










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSuma Psicológica
versión impresa ISSN 0121-4381
Suma Psicol. v.17 n.2 Bogotá jul./dic. 2010
EFFECTS OF FREE-FOOD AND CONTINUOUS REINFORCEMENT SCHEDULE PRESENTATION ON LEVER PRESSING FOR FOOD BY RATS
EFECTOS DE PRESENTAR COMIDA LIBRE Y UN PROGRAMA DE REFORZAMIENTO CONTINUO SOBRE LAS PRESIONES A UNA PALANCA POR COMIDA EN RATAS
Raul Avila1, Juan C. Gonzalez1 and Patricia Miranda1
1School of Psychology National Autonomous University of Mexico, Mexico
* The authors wish to thank Dr. Carlos Bruner for the facilities given to conduct this experiment in his laboratory. The authors also wish to thank Dr. Gustavo Bacha and Belgica Porras for their priceless commentaries on a previous version of this manuscript.
Correspondence should be sent to: Raul Ávila
Laboratory of Experimental Analysis of Behavior
Facultad de Psicologia, Universidad Nacional Autonoma de Mexico
Edificio C, 2do. Piso. Cubículo 6, Av. Universidad 3004, Mexico, D.F., 04510, MEXICO.
Phone: 01-55-56222303. Email: raulas@servidor.unam.mx.
Recibido: Septiembre 1 2010 Aceptado: Diciembre 15 2010
Abstract
Six food-deprived rats were each exposed to a procedure in which lever pressing produced food in the presence and absence of free food. Although the highest frequency of lever pressing was observed when each lever press was reinforced with food in the absence of any supplementary free food, all six rats continued to press the lever when free food was available. The rats consumed approximately the same amount of food when free food was available, whether or not lever pressing was also reinforced. When each lever press was reinforced in the presence of free food, approaches to the free-food cup (and free-food consumption) decreased from the beginning to the end of the session, whereas lever pressing increased gradually across the session. These intra-session changes in behavior were interpreted in terms of the satiation of consummatory behavior.
Keywords: free food; continuous reinforcement; lever pressing; rats.
Resumen
Se expuso a seis ratas privadas de comida a un procedimiento en el cual las presiones a una palanca resultaron en la entrega de una bolita de comida en presencia o ausencia de comida disponible. Aún cuando se observó la frecuencia más alta de presión a la palanca durante la condición en la cual se reforzó cada presión en la ausencia de comida disponible, las seis ratas continuaron presionando la palanca cuando había comida disponible. Cuando se reforzaron las presiones a la palanca o éstas ocurrieron sin consecuencia alguna, las ratas consumieron aproximadamente la misma cantidad de comida cuando ésta estuvo disponible. Cuando se reforzó cada presión a la palanca en la presencia de la comida libre, las aproximaciones a la charola (y el consumo de esta comida) disminuyeron del principio al final de la sesión, mientras que las presiones a la palanca aumentaron gradualmente conforme transcurrió la sesión. Los cambios intra sesión de la conducta se interpretaron en términos de la saciedad de la conducta consumatoria.
Palabras clave: comida libre; reforzamiento continuo; presiones la palanca; ratas.
Jensen (1963) exposed food-deprived rats to a set of contingencies in which each lever press produced a food pellet (continuous reinforcement schedule, or CRF) for a predetermined number of food pellets. In the next experimental condition, food pellets were also available in a metal cup in the rear wall of the chamber, in addition to the pellets that could be earned by pressing the lever. Thus, in this second experimental condition there were two food sources that obtained after each lever press and that available in the cup. The rats consumed mainly free food from the cup but they also continued pressing the lever for food.
Subsequent research has demonstrated that lever pressing for food in the presence of free food occurs with intermittent schedules of reinforcement, including fixed-ratio and variable interval schedules. The phenomenon, known as contrafreeloading, also occurs in different strains of rats and in different species (cf. Inglis, Forkman & Lazarus, 1997; see Osborne, 1977 for a review of the generality of the phenomenon).
Several explanations of contrafreeloading have been proposed. It has been suggested that a) the stimulus changes associated with the operation of the food-pellet dispenser acquire a conditioned reinforcement function on lever pressing for food (Alferink, Crossman & Cheney, 1973), b) food reinforcement during the preliminary training produces neo-phobia to the free food source when the two food sources are simultaneously available (P. Mitchell & White, 1977), c) stimulus changes associated with food delivery are reinforcing by themselves and this sensorial reinforcement maintains lever pressing for food (Osborne & Shelby 1975), d) lever pressing is self-reinforcing (Jensen, 1963), and e) lever pressing for food in the presence of a free-food source is a kind of exploratory behavior (Singh, 1970).
Another possibility, not considered previously, is that the contrafreeloading effect may be a product of the training procedures employed in the previous studies. For example, in some studies, the subjects were first trained to lever press for food with a continuous reinforcement schedule and thereafter the frequency of lever pressing in the presence of free food was determined (e.g., Jensen, 1963; Neuringer, 1969). In other studies one or two sessions of exposure only to the free-food cup were alternated with one or two sessions of continuous reinforcement of lever pressing before exposing the subjects to the two food sources simultaneously (e.g., Carder & Berkowitz, 1970; D. Mitchell, Scott & Williams, 1973). The variations in the training procedure previously mentioned were used to train the subject to press the lever and/or to familiarize it to the free-food cup presence. Therefore, data regarding the effects of these preparatory manipulations on lever pressing for food in the presence of free food were not reported.
Besides the variations in the preliminary training conditions, subjects were sometimes exposed to the two food sources simultaneously during a different number of sessions, whose duration also varied between the studies. For instance, in some studies the subjects lever pressed for food in the presence of free food during one or two sessions of 30 minutes each (e.g., Jensen, 1963; Neuringer, 1969). In other studies the two food sources were available simultaneously during 20 sessions of 20 minutes each (e.g., Taylor, 1972; Tarte & Snyder, 1973) or during 70 or more 30min sessions (e.g., Davidson, 1971).
Differences in the preliminary training conditions and in the exposure time to the procedure between the previous studies, may contribute to the lack of a satisfactory explanation of contrafreeloading. To clarify the conditions responsible of the occurrence of the phenomenon in the present study we explored the separate effects of the variables involved in its occurrence (cf. Ruiz & Bruner, 2005) and the effects of filling the free-food cup with food-pellets and reinforcing each lever press by rats.
METHOD
Subjects
Six male Wistar rats without experimental history and three months old served as subjects. The rats were housed individually with free access to water and were food deprived to the 80% of their free feeding weight.
Apparatus
An experimental chamber for rats was used (Mod. BRS Foringer, 20705). The chamber was equipped with a lever (Hartmann) sensitive to 10 grams of force and was located in the center of the front wall of the chamber. A food tray was located 3 centimeters to the right of the lever into which 25 mg food pellets were delivered. The food pellets were remolded from food for rats. This tray was 2 cm wide by 2 cm long and 1.5 cm deep. A food dispenser (BRS/LVE Mod. PDC/PPD) was used to deliver the food pellets. Four centimeters to the left of the lever there was a 1.5 cm hole on the metal-wall of the chamber. Six millimeters behind this hole there was a metal connected to a water bottle and to a licking recorder built for this purposes. A bulb in the upper part of the front wall 12 cm from the floor of the chamber illuminated the chamber. A metal cup was located on the rear wall of the chamber. This metal cup was 6 cm wide by 10 cm long and 4 cm deep and remained empty or full with 600 food pellets, according to the experimental conditions of the study. In the following sections this metal cup will be referred to as the free-food cup. After 50 sessions, a photoreceptor was attached to the metal cup to register the times the subject introduced its mouth into the cup.
Procedure
The lever and the free-food cup were present in the experimental chamber under all condition. None of the rats had any previous training to lever press or to approach to the free-food cup. In the first experimental condition only lever pressing without programmed consequences was recorded. Thereafter, lever pressing without consequences continued to be recorded and the free-food cup was filled with 600 food pellets. In the third experimental condition the free-food cup full of food pellets was present and each lever press produced a food pellet in the food tray located to the right of the lever. In the next condition the CRF schedule for food was present but the free-food cup remained empty. In the next two experimental conditions the free-food cup was filled with food pellets either with or without the CRF schedule simultaneously available. In the last experimental condition only lever pressing without programmed consequences was recorded.
For rats R1, R2 and R3 each experimental condition was in effect for 30 one-hour sessions, except the first exposure to free food plus the CRF schedule, which was in effect during 60 one-hour sessions. For rats R4, R5 and R6 all experimental conditions were in effect during 30 one-hour sessions each.
For rats R1, R2 and R3 a metal tube was connected to a water bottle and was available during the entire experiment. Each lick to the tube produced a drop of water. For rats R4, R5 and R6 the metal tube was absent.
RESULTS
The 3 rats (R1, R2 and R3), for whom water was available in all experimental conditions, consumed between 8 and 10 milliliters of water per session in the operant level condition and between 3 and 4 ml of water in the next six conditions. This water intake did not alter the effects of the contingencies on rate of lever pressing or free-food intake. Therefore, the water consumption data have been omitted in the description of the results.
Figure 1 shows the number of lever presses by each rat in consecutive sessions of exposure to each experimental condition. In the first experimental condition (operant level), in which lever presses produced no programmed consequences, all six rats pressed the lever at least some of the time, albeit at relatively low levels. The number of lever presses observed in this operant-level condition served as a point of comparison to assess the effects on lever pressing of adding free food to the experimental chamber.

In the second experimental condition (free food), adding 600 food pellets to the free-food cup decreased lever pressing in all 6 rats. In the next condition (free food and CRF), in which the free-food cup was filled with food pellets and each lever press was reinforced with food, increased lever pressing for rats R2, R3, R4 and R6. For rat R1 lever pressing increased and then decreased across sessions in this condition. For rat R5 lever pressing increased slightly in comparison to the previous condition in which there was only free food. In the next condition (CRF), in which only each lever press produced food but free food was unavailable, lever pressing increased notably in all 6 rats, in comparison to that observed in presence of free food in the previous condition.
Replication of the free food and CRF condition produced an abrupt decrease in lever pressing to a level similar to that observed in the first exposition to this condition. Replicating the free-food condition resulted in a rate of lever pressing even lower than that observed in the simultaneous exposure to the two food sources. In the last condition of the experiment (operant level), in which lever pressing produced no programmed consequences and the free-food cup was empty, lever pressing decreased to a level comparable to that observed in the initial exposure to this condition.
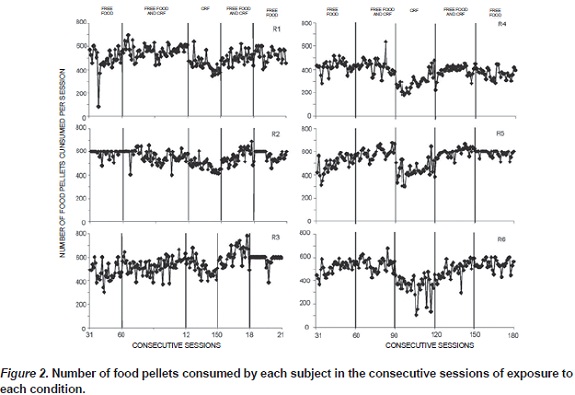
Figure 2 shows the total number of food pellets consumed per session across conditions for each rat. The dependent variable is the sum of pellets consumed from the free-food cup and the sum of the pellets earned by lever pressing. All 6 rats consumed approximately the same number of food pellets when free food was available, whether or not it was accompanied by the CRF schedule. When the CRF schedule was available in the absence of free food, food consumption was consistently lower that when free food was available.
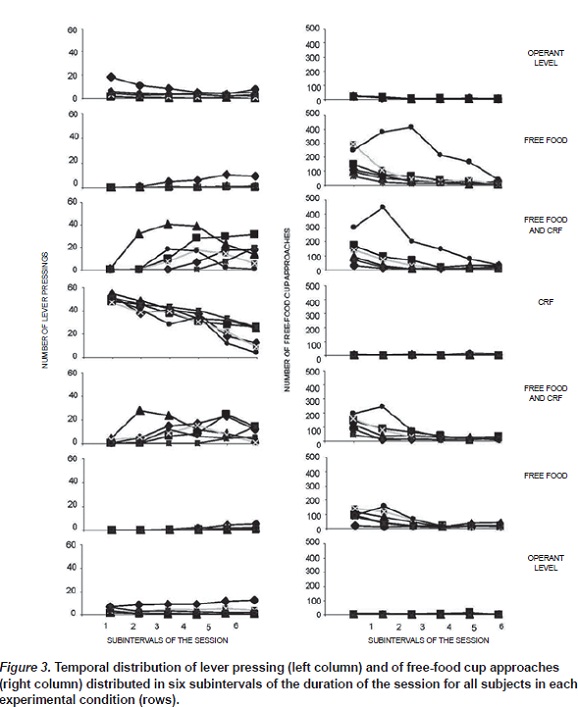
The temporal distribution of lever pressing was recorded across six 10-min intervals of the session. Figure 3 shows within-session frequencies of lever pressing (left panels) and free-food cup approaches (right panels) for the 6 rats across successive 10-min blocks in the session. These data are based on the final 5 sessions of exposure to each experimental condition. First, regarding lever pressing, the upper left panel of Figure 3 shows that in the operant level condition the frequency of lever pressing was very low at the beginning of the session, decreasing gradually until the end of the session. When free food was added to the free-food cup, lever pressing was infrequent at the beginning of the session and occurred more frequently toward the end of the session. When free food was accompanied by the CRF schedule for lever pressing (third left panel), this response occurred at a low frequency in the first 30 min, but increased gradually in the latter portions of the session. When the CRF schedule was available without free food, lever pressing was relatively high at the beginning of the session and decreased gradually toward the end of the session. Returning to the free-food plus CRF condition resulted in a similar pattern as that seen in the first exposure to this condition: relatively low frequency of lever pressing at the beginning of the session which increased as the time of the session elapsed. The other two replications also produced behavior similar to free-food pellets were consumed, the number of that seen in the initial exposures. times that the subjects interrupted the photorecep-Although in the present experiment it was tor located in the free-food cup provided an indicaimpossible to record the exact moment when the tor of free food consumption. As was mentioned before, the photoreceptor was located in the free food cup 50 sessions after the experiment began with rats R1, R2 and R3. Therefore, for these rats there are no data regarding the approaches to the free-food cup in the first operant-level condition.
Thus, in the upper right panel of Figure 3 only the temporal distribution of free-food approaches for rats R4, R5 and R6 are shown. For these three rats, the approaches to the free-food cup were relatively frequent at the beginning of the session and decreased towards the end of the session. The following panel shows that add free food to the cup resulted in that the six subjects consumed most of these free-food pellets at the beginning of the session and they consumed gradually less pellets as the session time elapsed. Adding the CRF schedule for lever presses produced little or no change in the temporal distribution of freefood consumption. In the absence of free food, approaches to the free-food cup were very low during the whole session. Adding free food to the cup while the CRF schedule was available generated essentially the same temporal distribution of free-food cup approaches as that observed in the first exposure to this condition. Replications of the other conditions produced similarly comparable performances as those seen in the initial exposure to these conditions.
DISCUSSION
The purpose of the present study was to explore the effects of free food access on the frequency of lever pressing in food-deprived rats. In general, rats pressed the lever in the presence of free food—the same food produced by lever pressing. This puzzling phenomenon, known most commonly as contrafreeloading, has been previously reported in the literature, though its parameters are not well understood. The goal of the present study was to examine thoroughly some of the variables of which contrafreeloading is a function. We will discuss below some of these variables in relation to specific experimental conditions. Thereafter, the relevance of these data to the studies on contrafreeloading with humans and work ethics with animals and humans will be discussed (e.g., Clement, Feltus, Kaiser & Zentall, 2000; Singh & Query, 1971; Zentall & Singer, 2007).
In the first two conditions, operant level and free food, a very low frequency of lever pressing was found, whether free food was available or not. This finding suggests that lever pressing by itself has little or no "intrinsic appeal," as was suggested by Jensen (1963). Adding free food to the cup did not affect the frequency of lever pressing without programmed consequences.
In the third experimental condition, free food and CRF, it was found that the frequency of lever pressing increased notably in comparison with the previous condition in which there was only free food in the cup. Because the rats had no previous training on the procedure, to find a substantial level of lever pressing in the presence of free food shows that a history of lever pressing is unnecessary to observe lever pressing in the presence of free food.
It has been suggested that the contrafreeloading phenomenon is transitory (e.g., Taylor, 1972), contrary to this suggestion, the present results show a substantial level of food-reinforced lever pressing in the presence of free food over 60 consecutive daily sessions. This finding shows that contrafreeloading is not transient, but rather, occurs consistently and over relatively long periods of time.
When the free-food cup was emptied and only the CRF schedule was available, it was found that the rate of lever pressing was at least two times higher than when the CRF schedule was accompanied by free food. When free food was again available, lever pressing returned to a level similar to that observed in the first exposure to the condition. This finding suggests that lever pressing for food in the presence of free food is relatively independent of history effects. This is, a substantial level of lever pressing in the presence of free food was obtained after 30 sessions of exposure only to the free food (second condition) or after a extensive exposition only to the CRF schedule (fourth condition). The replicability of the results also helps to rule out possible sequence effects as explanations of the effects.
In the literature on contrafreeloading, the main dependent variable reported is the rate of lever pressing in the presence of a cup filled with food. In the present study the amount of food consumed by each subject was also recorded. This dependent variable is of interest because the rats could consume different amounts of food per session when one or two different food sources were available. It was found that in general the rats consumed approximately the same amount of food when they had available only the cup filled with pellets or both food sources. The amount of food consumed per session in these conditions was higher that when only the CRF schedule was available. These findings are comparable with the capacity hypothesis advanced by DeMarse, Killeen & Baker (1999) to explain the intra-session changes of food-reinforced lever pressing by rats. In one of their experiments, the authors provided three methods to measure the capacity of the subjects, where capacity refers to the amount of food consumed. They exposed rats to a) a CRF schedule of lever pressing for food, b) a schedule of response-independent food, in which a food pellet was delivered at 15-s intervals irrespective of behavior, or c) free food in a cup (145 mg of food). They found that the rats consumed a relatively constant amount of food in the three sessions of exposure to each method of food presentation. However, the condition involving free food in the cup resulted in the highest amount of food consumed per session, and therefore provided the best estimation of the rats' capacity to consume food. In the present study, as in the DeMarse, et al. (1999) study, it was found that each rat consumed a relatively constant amount of food during the 30 sessions of exposure to each condition. The conditions of the present study in which free food was available in the context of the CRF schedule could be conceptualized as the combination of the two methods employed by DeMarse et al. Although there were two different food sources, the rats consumed approximately the same amount of food as when they had available only the cup filled with food pellets.
As was mentioned before, in the condition of the present experiment in which only the CRF schedule was available, the amount of food consumed per session was a little lower than when the two food sources or only the free food was available. This finding is also congruent with that reported by DeMarse, et al. (1999), who also found that the CRF schedule produced less food consumption than the free-food method. The authors suggested that this difference in the amount of food consumed per session between the two methods could be due to the difference in the time of access to food allowed by the two conditions. The free-food method provides free access to food and the CRF schedule requires a response to produce the food, and this requirement could limit the time of access to the food. It could be said that in the present experiment there was also a difference in the access time to free food and to the food pellet after each lever press. Therefore, it is possible that this difference in the access time to the two food sources could account for the differences in the amount of food consumed from the two food sources.
In the present study the frequency of lever pressing and the frequency of interruptions of a photoreceptor located in the free-food cup were recorded in successive blocks of 10-min each across the session for all experimental conditions. In the two experimental conditions in which free food was available in the absence of lever-pressing contingencies, food consumption was relatively high at the beginning of the session and decreased gradually as the session elapsed. In the two experimental conditions in which free food was available in the presence of the CRF schedule for lever presses, free-food consumption decreased gradually and lever pressing increased gradually from the beginning to the end of the session. In the experimental condition in which only the CRF schedule was available, lever pressing decreased monotonically from the beginning to the end of the session.
The gradual decrement of free-food consumption found in the four experimental conditions in which free food was available has no precedent in the literature on contrafreeloading. However, it is possible to relate this finding to studies from the operant conditioning literature on within-session changes in the rate of food-reinforced lever pressing. It has been suggested that within-session decreases in response rate are the result of the habituation to the food used as reinforcer (e.g., McSweeney & Roll, 1998). This explanation of free-food consumption may be viable; however, in the present study no manipulation typical of operant experiments on habituation was implemented and, therefore, the procedure and the findings of the present study hardly suggest that there was habituation of free-food consumption. An alternative and perhaps simpler explanation is that the within-session decrement of freefood consumption is the result of the satiation produced by the continuous availability of food (cf. Bizo, Bogdanov & Killeen, 1998; DeMarse, et al., 1999; Morgan, 1974). Satiation refers to the situation in which the subject will not emit a specific consummatory response (e.g., to eat) even though the appropriate incentive condition (e.g., the food) is present. The data from the present study are congruent with an explanation in terms of satiation given that the rats stopped consuming food pellets from the free-food cup even though there still was a substantial number of food pellets in the cup.
Along similar lines, Morgan (1974) has suggested a collateral effect of satiation is an increase in the general activity of the organism, similar to the increments in activity commonly observed after extinction of a conditioned operant. Therefore, the within-session increases in food-reinforced lever pressing could be the result of an increase in the general activity of the organism, including lever pressing. This explanation seems viable if we consider that in the two experimental conditions in which free food was available and lever pressing without programmed consequences was recorded, such lever presses occurred towards the end of the experimental session, after the rats stopped consuming free food. This very low level of lever pressing could be part of an increment of general activity as a result of the satiation of the consummatory behavior.
As said before, in this last section the relationship between the present results and those on contrafreeloading and work ethics with humans and animals will be discussed.
Although there are relatively few studies, it seems that contrafreeloading occurs reliably with humans. For instance, Singh (1970) exposed children to a choice between working for candies as rewards and obtaining them for free, and found that they preferred to work for the reward. In another study, Singh and Query (1971) tested if contrafreeloading could be related to the protestant ethics given that this kind of education emphasizes that people must work for rewards instead of obtaining them for free. To test this hypothesis, the authors exposed children with and without a protestant ethic education to the choice between working for candies and obtaining them for free. They found that regardless of the kind of education received all children worked to obtain the candies.
The results of the previous studies are promising, but insufficient for a description of the variables involved in the occurrence of the phenomenon with humans. Therefore, without the intention of an abrupt generalization from rats to humans, our procedure and results could be useful as an example of a strategy to find out the variables responsible of contrafreeloading in humans.
This strategy can be extended to the analysis of the procedure used to study work ethics with animals. Briefly, it has been shown that pigeons will peck a key to obtain a stimulus signaling a high response requirement for food more frequently than pecking another key which signals a comparatively lower response requirement (e.g., Clement, et al., 2000; Zentall & Singer, 2007). As in the case of the contrafreeloading procedure, the main procedure details involved in the study of work ethics can be analyzed one by one to clarify the variables responsible of this phenomenon.
REFERENCES
Alferink, L., Crossman, E., & Cheney, C. (1973). Control of responding by a conditioned reinforcer in the presence of free food. Learning & Behavior, 1, 38-40. [ Links ]
Bizo, L. A., Bogdanov, S. V. & Killeen, P. R. (1998). Satiation causes within-session decreases in instrumental responding. Journal of Experimental Psychology: Animal Behavior Processes, 24, 439-452. [ Links ]
Carder, B., & Berkowitz, K. (1970). Rats' preference for earned in comparison with free food. Science, 167, 1273-1274. [ Links ]
Clement, T. S., Feltus, J. R., Kaiser, D. H. & Zentall, T. R. (2000). "Work ethic" in pigeons: reward value is directly related to the effort or time required to obtain the reward. Psychonomic Bulletin and Rev, 7, 100-106. [ Links ]
Davidson, A. B. (1971). Factors affecting keypress responding by rats in the presence of free food. Psychonomic Science, 24, 135-137. [ Links ]
DeMarse, T. B., Killeen, P. R., & Baker, D. (1999). Satiation, capacity, and within-session responding. Journal of the Experimental Analysis of Behavior, 72, 407-423. [ Links ]
Inglis, I. R., Forkman, B., & Lazarus, J. (1997). Free food or earned food? A review and fuzzy model of contrafreeloading. Animal Behaviour, 53, 1171-1191. [ Links ]
Jensen, G. D. (1963). Preference bar pressing over free-loading as a function of number of unrewarded presses. Journal of Experimental Psychology, 65, 451-454. [ Links ]
McSweeney, F. K., & Roll, J. M. (1998). Do animals satiate or habituate to repeatedly presented reinforcers? Psychonomic Bulletin and Review, 5, 428-442. [ Links ]
Mitchell, D., Scott, D. W., & Williams, K. D. (1973). Container neophobia and the rat's preference for earned food. Behavioral Biology, 9, 613-624. [ Links ]
Mitchell, P., & White, K. G. (1977). Responding in the presence of free food: differential exposure to the reinforcement source. Bulletin of the Psychonomic Society, 10, 121-124. [ Links ]
Morgan, M. J. (1974). Resistance to satiation. Animal Behaviour, 22, 449-466. [ Links ]
Neuringer, A. J. (1969). Animals respond for food in the presence of free food. Science, 166, 399-401. [ Links ]
Osborne, S. R. (1977). The free food (contrafreeloading) phenomenon: A review and analysis. Learning & Behavior, 5, 221-235. [ Links ]
Osborne, S. R. & Shelby, M. (1975). Stimulus change as a factor in response maintenance with free food available. Journal of the experimental analysis of behavior, 24, 17-21. [ Links ]
Ruiz, J. A. & Bruner, C. A. (2005). Transformación de un programa de intervalo fijo de reforzamiento con agua en un procedimiento de beber inducido por el programa. Revista Mexicana de Análisis de la Conducta, 31, 47-65. [ Links ]
Singh, D. (1970). Preference for bar pressing to obtain reward over freeloading in rats and children. Journal of Comparative and Physiological Psychology, 73, 320-327. [ Links ]
Singh, D. & Query, W. T. (1971). Preference for work over "freeloading" in children. Psychonomic Society, 24, 7-79. [ Links ]
Tarte, R. D. & Snyder, R. L. (1973). Some sources of variation in the bar-pressing versus freeloading phenomenon in rats. Journal of Comparative and Physiological Psychology, 84, 128-133. [ Links ]
Taylor, G. T. (1972). A limitation of the contrafreeloading phenomenon. Psychonomic Science, 29, 173-174. [ Links ]
Zentall, T. R., & Singer, R. A. (2007). Within-trial contrast: pigeons prefer conditioned reinforcers that follow a relatively more rather than a less aversive event. Journal of the Experimental Analysis of Behavior, 88, 131-149. [ Links ]

