to marked changes in their dietary intake in men. J Lipid Res. 2005; 46(2):269-80.
[ Links ]
39. Weylandt KH, Chiu C, Gomolka B, Waechter SF, Wiedenmann B. Prostaglandins and other lipid mediators omega-3 fatty acids and their lipid mediators?: towards an understanding of resolvin and protectin formation omega-3 fatty acids and their resolvin / protectin mediators. Prostaglandins Other Lipid Mediat. 2012 Mar; 97(3-4):73-82.
[ Links ]
40. Weylandt KH, Kang JX. Rethinking lipid mediators. Lancet. 2005; 366(9486):618-20.
[ Links ]
41. Lukiw WJ, Cui J, Marcheselli VL, Bodker M, Botkjaer A, Gotlinger K, et al. A role for docosahexaenoic acid - derived neuroprotectin D1 in neural cell survival and Alzheimer disease. J Clin Invest. 2005; 115(10):2774-83.
[ Links ]
42. Mukherjee PK, Marcheselli VL, de Rivero Vaccari JC, Gordon WC, Jackson FE, Bazan NG. Photoreceptor outer segment phagocytosis attenuates oxidative stress-induced apoptosis with concomitant neuroprotectin D1 synthesis. Proc Natl Acad Sci U S A. 2007; 104(32):13158-63.
[ Links ]
43. Kang JX, Wang J, Wu L, Kang ZB. Transgenic mice: Fat-1 mice convert n-6 to n-3 fatty acids. Nature. 2004 Feb 5; 427(6974):504.
[ Links ]
44. Ratnayake WMN, Galli C. Fat and fatty acid terminology, methods of analysis and fat digestion and metabolism: a background review paper. Ann Nutr Metab. 2009; 55(1-3):8-43.
[ Links ]
45. Burlingame B, Nishida C, Uauy R, Weisell R. Fats and fatty acids in human nutrition: introduction. Ann Nutr Metab. 2009; 55(1-3):5-7.
[ Links ]
46. Calder PC. Fatty acids and inflammation: the cutting edge between food and pharma. Eur J Pharmacol. 2011 Sep; 668:S50-8.
[ Links ]
47. Stephenson JA, Arshad A, Morgan B, Metcalfe MS, Dennison AR, Infirmary LR, et al. The multifaceted effects of omega-3 polyunsaturated fatty acids on the hallmarks of cancer. J Lipids. 2013; 2013:261247. doi: 10.1155/2013/261247. Epub 2013 May 16.
[ Links ]
48. Dommels YEM, Haring MMG, Keestra NGM, Alink GM, van Bladeren PJ, van Ommen B. The role of cyclooxygenase in n-6 and n-3 polyunsaturated fatty acid mediated effects on cell proliferation, PGE(2) synthesis and cytotoxicity in human colorectal carcinoma cell lines. Carcinogenesis. 2003 Mar; 24(3):385-92.
[ Links ]
49. Mason JK, Fu M, Chen J, Thompson LU. Flaxseed oil enhances the effectiveness of trastuzumab in reducing the growth of HER2-overexpressing human breast tumors (BT-474). J Nutr Biochem. 2015; 26(1):16-23.
[ Links ]
50. Ward JM, Devor-Henneman DE. Mouse models of human familial cancer syndromes. Toxicol Pathol. 2004; 32(SUPPL. 1):90-8.
[ Links ]
51. Maddison K, Clarke AR. New approaches for modelling cancer mechanisms in the mouse. J Pathol. 2005; 205(2):181-93.
[ Links ]
52. Zock PL, Katan MB. Linoleic acid intake and cancer risk: a review and meta-analysis. Am J Clin Nutr. 1998 Jul; 68(1):142-53.
[ Links ]
53. Maclean CH, Newberry SJ, Mojica WA, Khanna P, Issa AM, Suttorp MJ, et al. Effects of omega-3 fatty acids on cancer risk: a systematic review. Am Med Assoc. 2006; 295(4):403-16.
[ Links ]
54. Calviello G, Di Nicuolo F, Gragnoli S, Piccioni E, Serini S, Maggiano N, et al. n-3 PUFAs reduce VEGF expression in human colon cancer cells modulating the COX-2/PGE 2 induced ERK-1 and -2 and HIF-1a induction pathway. Carcinogenesis. 2004 Dec 1; 25(12):2303-10.
[ Links ]
55. Ibrahim A, Mbodji K, Hassan A, Aziz M, Boukhettala N, Coëffier M, et al. Anti-inflammatory and anti-angiogenic effect of long chain n-3 polyunsaturated fatty acids in intestinal microvascular endothelium. Clin Nutr. 2011; 30(5):678-87.
[ Links ]
56. Storey A, McArdle F, Friedmann PS, Jackson MJ, Rhodes LE. Eicosapentaenoic acid and docosahexaenoic acid reduce UVB- and TNF-a-induced IL-8 secretion in keratinocytes and UVB-induced IL-8 in fibroblasts. J Invest Dermatol. 2005; 124(1): 248-55.
[ Links ]
57. Dolcet X, Llobet D, Pallares J, Matias-Guiu X. NF-kB in development and progression of human cancer. Virchows Arch. 2005; 446(5):475-82.
[ Links ]
58. Griffitts J, Saunders D, Tesiram YA, Reid GE, Salih A, Liu S, et al. Non-mammalian fat-1 gene prevents neoplasia when introduced to a mouse hepatocarcinogenesis model. Omega-3 fatty acids prevent liver neoplasia. Biochim Biophys Acta - Mol Cell Biol Lipids. 2010; 1801(10):1133-44.
[ Links ]
59. Zucker S, Lysik RM, Zarrabi MH, Moll U. M(r) 92,000 type IV collagenase is increased in plasma of patients with colon cancer and breast cancer. Cancer Res. 1993 Jan 1; 53(1):140-6.
[ Links ]
60. Cao W, Ma Z, Rasenick MM, Yeh S, Yu J. n-3 poly-unsaturated fatty acids shift estrogen signaling to inhibit human breast cancer cell growth. PLoS One. 2012; 7(12):1-11.
[ Links ]
61. Rogers KR, Kikawa KD, Mouradian M, Hernández K, McKinnon KM, Ahwah SM, et al. Docosahexaenoic acid alters epidermal growth factor receptor-related signaling by disrupting its lipid raft association. Carcinogenesis. 2010; 31(9):1523-30.
[ Links ]
62. Sharma A, Belna J, Espat J, Rodríguez G, Cannon VT, Hurteau JA. Effects of omega-3 fatty acids on components of the transforming growth factor beta-1 pathway: implication for dietary modification and prevention in ovarian cancer. Am J Obstet Gynecol. 2009; 200(5):516.e1-516.e6.
[ Links ]
63. Siriwardhana N, Kalupahana NS, Moustaid-Moussa N. Health Benefits of n-3 Polyunsaturated Fatty Acids: Eicosapentaenoic Acid and Docosahexaenoic Acid. Ad Food Nutr Res. 2012; 65: 211-22.
[ Links ]
64. D'Orazio N, Gammone MA, Gemello E, De Girolamo M, Cusenza S, Riccioni G. Marine bioactives: pharmacological properties and potential applications against inflammatory diseases. Mar Drugs. 2012; 10(4):812-33.
[ Links ]
65. Serini S, Fasano E, Piccioni E, Cittadini ARM, Calviello G. Differential anti-cancer effects of purified EPA and DHA and possible mechanisms involved. 2011; 4065-75.
[ Links ]
66. Serini S, Fasano E, Piccioni E, Monego G, Cittadini ARM, Ranelletti FO, et al. DHA induces apoptosis and differentiation in human melanoma cells in vitro: involvement of HuR-mediated COX-2 mRNA stabilization and b-catenin nuclear translocation. 2012; 33(1):164-73.
[ Links ]
67. Serini S, Donato V, Piccioni E, Trombino S, Monego G, Toesca A, et al. Docosahexaenoic acid reverts resistance to UV-induced apoptosis in human keratinocytes: involvement of COX-2 and HuR. J Nutr Biochem. 2011; 22(9):874-85.
[ Links ]
68. Fasano E, Serini S, Piccioni E, Toesca A, Monego G, Cittadini AR, et al. DHA induces apoptosis by altering the expression and cellular location of GRP78 in colon cancer cell lines. Biochim Biophys Acta - Mol Basis Dis. 2012; 1822(11):1762-72.
[ Links ]
69. Calviello G, Resci F, Serini S, Piccioni E, Toesca A, Boninsegna A, et al. Docosahexaenoic acid induces proteasome-dependent degradation of ß-catenin, down-regulation of survivin and apoptosis in human colorectal cancer cells not expressing COX-2. Carcinogenesis. 2007; 28(6):1202-9.
[ Links ]
70. Saw CLL, Huang Y, Kong AN. Synergistic anti-inflammatory effects of low doses of curcumin in combination with polyunsaturated fatty acids: docosahexaenoic acid or eicosapentaenoic acid. Biochem Pharmacol. 2010; 79(3):421-30.
[ Links ]
71. Finocchiaro C, Segre O, Fadda M, Monge T, Scigliano M, Schena M, et al. Effect of n-3 fatty acids on patients with advanced lung cancer: a double-blind, placebo-controlled study. Br J Nutr. 2012; 108(02):327-33.
[ Links ]
72. Zhu G, Ou Q, Zhang T, Jiang X, Sun G, Zhang N, et al. A more desirable balanced polyunsaturated fatty acid composition achieved by heterologous expression of? ?15/ ?4 desaturases in mammalian cells. PLoS One. 2013; 8(12):8-13.
[ Links ]
73. Read JA, Beale PJ, Volker DH, Smith N, Childs A, Clarke SJ. Nutrition intervention using an eicosapentaenoic acid (EPA)-containing supplement in patients with advanced colorectal cancer. Effects on nutritional and inflammatory status: a phase II trial. Support Care Cancer. 2007; 15(3):301-7.
[ Links ]
74. Lou YR, Peng QY, Li T, Medvecky CM, Lin Y, Shih WJ, et al. Effects of high-fat diets rich in either omega-3 or omega-6 fatty acids on UVB-induced skin carcinogenesis in SKH-1 mice. Carcinogenesis. 2011; 32(7):1078-84.
[ Links ]
75. Park KS, Lim JW, Kim H. Inhibitory mechanism of omega-3 fatty acids in pancreatic inflammation and apoptosis. Ann N Y Acad Sci. 2009 Aug; 1171:421-7.
[ Links ]
76. Rosa DD, Loureno FC, Da Fonseca ACMH, De Sales RL, Ribeiro SMHR, Neves CA, et al. Fish oil improves the lipid profile and reduces inflammatory cytokines in wistar rats with precancerous colon lesions. Nutr Cancer. 2012; 64(4):569-79.
[ Links ]
77. Serini S, Piccioni E, Calviello G. ?-3 PUFAs and colon cancer: experimental studies and human interventional trials. In: Dietary Omega-3 Polyunsaturated Fatty Acids and Cancer. Springer USA. Calviello G and Serini S Editors. 2010. 67-89p.
[ Links ]
78. Lucas M, Dewailly E, Muckle G, Ayotte P, Bruneau S, Gingras S, et al. Gestational age and birth weight in relation to n-3 fatty acids among Inuit (Canada). Lipids. 2004; 39(7):617-26.
[ Links ]
79. Gleissman H, Johnsen JI, Kogner P. Omega-3 fatty acids in cancer, the protectors of good and the killers of evil? Exp Cell Res. 2010; 316(8):1365-73.
[ Links ]
80. Lanier AP, Holck P, Ehrsam Day G, Key C. Childhood cancer among Alaska natives. Pediatrics. 2003; 112(5):e396.
[ Links ]
81. Chen YQ, Berquin IM, Daniel LW, Edwards IJ, O'Flaherty JT, Thomas MJ, et al. Omega-3 Fatty Acids and Cancer Risk. JAMA. 2006 Jul 19; 296(3):278.
[ Links ]
82. Anti M, Marra G, Armelao F, Bartoli GM, Ficarelli R, Percesepe A, et al. Effect of omega-3 fatty acids on rectal mucosal cell proliferation in subjects at risk for colon cancer. Gastroenterology. 1992; 103(1):883-91.
[ Links ]
83. Huang YC, Jessup JM, Forse RA, Flickner S, Pleskow D, Anastopoulos HT, et al. n-3 fatty acids decrease colonic epithelial cell proliferation in high-risk bowel mucosa. Lipids. 1996 Mar; 31 Suppl: S313-7.
[ Links ]
84. West NJ, Clark SK, Phillips RK, Hutchinson JM, Leicester RJ, Belluzzi A, et al. Eicosapentaenoic acid reduces rectal polyp number and size in familial adenomatous polyposis. Gut. 2010; 59(7):918-25.
[ Links ]
85. Ramchurren N, Karmali R. Effects of gamma-linolenic and dihomo-gamma-linolenic acids on 7,12-dimethylbenz(a)anthracene-induced mammary tumors in rats. Prostaglandins Leukot Essent Fatty Acids. 1995 Aug; 53(2):95-101.
[ Links ]
86. Sauer LA, Blask DE, Dauchy RT. Dietary factors and growth and metabolism in experimental tumors. J Nutr Biochem. 2007; 18(10):637-49.
[ Links ]
87. Welsch CW. Relationship between dietary fat and experimental mammary tumorigenesis: a review and critique. Cancer Res. 1992; 52 (7 Supplement):2040s-2048s.
[ Links ]
88. Mouradian M, Kikawa KD, Johnson ED, Beck KL, Pardini RS. Key roles for GRB2-associated-binding protein 1, phosphatidylinositol-3-kinase, cyclooxygenase 2, prostaglandin E2 and transforming growth factor alpha in linoleic acid-induced upregulation of lung and breast cancer cell growth. Prostaglandins Leukot Essent Fat Acids. 2014; 90(4):105-15.
[ Links ]
89. Hudson JA, Cai Y, Corner RJ, Morvan B, Joblin KN. Identification and enumeration of oleic acid and linoleic acid hydrating bacteria in the rumen of sheep and cows. J Appl Microbiol. 2000; 88(2):286-92.
[ Links ]
90. Connolly JM, Coleman M, Rose DP. Effects of dietary fatty acids on DU145 human prostate cancer cell growth in athymic nude mice. Nutr Cancer. 1997; 29(2):114-9
[ Links ]
91. Kepler CR, Hirons KP, McNeill JJ, Tove SB. Intermediates and products of the biohydrogenation of linoleic acid by Butyrinvibrio fibrisolvens. J Biol Chem. 1966 Mar 25; 241(6):1350-4.
[ Links ]
92. Ewaschuk JB, Walker JW, Diaz H, Madsen KL. Bioproduction of conjugated linoleic acid by probiotic bacteria occurs in vitro and in vivo in mice. J Nutr. 2006; 136(6):1483-7.
[ Links ]
93. Kelley NS, Hubbard NE, Erickson KL. Conjugated linoleic acid isomers and cancer. J Nutr. 2007; 137(12):2599-607.
[ Links ]
94. Chujo H, Yamasaki M, Nou S, Koyanagi N, Tachibana H, Yamada K. Effect of conjugated linoleic acid isomers on growth factor-induced proliferation of human breast cancer cells. Cancer Lett. 2003; 202(1):81-7.
[ Links ]
95. Tanmahasamut P, Liu J, Hendry LB, Sidell N. Conjugated linoleic acid blocks estrogen signaling in human breast cancer cells. J Nutr. 2004; 134(3):674-80.
[ Links ]
96. De la Torre A, Debiton E, Durand D, Chardigny J-M, Berdeaux O, Loreau O, et al. Conjugated linoleic acid isomers and their conjugated derivatives inhibit growth of human cancer cell lines. Anticancer Res. 2005; 25(6B):3943-9.
[ Links ]
97. Beppu F, Hosokawa M, Tanaka L, Kohno H, Tanaka T, Miyashita K. Potent inhibitory effect of trans9, trans11 isomer of conjugated linoleic acid on the growth of human colon cancer cells. J Nutr Biochem. 2006;17(12):830-6.
[ Links ]
98. Palombo JD, Ganguly A, Bistrian BR, Menard MP. The antiproliferative effects of biologically active isomers of conjugated linoleic acid on human colorectal and prostatic cancer cells. Cancer Lett. 2002; 177(2):163-72.
[ Links ]
99. Kim EJ, Shin H-K, Cho JS, Lee SK, Won MH, Kim JW, et al. trans-10, cis-12 conjugated linoleic acid inhibits the G1-S cell cycle progression in DU145 human prostate carcinoma cells. J Med Food. 2006; 9(3):293-9.
[ Links ]
100. Ochoa JJ, Farquharson AJ, Grant I, Moffat LE, Heys SD, Wahle KWJ. Conjugated linoleic acids (CLAs) decrease prostate cancer cell proliferation: different molecular mechanisms for cis-9, trans-11, and trans-10 cis-12 isomers. Carcinogenesis. 2004; 25(7):1185-91.
[ Links ]
101. Liu JR, Chen BQ, Yang YM, Wang XL, Xue Y Ben, Zheng YM, et al. Effect of apoptosis on gastric adenocarcinoma cell line SGC-7901 induced by cis-9, trans-11-conjugated linoleic acid. World J Gastroenterol. 2002; 8(6):999-1004.
[ Links ]
102. Horrobin DF, Ziboh VA. The importance of linoleic acid metabolites in cancer metastasis and in the synthesis and actions of 13-HODE. Adv Exp Med Biol. 1997; 433:291-4.
[ Links ]
103. Abou-el-Ela SH, Prasse KW, Carroll R, Wade AE, Dharwadkar S, Bunce OR. Eicosanoid synthesis in 7,12-dimethylbenz(a)anthracene-induced mammary carcinomas in Sprague-Dawley rats fed primrose oil, menhaden oil or corn oil diet. Lipids. 1988 Oct; 23(10):948-54.
[ Links ]
104. Kitagawa H, Noguchi M. Comparative effects of piroxicam and esculetin on incidence, proliferation, and cell kinetics of mammary carcinomas induced by 7,12-dimethylbenz[a]anthracene in rats on high- and low-fat diets. Oncology. 1994; 51(5): 401-10.
[ Links ]
105. Wymann MP, Schneiter R. Lipid signalling in disease. Nat Rev Mol Cell Biol. 2008; 9(2):162-76.
[ Links ]
106. Hillyard LA, Abraham S. Effect of dietary polyunsaturated fatty acids on growth of mammary adenocarcinomas in mice and rats. Cancer Res. 1979 Nov; 39(11):4430-7.
[ Links ]
107. Li Y, Zhao H, Wang Y, Zheng H, Yu W, Chai H, et al. Isoliquiritigenin induces growth inhibition and apoptosis through downregulating arachidonic acid metabolic network and the deactivation of PI3K/Akt in human breast cancer. Toxicol Appl Pharmacol. 2013; 272(1):37-48.
[ Links ]
108. Pender-Cudlip MC, Krag KJ, Martini D, Yu J, Guidi A, Skinner SS, et al. Delta-6-desaturase activity and arachidonic acid synthesis are increased in human breast cancer tissue. Cancer Sci. 2013; 104(6):760-4.
[ Links ]
109. Sakai M, Kakutani S, Horikawa C, Tokuda H, Kawashima H, Shibata H, et al. Arachidonic acid and cancer risk: a systematic review of observational studies. BMC Cancer. 2012; 12(1):1-27.
[ Links ]
110. Yang P, Cartwright CA, Li J, Wen S, Prokhorova IN, Shureiqi I, et al. Arachidonic acid metabolism in human prostate cancer. Int J Oncol. 2012; 41(4):1495-503.
[ Links ]
111. Amirian ES, Ittmann MM, Scheurer ME. Associations between arachidonic acid metabolism gene polymorphisms and prostate cancer risk. Prostate. 2011 Sep 15; 71(13):1382-9.
[ Links ]
112. Fujiwara F, Todo S, Imashuku S. Antitumor effect of gamma-linolenic acid on cultured human neuroblastoma cells. Prostaglandins Leukot Med. 1986 Aug; 23(2-3):311-20.
[ Links ]
113. Sagar PS, Das UN. Cytotoxic action of cis-unsaturated fatty acids on human cervical carcinoma (HeLa) cells in vitro. Prostaglandins, Leukot Essent Fat Acids. 1995; 53(4):287-99.
[ Links ]
114. Das UN. Tumoricidal action of cis-unsaturated fatty acids and their relationship to free radicals and lipid peroxidation. Cancer Lett. 1991. Mar 1; 56(3):235-43.
[ Links ]
115. Bégin ME, Das UN, Ells G, Horrobin DF. Selective killing of human cancer cells by polyunsaturated fatty acids. Prostaglandins, Leukot Med [Internet]. 1985 Aug 1 [cited 2017 Nov 1]; 19(2):177-86.
[ Links ]
116. Bégin ME, Ells G, Das UN, Horrobin DF. Differential killing of human carcinoma cells supplemented with n-3 and n-6 polyunsaturated fatty acids. JNCI J Natl Cancer Inst. 1986 Nov 1; 77(5):1053-62.
[ Links ]
117. Leaver H a, Bell HS, Rizzo MT, Ironside JW, Gregor A, Wharton SB, et al. Antitumour and pro-apoptotic actions of highly unsaturated fatty acids in glioma. Prostaglandins Leukot Essent Fatty Acids. 2002; 66:19-29.
[ Links ]
118. Sangeetha Sagar P, Das UN, Koratkar R, Ramesh G, Padma M, Sravan Kumar G. Cytotoxic action of cis-unsaturated fatty acids on human cervical carcinoma (HeLa) cells: relationship to free radicals and lipid peroxidation and its modulation by calmodulin antagonists. Cancer Lett. 1992; 63(3):189-98.
[ Links ]
119. Vartak S, McCaw R, Davis CS, Robbins ME, Spector AA. Gamma-linolenic acid (GLA) is cytotoxic to 36B10 malignant rat astrocytoma cells but not to "normal" rat astrocytes. Br J Cancer. 1998; 77(10):1612-20.
[ Links ]
120. Colquhoun A. Gamma-linolenic acid alters the composition of mitochondrial membrane subfractions, decreases outer mitochondrial membrane binding of hexokinase and alters carnitine palmitoyltransferase I properties in the Walker 256 rat tumour. Biochim Biophys Acta - Mol Cell Biol Lipids. 2002; 1583(1):74-84.
[ Links ]
121. Das UN. Essential fatty acids and their metabolites and cancer. Nutrition. 1999 Mar; 15(3):239-40.
[ Links ]
122. Das UN. Tumoricidal action of cis-unsaturated fatty acids and their relationship to free radicals and lipid peroxidation. Cancer Lett. 1991 Mar 1;56 (3):235-43.
[ Links ]
123. Abou-el-ela SH, Prasse KW, Farrell RL, Prasse K V, Carroll RW, Wade AE, et al. Effects of D, L-2-difluoromethylornithine and indomethacin on mammary tumor promotion in rats fed high n-3 and/or n-6 fat diets. Cancer Research 49(6):1434-40
[ Links ]
124. Yamasaki M, Chujo H, Koga Y, Oishi A, Rikimaru T, Shimada M, et al. Potent cytotoxic effect of the trans10, cis12 isomer of conjugated linoleic acid on rat hepatoma dRLh-84 cells. Cancer Lett. 2002; 188(1-2):171-80.
[ Links ]
125. Santen RJ, Lobenhofer EK, Afshari CA, Bao Y, Song RX. Adaptation of estrogen-regulated genes in long-term estradiol deprived MCF-7 breast cancer cells. Breast Cancer Res Treat. 2005; 94(3):213-23.
[ Links ]
126. Cho HJ, Kim WK, Jung JI, Kim EJ, Lim SS, Kwon DY, et al. Trans-10,cis-12, not cis-9,trans-11, conjugated linoleic acid decreases ErbB3 expression in HT-29 human colon cancer cells. World J Gastroenterol. 2005; 11(33):5142-50.
[ Links ]
127. Das UN. Gamma-linolenic acid therapy of human glioma-a review of in vitro, in vivo, and clinical studies. Med Sci Monit. 2007; 13(7):RA119-31.
[ Links ]
128. Shiau AK, Barstad D, Loria PM, Cheng L, Kushner PJ, Agard DA, et al. The structural basis of estrogen receptor/coactivator recognition and the antagonism of this interaction by tamoxifen. Cell. 1998; 95(7):927-37.
[ Links ]
129. Kelley NS, Hubbard NE, Erickson KL. Conjugated linoleic acid isomers and cancer. J Nutr. 2007; 137(12):2599-607.
[ Links ]
130. Colquhoun A, Schumacher RI. ?-Linolenic acid and eicosapentaenoic acid induce modifications in mitochondrial metabolism, reactive oxygen species generation, lipid peroxidation and apoptosis in Walker 256 rat carcinosarcoma cells. Biochim Biophys Acta - Mol Cell Biol Lipids. 2001; 1533(3):207-19.
[ Links ]
131. Schley PDD, Brindley DNN, Field CJJ. (n-3) PUFA alter raft lipid composition and decrease epidermal growth factor receptor levels in lipid rafts of human breast cancer cells. J Nutr. 2007; 137(3):548-53.
[ Links ]
132. Reddy BS, Simi B, Patel N, Aliaga C, Rao CV. Effect of amount and types of dietary fat on intestinal bacterial 7-a-dehydroxylase and phosphatidylinositol-specific phospholipase C and colonic mucosal diacylglycerol kinase and PKC activities during different stages of colon tumor promotion. Cancer Res. 1996; 56:2314-20.
[ Links ]
133. McCarty MF. Fish oil may impede tumour angiogenesis and invasiveness by down-regulating protein kinase C and modulating eicosanoid production. Med Hypotheses. 1996; 46(2):107-15.
[ Links ]
134. Reddy S, Hamid R, Singh J. Dietary fat and colon cancer: modulating effect of types and amount of dietary fat on ras-p21 function during promotion and progression stages of colon cancer. Anim Res. 1997; 57:253-8.
[ Links ]
135. Novak TE, Babcock TA, Jho DH, Helton WS, Espat NJ. NF-?B inhibition by ?-3 fatty acids modulates LPS-stimulated macrophage TNF-a transcription. Am J Physiol - Lung Cell Mol Physiol. 2003; 284(1):L84-9.
[ Links ]
136. Narayanan NK, Narayanan BA, Bosland M, Condon MS, Nargi D. Docosahexaenoic acid in combination with celecoxib modulates HSP70 and p53 proteins in prostate cancer cells. Int J Cancer. 2006; 119(7):1586-98.
[ Links ]
137. Narayanan NK, Narayanan BA, Reddy BS. A combination of docosahexaenoic acid and celecoxib prevents prostate cancer cell growth in vitro and is associated with modulation of nuclear factor-kappa B, and steroid hormone receptors. Int J Oncol. 2005 Mar; 26(3):785-92.
[ Links ]
138. Lala PK, Chakraborty C. Role of nitric oxide in carcinogenesis and tumour progression. Lancet Oncol. 2001; 2(3):149-56.
[ Links ]
139. Wink DA, Vodovotz Y, Laval J, Laval F, Dewhirst MW, Mitchell JB. The multifaceted roles of nitric oxide in cancer. Carcinogenesis. 1998 May;19(5):711-21.
[ Links ]
140. Ohata T, Fakuda K, Takahashi M, Sugimura T, Wakabayashi K. Suppression of nitric oxideproduction in lipopolysaccharide-stimulated macrophage cells by omega 3 polyunsaturated fatty acids. Japanese J Cancer Res. 1997; 88(3):234-7.
[ Links ]
141. Larsson SC, Kumlin M, Ingelman-Sundberg M, Wolk A. Dietary long-chain n-3 fatty acids for the prevention of cancer: a review of potential mechanisms. Am J Clin Nutr. 2004 Jun; 79(6):935-45.
[ Links ]
142. Kim HY, Akbar M, Lau A, Edsall L. Inhibition of neuronal apoptosis by docosahexaenoic acid (22:6n-3). Role of phosphatidylserine in antiapoptotic effect. J Biol Chem. 2000 Nov 10; 275(45):35215-23
[ Links ]
143. Fu S, Wu Y, Zhang Y, Qiao M, Chen Y. Anti-cancer effects of COX-2 inhibitors and their correlation with angiogenesis and invasion in gastric cancer. 2004; 10(13): 1971-4.
[ Links ]
144. Rao CV, Hirose Y, Indranie C, Reddy BS. Modulation of experimental colon tumorigenesis by types and amounts of dietary fatty acids. Cancer Res. 2001; 61(5): 1927-33.
[ Links ]
145. Murray NR, Weems C, Chen L, Leon J, Wangsheng W, Davidson LA, et al. Protein kinase C ßII and TGFßRII in ?-3 fatty acid-mediated inhibition of colon carcinogenesis. J Cell Biol. 2002; 157(6):915-20.
[ Links ]
146. Davidson LA, Brown RE, Chang WC, Morris JS, Wang N, Carroll RJ, et al. Morphodensitometric analysis of protein kinase C beta(II) expression in rat colon: modulation by diet and relation to in situ cell proliferation and apoptosis. Carcinogenesis. 2000 Aug; 21(8):1513-9.
[ Links ]
147. Jiang YH, Lupton JR, Chapkin RS. Dietary fish oil blocks carcinogen-induced down-regulation of colonic protein kinase C isozymes. Carcinogenesis. 1997; 18(2): 351-7.
[ Links ]
148. Gökmen-Polar Y, Murray NR, Velasco MA, Gatalica Z, Fields AP. Elevated protein kinase C ßII is an early promotive event in colon carcinogenesis. Cancer Res. 2001; 61(61):1375-81.
[ Links ]
149. Murray NR, Davidson LA, Chapkin RS, Gustafson WC, Schattenberg DG, Fields AP. Overexpression of protein kinase C ßII induces colonic hyperproliferation and increased sensitivity to colon carcinogenesis. J Cell Biol. 1999; 145(4):699-711.
[ Links ]
150. Sauma S, Yan Z, Ohno S, Friedman E. Protein kinase Cß1 and protein kinase Cß2 activate mitogen-activated protein kinase and block differentiation in colon carcinoma cells. J Cell Growth Differ. 1996; 7(May):587-94.
[ Links ]
151. Saxon ML, Zhao X, Black JD. Activation of protein kinase C isozymes is associated with post- mitotic events in intestinal epithelial cells in situ. J Cell Biol. 1994; 126(3): 747-63.
[ Links ]
152. Tsai WS, Nagawa H, Kaizaki S, Tsuruo T, Muto T. Inhibitory effects of n-3 polyunsaturated fatty acids on sigmoid colon cancer transformants. J Gastroenterol. 1998 Apr; 33(2):206-12.
[ Links ]
153. Chen ZY, Istfan NW. Docosahexaenoic acid is a potent inducer of apoptosis in HT-29 colon cancer cells. Prostaglandins Leukot Essent Fatty Acids. 2000; 63:301-8.
[ Links ]
154. Clarke RG, Lund EK, Latham P, Pinder AC, Johnson IT. Effect of eicosapentaenoic acid on the proliferation and incidence of apoptosis in the colorectal cell line HT29. Lipids. 1999; 34(12):1287-95.
[ Links ]
155. Palozza P, Calviello G, Maggiano N, Lanza P, Ranelletti FO, Bartoli GM. Beta-carotene antagonizes the effects of eicosapentaenoic acid on cell growth and lipid peroxidation in WiDr adenocarcinoma cells. Free Radic Biol Med. 2000 Jan 15; 28(2):228-34.
[ Links ]
156. Jourdan M-L, Mahéo K, Barascu A, Goupille C, De Latour M, Bougnoux P, et al. Increased BRCA1 protein in mammary tumours of rats fed marine ?-3 fatty acids. Oncol Rep. 2007 Apr; 17(4):713-9.
[ Links ]
157. Gleissman H, Yang R, Martinod K, Lindskog M, Serhan CN, Johnsen JI, et al. Docosahexaenoic acid metabolome in neural tumors: identification of cytotoxic intermediates. FASEB J. 2010 Mar 1; 24(3):906-15.
[ Links ]
158. Lindskog M, Gleissman H, Ponthan F, Castro J, Kogner P, Johnsen JI. Neuroblastoma cell death in response to docosahexaenoic acid: sensitization to chemotherapy and arsenic-induced oxidative stress. Int J cancer. 2006 May 15; 118(10):2584-93.
[ Links ]
159. Serini S, Piccioni E, Merendino N, Calviello G. Dietary polyunsaturated fatty acids as inducers of apoptosis: implications for cancer. Apoptosis. 2009 Feb;14(2):135-52.
[ Links ]
160. Arita K, Kobuchi H, Utsumi T. Mechanism of apoptosis in HL-60 cells induced by n-3 and n-6 polyunsaturated fatty acids. Biochem Pharmacol. 2001 Oct 1; 62 (7): 821-8.
[ Links ]
161. Fan Z, Beresford PJ, Oh DY, Zhang D, Lieberman J. Tumor suppressor NM23-H1 is a granzyme A-activated DNase during CTL-mediated apoptosis, and the nucleosome assembly protein SET is its inhibitor. Cell. 2003 Mar 7; 112(5):659-72.
[ Links ]
162. Chambrier C, Bastard J-P, Rieusset J, Chevillotte E, Bonnefont-Rousselot D, Therond P, et al. Eicosapentaenoic acid induces mRNA expression of peroxisome proliferator-activated receptor gamma. Obes Res. 2002; 10(6):518-25.
[ Links ]
163. Fu SL, Wu YL, Zhang YP, Qiao MM, Chen Y. Anti-cancer effects of COX-2 inhibitors and their correlation with angiogenesis and invasion in gastric cancer. World J Gastroenterol. 2004; 10(13):1971-4.
[ Links ]
164. Zand H, Rhimipour A, Bakhshayesh M, Shafiee M, Nour Mohammadi I, Salimi S. Involvement of PPAR-? and p53 in DHA-induced apoptosis in Reh cells. Mol Cell Biochem. 2007 Sep 18; 304(1-2):71-7.
[ Links ]
165. Calviello G, Di Nicuolo F, Serini S, Piccioni E, Boninsegna A, Maggiano N, et al. Docosahexaenoic acid enhances the susceptibility of human colorectal cancer cells to 5-fluorouracil. Cancer Chemother Pharmacol. 2005 Jan; 55(1):12-20
[ Links ]
166. Tsujita-Kyutoku M, Yuri T, Danbara N, Senzaki H, Kiyozuka Y, Uehara N, et al. Conjugated docosahexaenoic acid suppresses KPL-1 human breast cancer cell growth in vitro and in vivo: potential mechanisms of action. Breast Cancer Res. 2004; 6(4):291-9.
[ Links ]
167. Chiu LC, Wan JM. Induction of apoptosis in HL-60 cells by eicosapentaenoic acid (EPA) is associated with downregulation of bcl-2 expression. Cancer Lett. 1999 Oct 18; 145(1-2):17-27.
[ Links ]
168. Narayanan B, Narayanan N, Reddy B. Docosahexaenoic acid regulated genes and transcription factors inducing apoptosis in human colon cancer cells. Int J Oncol. 2001 Dec; 19 (6):1255-62.
[ Links ]
169. Yamagami T, Porada CD, Pardini RS, Zanjani ED, Almeida-Porada G. Docosahexaenoic acid induces dose dependent cell death in an early undifferentiated subtype of acute myeloid leukemia cell line. Cancer Biol Ther. 2009 Feb;8(4):331-7.
[ Links ]
170. Schwartz S, Hernández A, Mark Evers B. The role of NF-kappaB/IkappaB proteins in cancer: implications for novel treatment strategies. Surg Oncol. 1999;8(3):143-53.
[ Links ]
171. Tsujii M, DuBois RN. Alterations in cellular adhesion and apoptosis in epithelial cells overexpressing prostaglandin endoperoxide synthase 2. Cell. 1995 Nov 3; 83(3):493-501.
[ Links ]
172. Sun X-M, Bratton SB, Butterworth M, MacFarlane M, Cohen GM. Bcl-2 and Bcl-xL inhibit CD95-mediated apoptosis by preventing mitochondrial release of Smac/DIABLO and subsequent inactivation of X-linked inhibitor-of-apoptosis protein. J Biol Chem. 2002 Mar 29; 277(13):11345-51
[ Links ]
173. Datta SR, Dudek H, Tao X, Masters S, Fu H, Gotoh Y, et al. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell. 1997 Oct 17; 91(2):231-41.
[ Links ]
174. Berquin I, Min Y, Wu R, Wu J. Modulation of prostate cancer genetic risk by omega-3 and omega-6 fatty acids. J Clin Invest. 2007; 117 (7):1866-1875.
[ Links ]
175. Zha J. Posttranslational N-myristoylation of BID as a molecular switch for targeting mitochondria and apoptosis. Science. 2000 Dec 1; 290 (5497):1761-5.
[ Links ]
176. Yang E, Zha J, Jockel J, Boise LH, Thompson CB, Korsmeyer SJ. Bad, a heterodimeric partner for Bcl-xL and Bcl-2, displaces bax and promotes cell death. Cell. 1995 Jan 27; 80(2):285-91.
[ Links ]
177. del Peso L, González-García M, Page C, Herrera R, Nuñez G. Interleukin-3-induced phosphorylation of BAD through the protein kinase Akt. Science. 1997 Oct 24; 278(5338):687-9.
[ Links ]
178. Blobe, G. C.; L. M. Obeid and YAH. Regulation of protein kinase C and role in cancer biology. Cancer Metastasis Rev. 1994; 13(3-4):411-431.
[ Links ]
179. Collett ED, Davidson LA, Fan YY, Lupton JR, Chapkin RS. n-6 and n-3 polyunsaturated. Am J Physiol Cell Physiol. 2001 May; 280(5):C1066-75
[ Links ]
180. Liu G, Bibus DM, Bode AM, Ma WY, Holman RT, Dong Z. Omega 3 but not omega 6 fatty acids inhibit AP-1 activity and cell transformation in JB6 cells. Proc Natl Acad Sci U S A. 2001; 98(13):7510-5.
[ Links ]
181. Rose DP, Connolly JM. Effects of fatty acids and inhibitors of eicosanoid synthesis on the growth of a human breast cancer cell line in culture effects of fatty acids and inhibitors of eicosanoid synthesis on the growth of a human breast cancer cell line in culture. Cancer Res. 1990; 50:7139-44.
[ Links ]
182. Cannizzo F, Broitman SA. Postpromotional effects of dietary marine or safflower oils on large bowel or pulmonary implants of CT-26 in mice. Cancer Res. 1989; 49 (15):4289-94.
[ Links ]
183. Paulsen JE, Elvsaas IK, Steffensen IL, Alexander J. A fish oil derived concentrate enriched in eicosapentaenoic and docosahexaenoic acid as ethyl ester suppresses the formation and growth of intestinal polyps in the Min mouse. Carcinogenesis. 1997; 18 (10):1905-10.
[ Links ]
184. Boudreau MD, Sohn KH, Rhee SH, Lee SW, Hunt JD, Hwang DH. Suppression of tumor cell growth both in nude mice and in culture by n-3 polyunsaturated fatty acids: mediation through cyclooxygenase-independent pathways. Cancer Res. 2001 Feb 15; 61(4):1386-91.
[ Links ]
185. Calviello G, Di Nicuolo F, Gragnoli S, Piccioni E, Serini S, Maggiano N, et al. n-3 PUFAs reduce VEGF expression in human colon cancer cells modulating the COX-2/PGE2 induced ERK-1 and -2 and HIF-1alpha induction pathway. Carcinogenesis. 2004 Dec; 25(12):2303-10.
[ Links ]
186. Minoura T, Takata T, Sakaguchi M, Takada H, Yamamura M, Hioki K, et al. Effect of dietary eicosapentaenoic acid on azoxymethane-induced colon carcinogenesis in rats. Cancer Res. 1988; 48(17):4790-4.
[ Links ]
187. Hendrickse CW, Keighley MRB, Neoptolemos JP. Dietary ?-3 fats reduce proliferation and tumor yields at colorectal anastomosis in rats. Gastroenterology. 1995; 109 (2):431-9.
[ Links ]
188. Davidson LA. Carcinogen and dietary lipid regulate ras expression and localization in rat colon without affecting farnesylation kinetics. Carcinogenesis. 1999; 20(5): 785-91.
[ Links ]
189. Dommels EM, Alink GM, Peter J. Dietary n-6 and n-3 polyunsaturated fatty acids and colorectal carcinogenesis- results from cultured colon cells, animal models and human studies. Environ Toxicol Pharmacol. 2002 Jul; 11 (3-4):297-308
[ Links ]
190. Reddy BS, Burill C, Rigotty J. Effect of diets high in ?-3 and ?-6 fatty acids on initiation and postinitiation stages of colon carcinogenesis. Cancer Res. 1991; 51:487-91.
[ Links ]
191. Takahashi M, Fukutake M, Isoi T, Fukuda K, Sato H, Yazawa K, et al. Suppression of azoxymethane-induced rat colon carcinoma development by a fish oil component, docosahexaenoic acid (DHA). Carcinogenesis. 1997; 18(7):1337-42.
[ Links ]
192. Arterburn L, Hall E, Oken H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am J Cinical Nutr. 2006; 83(6S):1467S-1476S.
[ Links ]
193. Hunter DJ, Rimm EB, Sacks FM, Stampfer MJ, Colditz GA, Litin LB, et al. Comparison of measures of fatty acid intake by subcutaneous fat aspirate, food frequency questionnaire, and diet records in a free-living population of US men. Am J Epidemiol. 1992 Feb 15; 135(4):418-27.
[ Links ]
194. Pardini RS. Nutritional intervention with omega-3 fatty acids enhances tumor response to anti-neoplastic agents. Chem Biol Interact. 2006;162(2):89-105.
[ Links ]
195. Hardman WE, Avula CPR, Fernandes G, Cameron IL. Three percent dietary fish oil concentrate increased efficacy of doxorubicin against MDA-MB 231 breast cancer xenografts 1. 2001; 7 (July):2041-9.
[ Links ]
196. Boutard V, Fouqueray B, Philippe C, Perez J, Baud L. Fish oil supplementation and essential fatty acid deficiency reduce nitric oxide synthesis by rat macrophages. Kidney Int. 1994; 46(5):1280-6.
[ Links ]
197. Shao Y, Pardini L, Pardini RS. Dietary menhaden oil enhances mitomycin C antitumor activity toward human mammary carcinoma MX-1. Lipids. 1995; 30(11): 1035-45.
[ Links ]
198. Chen J, Hui E, Ip T, Thompson LU. Dietary flaxseed enhances the inhibitory effect of tamoxifen on the growth of estrogen-dependent human breast cancer (MCF-7) in nude mice. Clin Cancer Res. 2004; 10(22):7703-11.
[ Links ]
199. Bougnoux P, Hajjaji N, Ferrasson MN, Giraudeau B, Couet C, Le Floch O. Improving outcome of chemotherapy of metastatic breast cancer by docosahexaenoic acid: a phase II trial. Br J Cancer. 2009 Dec 15; 101(12):1978-85.
[ Links ]
200. Rusca A, Di Stefano AFD, Doig MV, Scarsi C, Perucca E. Relative bioavailability and pharmacokinetics of two oral formulations of docosahexaenoic acid/ eicosapentaenoic acid after multiple-dose administration in healthy volunteers. Eur J Clin Pharmacol. 2009; 65(5):503-10.
[ Links ]
201. Childs CE, Romeu-Nadal M, Burdge GC, Calder PC. Gender differences in the n-3 fatty acid content of tissues. Proc Nutr Soc. 2008; 67(01):19-27.
[ Links ]
202. Murphy RA, Mourtzakis M, Mazurak VC. n-3 polyunsaturated fatty acids: the potential role for supplementation in cancer. Curr Opin Clin Nutr Metab Care. 2012; 15(3):246-51.
[ Links ]
203. Tsuzuki T, Shibata A, Kawakami Y, Nakagawa K, Miyazawa T. Conjugated eicosapentaenoic acid inhibits vascular endothelial growth factor-induced angiogenesis by suppressing the migration of human umbilical vein endothelial cells 1. J Nutr Dis. 2007; 137(3):641-6.
[ Links ]
204. Tsuji M, Murota SI, Morita I. Docosapentaenoic acid (22:5, n-3) suppressed tube-forming activity in endothelial cells induced by vascular endothelial growth factor. Prostaglandins Leukot Essent Fat Acids. 2003; 68(5):337-42.
[ Links ]
205. Yang SP, Morita I, Murota SI. Eicosapentaenoic acid attenuates vascular endothelial growth factor-induced proliferation via inhibiting Flk-1 receptor expression in bovine carotid artery endothelial cells. J Cell Physiol. 1998; 176(2):342-9.
[ Links ]
206. Fox PL, DiCorleto PE. Fish oils inhibit endothelial cell production of platelet-derived growth factor-like protein. Science. 1988 Jul 22; 241(4864):453-6.
[ Links ]
207. Terano T, Shiina T, Tamura Y. Terano T, Shiina T, Tamura Y. Eicosapentaenoic acid suppressed the proliferation of vascular smooth muscle cells through modulation of various steps of growth signals. Lipids. 1996; 31 (Suppl):S301-4.
[ Links ]
208. Reddy BS, Patlolla JM, Simi B, Wang SH, Rao CV. Prevention of colon cancer by low doses of celecoxib, a cyclooxygenase inhibitor, administered in diet rich in ?-3 polyunsaturated fatty acids. Cancer Res. 2005; 65(17):8022-7.
[ Links ]
209. Dimmeler S, Hermann C, Galle J, Zeiher AM. Upregulation of superoxide dismutase and nitric oxide synthase mediates the apoptosis-suppressive effects of shear stress on endothelial cells. Arter Thromb Vasc Biol. 1999; 19(3):656-64.
[ Links ]
210. Cianchi F, Cortesini C, Bechi P, Fantappiè O, Messerini L, Vannacci A, et al. Up-regulation of cyclooxygenase 2 gene expression correlates with tumor angiogenesis in human colorectal cancer. Gastroenterology. 2001; 121(6):1339-47.
[ Links ]
211. Ziche M, Morbidelli L, Choudhuri R, Zhang HT, Donnini S, Granger HJ, et al. Nitric oxide synthase lies downstream from vascular endothelial growth factor-induced but not basic fibroblast growth factor-induced angiogenesis. J Clin Invest. 1997; 99(11): 2625-34.
[ Links ]
212. Komatsu W, Ishihara K, Murata M, Saito H, Shinohara K. Docosahexaenoic acid suppresses nitric oxide production and inducible nitric oxide synthase expression in interferon-gamma plus lipopolysaccharide-stimulated murine macrophages by inhibiting the oxidative stress. Free Radic Biol Med. 2003; 34(8):1006-16.
[ Links ]
213. Narayanan BA, Narayanan NK, Simi B, Reddy BS. Modulation of inducible nitric oxide synthase and related proinflammatory genes by the omega-3 fatty acid docosahexaenoic acid in human colon cancer cells. Cancer Res. 2003; 63(5):972-9.
[ Links ]
214. Narayanan BA, Narayanan NK, Desai D, Pittman B, Reddy BS. Effects of a combination of docosahexaenoic acid and 1, 4-phenylene bis(methylene) selenocyanate on cyclooxygenase 2, inducible nitric oxide synthase and ß-catenin pathways in colon cancer cells. Carcinogenesis. 2004; 25(12):2443-9.
[ Links ]
215. Gleissman H, Segerström L, Hamberg M, Ponthan F, Lindskog M, Johnsen JI, et al. Omega-3 fatty acid supplementation delays the progression of neuroblastoma in vivo. Int J Cancer. 2011; 128(7):1703-11.
[ Links ]
216. Swamy MV, Citineni B, Patlolla JMR, Mohammed A, Zhang Y, Rao CV. Prevention and treatment of pancreatic cancer by curcumin in combination with omega-3 fatty acids. Nutr Cancer. 2008; 60 (suppl. 1):81-9.
[ Links ]
217. Tevar R, Jho DH, Babcock T, Helton WS, Espat NJ. Omega-3 fatty acid supplementation reduces tumor growth and vascular endothelial growth factor expression in a model of progressive non-metastasizing malignancy. JPEN J Parenter Enteral Nutr. 2002; 26(5):285-9.
[ Links ]
218. Rose DP, Connolly JM. Omega-3 fatty acids as cancer chemopreventive agents. Pharmacol Ther. 1999; 83(3):217-44.
[ Links ]
219. Mukutmoni-Norris M, Hubbard NE, Erickson KL. Modulation of murine mammary tumor vasculature by dietary n-3 fatty acids in fish oil. Cancer Lett. 2000; 150 (1): 101-9.
[ Links ]
220. Bommareddy A, Arasada BL, Mathees DP, Dwivedi C. Chemopreventive effects of dietary flaxseed on colon tumor development. Nutr Cancer. 2006 Jul 1; 54(2): 216-22.
[ Links ]
221. Schaefer MB, Wenzel A, Fischer T, Braun-Dullaeus RC, Renner F, Dietrich H, et al. Fatty acids differentially influence phosphatidylinositol 3-kinase signal transduction in endothelial cells: impact on adhesion and apoptosis. Atherosclerosis. 2008; 197(2): 630-7.
[ Links ]
222. Ambs S, Hussain SP, Harris CC. Interactive effects of nitric oxide and the p53 tumor suppressor gene in carcinogenesis and tumor progression. FASEB J. 1997 May 1; 11(6): 443-8.
[ Links ]
223. Yi L, Zhang Q-Y, Mi M-T. Role of Rho GTPase in inhibiting metastatic ability of human prostate cancer cell line PC-3 by omega-3 polyunsaturated fatty acid. Ai Zheng. 2007 Dec; 26 (12):1281-6.
[ Links ]
224. Cooke JP, Losordo DW. Nitric oxide and angiogenesis. 2002; 2133-5.
[ Links ]
225. Ohno R, Yoshinaga K, Fujita T, Hasegawa K, Iseki H, Tsunozaki H, et al. Depth of invasion parallels increased cyclooxygenase-2 levels in patients with gastric carcinoma. Cancer. 2001; 91(10):1876-81.
[ Links ]
226. Iwamoto S, Senzaki H, Kiyozuka Y, Ogura E, Takada H, Hioki K, et al. Effects of fatty acids on liver metastasis of ACL-15 rat colon cancer cells. Nutr Cancer. 1998 Jan 1; 31(2):143-50.
[ Links ]
227. Kontogiannea M, Sc B, Gupta A, Sc B, Ntanios F, Ph D, et al. n-3 fatty acids decrease endothelial adhesion of human colorectal carcinoma cells 1. 2000; 205: 201-5.
[ Links ]
228. Rose DMP, Connolly CJM. Effect of omega-3 fatty acids on the progression of metastases after the surgical excision of human breast cancer cell solid tumors growing in nude mice. Clin Cancer Res. 1996; 2(914):1751-6.
[ Links ]
229. Press CRUK. Breast screening saves lives say experts. Cancer Research UK. 2009 [cited 2018 Jan 31]. p.1. https://www.cancerresearchuk.org/
[ Links ]
230. Sims AH, Howell A, Howell SJ, Clarke RB. Origins of breast cancer subtypes and therapeutic implications. Nat Clin Pract Oncol. 2007 Sep 1; 4:516.
[ Links ]
231. Matsuda I, Matsubara N, Aoyama N, Hamanaka M, Yamagishi D, Kuno T, et al. Metastatic lobular carcinoma of the breast masquerading as a primary rectal cancer. World J Surg Oncol. 2012; 10: 231.
[ Links ]
232. Chang H-Y, Huang H-C, Huang T-C, Yang P-C, Wang Y-C, Juan H-F. Ectopic ATP synthase blockade suppresses lung adenocarcinoma growth by activating the unfolded protein response. Cancer Res. 2012 Sep 15; 72 (18):4696-706.
[ Links ]
233. Levine MN, Gent M, Hirsh J, Arnold A, Goodyear MD, Hryniuk W, et al. The thrombogenic effect of anticancer drug therapy in women with stage II breast cancer. N Engl J Med. 1988 Feb 18; 318(7):404-7.
[ Links ]
234. Benson JR, Querci della Rovere G. Sentinel lymph node in breast cancer. Lancet (London, England). 1999. Dec 4; 354 (9194):1998; author reply 1999.
[ Links ]
235. Narod SA. Hormone replacement therapy and the risk of breast cancer. Nat Rev Clin Oncol. 2011; 8(11):669-76.
[ Links ]
236. Beatson GT. On the treatment of inoperable cases of carcinoma of the mamma: suggestions for a new method of treatment, with illustrative cases. The Lancet. 1896; July 11 148 (3802), p104-107
[ Links ]
237. Dowsett M. Proliferation and apoptosis as markers of benefit in neoadjuvant endocrine therapy of breast cancer. Clin Cancer Res. 2006; 12(3s):1024s-1030s.
[ Links ]
238. Doisneau-Sixou SF, Sergio CM, Carroll JS, Hui R, Musgrove EA, Sutherland RL. Estrogen and antiestrogen regulation of cell cycle progression in breast cancer cells. Endocr Relat Cancer. 2003; 10(2):179-86.
[ Links ]
239. Wakeling AE. Similarities and distinctions in the mode of action of different classes of antioestrogens. Endocr Relat Cancer. 2000; 7(1):17-28.
[ Links ]
240. Klinge CM, Jernigan SC, Smith SL, Tyulmenkov VV, Kulakosky PC. Estrogen response element sequence impacts the conformation and transcriptional activity of estrogen receptor a. Mol Cell Endocrinol. 2001; 174(1-2):151-66.
[ Links ]
241. Zaichuk T, Ivancic D, Scholtens D, Schiller C, Khan SA. Tissue-specific transcripts of human steroid sulfatase are under control of estrogen signaling pathways in breast carcinoma. J Steroid Biochem Mol Biol. 2007; 105(1-5):76-84.
[ Links ]
242. Stanway SJ, Delavault P, Purohit A, Woo LWL, Thurieau C, Potter BVL, et al. Steroid sulfatase: a new target for the endocrine therapy of breast cancer. Oncologist. 2007; 12 (4):370-4.
[ Links ]
243. Marjoribanks J, Farquhar C, Roberts H, Lethaby A. Long term hormone therapy for perimenopausal and postmenopausal women. Cochrane Database Syst Rev. 2012 Jul 11; (7):CD004143
[ Links ]
244. Bray F, Colombet M, Mery L, Piñeros M, Znaor A, Zanetti R and Ferlay J, editors (2017). Cancer incidence in five continents, Vol. XI (electronic version). Lyon: International Agency for Research on Cancer. Available from: http://ci5.iarc.fr, accessed [date].
[ Links ]
245. Miller WR, Anderson TJ, Jack WJL. Relationship between tumour aromatase activity, tumour characteristics and response to therapy. J Steroid Biochem Mol Biol. 1990; 37(6):1055-9.
[ Links ]
246. van Landeghem AA, Poortman J, Nabuurs M, Thijssen JHH. Endogenous concentration and subcellular distribution of estrogens in normal and malignant human breast tissue. Cancer Res. 1985; 45(June):2900-6.
[ Links ]
247. Vermeulen A, Deslypere JP, Paridaens R, Lecrerq G, Roy F, Heuson JC. Aromatase, 17ß-hydroxysteroid dehydrogenase and intratissular sex hormone concentrations in cancerous and normal glandular breast tissue in postmenopausal women. Eur J Cancer Clin Oncol. 1985; 22(4):515-25.
[ Links ]
248. Geisler J, Detre S, Berntsen H, Ottestad L, Lindtjrn B, Dowsett M, et al. Influence of neoadjuvant anastrozole (Arimidex) on intratumoral estrogen levels and proliferation markers in patients with locally advanced breast cancer. Clin Cancer Res. 2001; 7(5):1230-6.
[ Links ]
249. Ma CX, Adjei AA, Salavaggione OE, Coronel J, Pelleymounter L, Wang L, et al. Human aromatase: gene resequencing and functional genomics. Cancer Res. 2005; 65(23):11071-82.
[ Links ]
250. Fournier A, Berrino F, Riboli E, Avenel V, Clavel-Chapelon F. Breast cancer risk in relation to different types of hormone replacement therapy in the E3N-EPIC cohort. Int J Cancer. 2005; 114(3):448-54.
[ Links ]
251. Schairer C, Lubin J, Troisi R, Sturgeon S, Brinton L, Hoover R, et al. Menopausal Estrogen and Estrogen-Progestin Replacement Therapy and Breast Cancer Risk. JAMA. 2000; 283(4):485.
[ Links ]
252. Key T, Appleby P, Barnes I, Reeves G. Endogenous sex hormones and breast cancer in postmenopausal women: reanalysis of of nine prospective studies. J Natl Cancer Inst. 2002; 94(8):606-16.
[ Links ]
253. Lee SA, Ross RK, Pike MC. An overview of menopausal oestrogen-progestin hormone therapy and breast cancer risk. Br J Cancer. 2005; 92(11):2049-58.
[ Links ]
254. Howell A, Robertson JFR, Abram P, Lichinitser MR, Elledge R, Bajetta E, et al. Comparison of fulvestrant versus tamoxifen for the treatment of advanced breast cancer in postmenopausal women previously untreated with endocrine therapy: a multinational, double-blind, randomized trial. J Clin Oncol. 2004; 22(9):1605-13.
[ Links ]
255. Buzdar AU. Fulvestrant-A novel estrogen receptor antagonist for the treatment of advanced breast cancer. Drugs Today (Barc). 2008 Sep; 44(9):679-92.
[ Links ]
256. Long X, Nephew KP. Fulvestrant (ICI 182,780)-dependent interacting proteins mediate immobilization and degradation of estrogen receptor-a. J Biol Chem. 2006; 281(14):9607-15.
[ Links ]
257. Lobysheva NV, Selin AA, Vangeli IM, Byvshev IM, Yaguzhinsky LS. Glutamate induces H2O2 synthesis in nonsynaptic brain mitochondria. Free Radic Biol Med. 2013 Dec; 65:428-435.
[ Links ]
258. Yue W, Santen RJ, Wang JP, Li Y, Verderame MF, Bocchinfuso WP, et al. Genotoxic metabolites of estradiol in breast: potential mechanism of estradiol induced carcinogenesis. J Steroid Biochem Mol Biol. 2003 Sep; 86 (3-5):477-86.
[ Links ]
259. Devanesan P. Catechol estrogen metabolites and conjugates in mammary tumors and hyperplastic tissue from estrogen receptor-alpha knock-out (ERKO)/Wnt-1 mice: implications for initiation of mammary tumors. Carcinogenesis. 2001; 22(9):1573-6.
[ Links ]
260. Cavalieri EL, Stack DE, Devanesan PD, Todorovic R, Dwivedy I, Higginbotham S, et al. Molecular origin of cancer: catechol estrogen-3,4-quinones as endogenous tumor initiators. Proc Natl Acad Sci USA. 1997; 94(20):10937-42.
[ Links ]
261. Santen RJ, Harvey HA. Use of aromatase inhibitors in breast carcinoma. Endocr Relat Cancer. 1999; 6(1):75-92.
[ Links ]
262. Webb P, Nguyen P, Valentine C, Lopez GN, Kwok GR, McInerney E, et al. The estrogen receptor enhances AP-1 activity by two distinct mechanisms with different requirements for receptor transactivation functions. Mol Endocrinol. 1999; 13(10):1672-85.
[ Links ]
263. Razandi M, Pedram A, Greene GL, Levin ER. Cell membrane and nuclear estrogen receptors (ERs) originate from a single transcript: studies of ERa and ERß expressed in Chinese hamster ovary cells. Mol Endocrinol. 1999; 13(2):307-19.
[ Links ]
264. Sabbah M, Courilleau D, Mester J, Redeuilh G. Estrogen induction of the cyclin D1 promoter: involvement of a cAMP response-like element. Proc Natl Acad Sci. 1999; 96 (20):11217-22.
[ Links ]
265. Levin ER. Bidirectional signaling between the estrogen receptor and the epidermal growth factor receptor. Mol Endocrinol. 2003; 17(3):309-17.
[ Links ]
266. Stoica GE, Franke TF, Moroni M, Mueller S, Morgan E, Iann MC, et al. Effect of estradiol on estrogen receptor-a gene expression and activity can be modulated by the ErbB2/PI 3-K/Akt pathway. Oncogene. 2003; 22(39):7998-8011.
[ Links ]
267. Sekeris CE. The mitochondrial genome: a possible primary site of action of steroid hormones. In Vivo. 1990; 4 (5):317-320.
[ Links ]
268. Liao JK. Cross-coupling between the oestrogen receptor and phosphoinositide 3-kinase. Biochem Soc Trans. 2003; 31:66-70.
[ Links ]
269. Aronica SM, Kraus WL, Katzenellenbogen BS. Estrogen action via the cAMP signaling pathway: stimulation of adenylate cyclase and cAMP-regulated gene transcription. Proc Natl Acad Sci USA. 1994; 91(18):8517-21.
[ Links ]
270. Migliaccio A, Di Domenico M, Castoria G, De Falco A, Bontempo P, Nola E, et al. Tyrosine kinase/p21ras/MAP-kinase pathway activation by estradiol-receptor complex in MCF-7 cells. EMBO J. 1996; 15(6):1292-300.
[ Links ]
271. Song RX-D, McPherson R a, Adam L, Bao Y, Shupnik M, Kumar R, et al. Linkage of rapid estrogen action to MAPK activation by ERa-Shc association and Shc pathway activation. Mol Endocrinol. 2002; 16(1):116-27.
[ Links ]
272. Brodie AMH, Schwarzel WC, Shaikh AA, Brodie HJ. The effect of an aromatase inhibitor, 4-hydroxy-4-androstene- 3, 17-dione, on estrogen-dependent processes in reproduction and breast cancer. Endocrinology. 1977 Jun 1; 100(6): 1684-95.
[ Links ]
273. Longcope C, Pratt JH, Schneider SH, Fineberg SE. Aromatization of androgens by muscle and adipose tissue in vivo. J Clin Endocrinol Metab. 1978; 46(1):146-52.
[ Links ]
274. Miller WR. Aromatase activity in breast tissue. J Steroid Biochem Mol Biol. 1991; 39(5):783-90.
[ Links ]
275. Ryan KJ. Biological aromatization of steroids. J Biol Chem. 1958; 234(8):268-73.
[ Links ]
276. Meigs RA, Ryan KJ. Cytochrome P-450 and steroid biosynthesis in the human placenta. Biochim Biophys Acta - Mol Cell Biol Lipids. 1968; 165 (1):476-82.
[ Links ]
277. Fishman J, Rajus MS. Mechanism of estrogen biosynthesis. J Biol Chem. 1981; 256 (9): 4472-7.
[ Links ]
278. Lonning P. Aromatase inhibitors in breast cancer. Endocr Relat Cancer. 2004; 11(2): 179-89.
[ Links ]
279. Schweikert HU, Milewich L, Wilson JD. Aromatization of androstenedione by cultured human fibroblasts. J Clin Endocrinol Metab. 1976; 43(4):785-95.
[ Links ]
280. Miller WR, Forrest APM. Oestradiol synthesis by a human breast carcinoma. Lancet. 1974; 304 (7885):866-8.
[ Links ]
281. Perel E, Wilkins D, Killinger DW. The conversion of androstenedione to estrone, estradiol, and testosterone in breast tissue. J Steroid Biochem. 1980; 13(1):89-94.
[ Links ]
282. Reed MJ, Owen AM, Lai LC, Coldham NG, Ghilchik MW, Shaikh NA, et al. In situ oestrone synthesis in normal breast and breast tumour tissues: effect of treatment with 4-hydroxyandrostenedione. Int J Cancer. 1989; 44(2):233-7.
[ Links ]
283. Santen RJ, Santner S, Davis B, Veldhuis J, Samojlik E, Ruby E. Aminoglutethimide inhibits extraglandular estrogen production in postmenopausal women with breast carcinoma. J Clin Endocrinol Metab. 1978; 47(6):1257-65.
[ Links ]
284. Häusler A, Schenkel L, Krähenbühl C, Monnet G, Bhatnagar AS. An in vitro method to determine the selective inhibition of estrogen biosynthesis by aromatase inhibitors. J Steroid Biochem. 1989 Jul; 33(1):125-31.
[ Links ]
285. Miller WR. Biology of aromatase inhibitors: pharmacology/endocrinology within the breast. Endocr Relat Cancer. 1999; 6(2):187-95.
[ Links ]
286. Pittman JA, Brown RW. Antithyroid and antiadrenocortical activity of aminoglutethimide. J Clin Endocrinol Metab. 1966; 26(9):1014-6.
[ Links ]
287. Demers LM, Lipton A, Harvey HA, Kambic KB, Grossberg H, Brady C, et al. The efficacy of CGS 20267 in suppressing estrogen biosynthesis in patients with advanced stage breast cancer. J Steroid Biochem Mol Biol.1993 Mar; 44(4-6):687-91.
[ Links ]
288. Falkson CI, Falkson HC. A randomised study of CGS 16949A (fadrozole) versus tamoxifen previously untreated postmenopausal patients with metastatic breast cancer. Ann Oncol. 1996; 7(5):465-9.
[ Links ]
289. Thürlimann B, Beretta K, Bacchi M, Castiglione-Gertsch M, Goldhirsch A, Jungi WF, et al. First-line fadrozole HCI (CGS 16949A) versus tamoxifen in postmenopausal women with advanced breast cancer. Prospective randomised trial of the Swiss Group for Clinical Cancer Research SAKK 20/88. Ann Oncol. 1996; 7(5):471-9.
[ Links ]
290. Lang M, Batzl C, Furet P, Bowman R, Häusler A, Bhatnagar AS. Structure-activity relationships and binding model of novel aromatase inhibitors. J Steroid Biochem Mol Biol. 1993; 44(4):421-8.
[ Links ]
291. Furet P, Batzl C, Bhatnagar A, Francotte E, Rihs G, Lang M. Aromatase inhibitors: synthesis, biological activity, and binding mode of azole-type compounds. J Med Chem. 1993 May 14; 36 (10):1393-400.
[ Links ]
292. Bhatnagar AS, Brodie AMH, Long BJ, Evans DB, Miller WR. Intracellular aromatase and its relevance to the pharmacological efficacy of aromatase inhibitors. J Steroid Biochem Mol Biol. 2001;76 (1-5):199-202.
[ Links ]
293. Long BJ, Tilghman SL, Yue W, Thiantanawat A, Grigoryev DN, Brodie AMH. The steroidal antiestrogen ICI 182,780 is an inhibitor of cellular aromatase activity. J Steroid Biochem Mol Biol. 1998; 67(4):293-304.
[ Links ]
294. Darzynkiewicz Z, Smolewski P, Bedner E. Use of flow and laser scanning cytometry to study mechanisms regulating cell cycle and controlling cell study mechanisms regulating cell cycle and controlling cell death. Clin Lab Med. 2001 Dec; 21 (4):857-73.
[ Links ]
295. Odum J, Ashby J. Detection of aromatase inhibitors in vitro using rat ovary microsomes. Toxicol Lett. 2002; 129:119-22.
[ Links ]
296. Haynes BP, Dowsett M, Miller WR, Dixon JM, Bhatnagar AS. The pharmacology of letrozole. J Steroid Biochem Mol Biol. 2003 Oct;87 (1):35-45.
[ Links ]
297. Pfister CU, Martoni A, Zamagni C, Lelli G, De Braud F, Souppart C, et al. Effect of age and single versus multiple dose pharmacokinetics of letrozole (Femara) in breast cancer patients. Biopharm Drug Dispos. 2001; 22 (5):191-7.
[ Links ]
298. Demers LM. Effects of fadrozole (CGS 16949A) and letrozole (CGS 20267) on the inhibition of aromatase activity in breast cancer patients. Breast Cancer Res Treat. 1994; 30 (1):95-102.
[ Links ]
299. Dowsett M, Jones A, Johnston SRD, Jacobs S, Trunet P, Smith IE. In vivo measurement of aromatase inhibition by letrozole (CGS 20267) in postmenopausal patients with breast cancer. Clin Cancer Res. 1995; 1(12):1511-5.
[ Links ]
300. Geisler J, King N, Dowsett M, Ottestad L, Lundgren S, Walton P, et al. Influence of anastrozole (Arimidex), a selective, non-steroidal aromatase inhibitor, on in vivo aromatisation and plasma oestrogen levels in postmenopausal women with breast cancer. Br J Cancer. 1996;74: 1286-91.
[ Links ]
301. Iveson TJ, Smith IE, Ahern J, Smithers DA, Trunet PF, Dowsett M. Phase I study of the oral nonsteroidal aromatase inhibitor CGS 20267 in healthy postmenopausal women. J Clin Endocrinol Metab. 1993 Aug 1;77 (2):324-31.
[ Links ]
302. Bajetta E. Double-blind, randomised, multi-centre endocrein trial comparing two letrozole doses, in postmenopausal breast cancer patients. Eur J Cancer. 1999; 35 (2): 208-13.
[ Links ]
303. Goss, P. E. Ingle, J. N., Martino, S., Robert, N. J., Muss, H. B., Piccart, M. J, Livingston RB. A randomized trial of letrozole in postmenopausal women after five years of tamoxifen therapy for early-stage breast cancer. N Engl J Med. 2003;349 (19):1793-802.
[ Links ]
304. Goss PE, Ingle JN, Martino S, Robert NJ, Muss HB, Piccart MJ, et al. Randomized trial of letrozole following tamoxifen as extended adjuvant therapy in receptor-positive breast cancer: updated findings from NCIC CTG MA.17. J Natl Cancer Inst. 2005; 97(17):1262-71.
[ Links ]
305. Ellis BMJ, Coop A, Singh B, Mauriac L, Llombert-cussac A, Ja F, et al. Breast cancer: evidence from a phase III. 2001;19 (18):3808-16.
[ Links ]
306. Dixon JM, Renshaw L, Young O, Murray J, Macaskill EJ, Mchugh M, et al. Letrozole suppresses plasma estradiol and estrone sulphate more completely than anastrozole in postmenopausal women with breast cancer. J Clin Oncol. 2008; 26(10):0-5.
[ Links ]
307. Brodie A, Lu Q, Liu Y, Long B. Aromatase inhibitors and their antitumor effects in model systems. Endocr Relat Cancer. 1999;6: 205-10.
[ Links ]
308. Yue W, Wang J, Savinov A, Brodie A. Effect of aromatase inhibitors on growth of mammary tumors in a nude mouse model effect of aromatase inhibitors on growth of mammary tumors in a nude mouse model. 1995; 3073-7.
[ Links ]
309. Lu Q, Yue W, Wang J, Liu Y, Long B, Brodie A. The effects of aromatase inhibitors and antiestrogens in the nude mouse model. Breast Cancer Res Treat. 1998 Jul;50 (1) :63-71.
[ Links ]
310. Long BJ, Jelovac D, Handratta V, Thiantanawat A, MacPherson N, Ragaz J, et al. Therapeutic strategies using the aromatase inhibitor letrozole and tamoxifen in a breast cancer model. J Natl Cancer Inst. 2004 Mar 17; 96 (6):456-65.
[ Links ]
311. Jelovac D, Macedo L, Goloubeva OG, Handratta V, Brodie AMH. Additive antitumor effect of aromatase inhibitor letrozole and antiestrogen fulvestrant in a postmenopausal breast cancer model additive antitumor effect of aromatase inhibitor letrozole breast cancer model. Cancer Res. 2005 Jun 15; 65(12):5439-44
[ Links ]
312. Benelli R, Venè R, Minghelli S, Carlone S, Gatteschi B, Ferrari N. Celecoxib induces proliferation and amphiregulin production in colon subepithelial myofibroblasts, activating erk1-2 signaling in synergy with EGFR. Cancer Lett. 2013 Jan 1;328(1):73-82.
[ Links ]
313. Lim SJ, Lee E, Lee EH, Kim SY, Cha JH, Choi H, et al. Docosahexaenoic acid sensitizes colon cancer cells to sulindac sulfide-induced apoptosis. Oncol Rep. 2012;27(6):2023-30.
[ Links ]
314. Ding W-Q, Liu B, Vaught JL, Palmiter RD, Lind SE. Clioquinol and docosahexaenoic acid act synergistically to kill tumor cells. Mol Cancer Ther. 2006;5(7):1864-72.
[ Links ]
315. Altenburg JD, Bieberich AA, Terry C, Harvey KA, VanHorn JF, Xu Z, et al. A synergistic antiproliferation effect of curcumin and docosahexaenoic acid in SK-BR-3 breast cancer cells: unique signaling not explained by the effects of either compound alone. BMC Cancer. 2011;11 (1):149.
[ Links ]
316. Shaikh I a a, Brown I, Wahle KWJ, Heys SD. Enhancing cytotoxic therapies for breast and prostate cancers with polyunsaturated fatty acids. Nutr Cancer. 2010;62 (3):284-96.
[ Links ]
317. Steinert BW, Tang DG, Grossi IM, Umbarger LA, Honn K V. Studies on the role of platelet eicosanoid metabolism and integrin alpha IIb beta 3 in tumor-cell-induced platelet aggregation. Int J Cancer. 1993;54 (1):92-101.
[ Links ]
318. Menter DG, Onoda JM, Taylor JD, Honn K V. Effects of prostacyclin on tumor cell-induced platelet aggregation. Cancer Res. 1984;44 (1):450-6.
[ Links ]
319. Tang K, Honn KV. 2(S)-HETE in cancer metastasis. Adv Exp Med Biol. 1999;447:181.
[ Links ]
320. Jurasz P, Alonso-Escolano D, Radomski MW. Platelet-cancer interactions: mechanisms and pharmacology of tumour cell-induced platelet aggregation. Br J Pharmacol. 2004 Dec; 143 (7):819-26.
[ Links ]
321. Tang DG, Renaud C, Stojakovic S, Diglio CA, Porter A, Honn K V. 12(S)-HETE is a mitogenic factor for microvascular endothelial cells: its potential role in angiogenesis. Biochem Biophys Res Commun. 1995 Jun 15;211 (2):462-8
[ Links ]
322. Gross ER, Hsu AK, Gross GJ. Acute aspirin treatment abolishes, whereas acute ibuprofen treatment enhances morphine-induced cardioprotection: role of 12-lipoxygenase. J Pharmacol Exp Ther. 2004;310 (1):185-91.
[ Links ]
323. Apraiz A, Idkowiak-Baldys JK, Boyano MD, Pérez-Yarza G, Hannun YA, Asumendi A. Evaluation of bioactive sphingolipids in 4-HPR-resistant leukemia cells. BMC Cancer. 2011 Nov 7;11: 477
[ Links ]
324. Liu Y-Y, Yu JY, Yin D, Patwardhan GA, Gupta V, Hirabayashi Y, et al. A role for ceramide in driving cancer cell resistance to doxorubicin. FASEB J . 2008; 22 (7): 2541-51.
[ Links ]
325. Ahn Q, Jeong SJ, Lee HJ, Kwon HY, Han I, Kim HS, Lee HJ, Lee EO, Ahn KS, Jung MH, Zhu S, Chen CY, Kim SH. Inhibition of cyclooxygenase-2-dependent survivin mediates decursin-induced apoptosis in human KBM-5 myeloid leukemia cells. Cancer Lett. 2010 Dec 8;298 (2):212-21 .
[ Links ]
326. Baumgartner M, Sturlan S, Roth E, Wessner B, Bachleitner-Hofmann T. Enhancement of arsenic trioxide-mediated apoptosis using docosahexaenoic acid in arsenic trioxide-resistant solid tumor cells. Int J Cancer. 2004;112(4):707-12.
[ Links ]
327. Aroui S, Brahim S, Hamelin J, De Waard M, Bréard J, Kenani A. Conjugation of doxorubicin to cell penetrating peptides sensitizes human breast MDA-MB 231 cancer cells to endogenous TRAIL-induced apoptosis. Apoptosis. 2009;14(11): 1352-65.
[ Links ]
328. Györffy B, Serra V, Jürchott K, Abdul-Ghani R, Garber M, Stein U, et al. Prediction of doxorubicin sensitivity in breast tumors based on gene expression profiles of drug-resistant cell lines correlates with patient survival. Oncogene. 2005;24(51): 7542-51.
[ Links ]
329. Patel NM, Nozaki S, Shortle NH, Bhat-Nakshatri P, Newton TR, Rice S, et al. Paclitaxel sensitivity of breast cancer cells with constitutively active NF-?B is enhanced by IkBa super-repressor and parthenolide. Oncogene.2000 Aug 24;19 (36):4159-69.
[ Links ]
330. Horia E, Watkins BA. Complementary actions of docosahexaenoic acid and genistein on COX-2 PGE 2 and invasiveness in MDA-MB-231 breast cancer cells. Carcinogenesis. 2007 Apr;28 (4):809-15.
[ Links ]




![SEROTONERGIC-LIKE PROFILE OF 4-PROPYL-2H-BENZO[H]-CHROMEN-2-ONE (FCS-304) IN MICE AND RATS](/img/en/prev.gif)







 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink