










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSuma Psicológica
Print version ISSN 0121-4381
Suma Psicol. vol.18 no.1 Bogotá Jan./June 2011
Fundamentos biológicos en la extracción de regularidades del habla
Biological foundations in the extraction of regularities from speech
Juan Manuel Toro1
1Unidad de Cognición y Cerebro, Universitat Pompeu Fabra, España
Este trabajo fue financiado por el programa Ramón y Cajal (RYC-2008-02909) y por el proyecto PSI2010-20029 del Ministerio de Ciencia y Educación.
La correspondencia relacionada con este articulo debe ser dirigida a Juan Manuel Toro
DTIC - Universitat Pompeu Fabra, C. Roc Boronat, 138, CP. 08018, Barcelona, España
Correo electrónico: juanmanuel.toro@upf.edu.
Recibido: Noviembre 4 2010 Aceptado: Marzo 8 2011
Resumen
Uno de los mayores problemas en el estudio del lenguaje es la comprensión de sus fundamentos biológicos. Una forma de abordar este problema que ha resultado efectiva es la de estudiar hasta qué punto compartimos con otros animales algunos de los mecanismos responsables de su procesamiento. En este artículo se presentan una serie de experimentos que demuestran que las habilidades para extraer regularidades rítmicas y estadísticas del habla están ya presentes en algunos roedores. A su vez, otros estudios sugieren que los animales también pueden procesar ciertas estructuras abstractas tradicionalmente ligadas a la gramática. Estos estudios apuntan en la dirección de que muchos de los componentes básicos del procesamiento lingüístico son compartidos a través de diferentes especies animales. Sin embargo, existe todavía un amplio abanico de fenómenos lingüísticos que deben ser estudiados desde una perspectiva comparada para determinar hasta qué punto el lenguaje humano hace uso de habilidades ya existentes en otros animales.
Palabras clave: percepción del habla; evolución del lenguaje; ratas; prosodia; gramática.
Abstract
One of the major problems in the study of language is to understand its biological roots. An effective way to tackle this issue is to explore to which extent we share with other animals some of the mechanisms responsible for its processing. In this paper we present a series of experiments that demonstrate that the abilities required to extract rhythmic and statistical regularities from speech are already present in rodents. Other studies also suggest that some animals might be able to process certain abstract structures that have been traditionally linked to grammar. Together, these studies point towards the idea that many of the basic components of language processing are shared across different species. Nevertheless, there is still a wide array of linguistic phenomena that should be studied from a comparative perspective to determine the extent to which human language uses abilities found in other animals.
Keywords: speech perception; language evolution; rats; prosody; grammar.
A mediados del siglo XIX la Sociedad Lingüística de París prohibió discusiones sobre el origen del lenguaje. Las especulaciones que se hacían sobre sus fundamentos biológicos y sus posibles precursores en otros animales no aportaban ninguna idea relevante y se consideró que era mejor dejarlas de lado (véase Hauser, Barner & O'Donnell, 2007). Afortunadamente, la situación ha cambiado, especialmente en los últimos años. Estudios empíricos hechos desde una perspectiva comparada han contribuido a replantear ideas sobre la naturaleza de los procesos involucrados en el procesamiento lingüístico, a la vez que han ayudado a contestar algunas preguntas sobre los orígenes de esta capacidad típicamente humana. En el mismo sentido, diferentes cambios en los niveles metodológico y conceptual han permitido un diálogo más fluido entre disciplinas tradicionalmente alejadas como la biología y la lingüística, pasando por la psicolingüística, la psicología comparada y la psicología del desarrollo. En la actualidad, los estudios realizados con animales son reconocidos cada vez con más fuerza como una fuente valiosa de información sobre la adquisición y procesamiento del lenguaje.
El lenguaje humano es un sistema de comunicación muy eficiente y los procesos que llevan a su adquisición son representativos de los retos cognitivos a los que nos enfrentamos los humanos desde el nacimiento. Antes del primer año de edad, se comienza a desarrollar en el bebé un sistema de comunicación que le permitirá expresar un número infinito de ideas basándose en la combinación de un número relativamente pequeño de sonidos (en el caso del habla, o de gestos en el caso del lenguaje de señas). Sin ningún tipo de instrucción explícita se aprenden no sólo centenares de palabras, sino también los sonidos propios de la lengua materna y las reglas para combinar de manera adecuada estos sonidos para formar palabras y estas palabras para construir frases.
En los humanos, el lenguaje se desarrolla de forma natural bajo condiciones normales. Sin embargo, resulta llamativo el hecho de que aparentemente el lenguaje no tiene paralelos en otros sistemas de comunicación animal. Algunas especies de aves tienen cantos formados por complicadas secuencias de sonidos (Doupe & Kuhl, 1999), y algunas especies de monos pueden informar sobre la presencia de depredadores específicos a otros miembros de su grupo (Zuberbuhler, 2006). A su vez, otros animales como los delfines parecen poder comunicarse puntualmente entre individuos (Janik, Sayigh & Wells, 2006). Pero, hasta el momento no se ha documentado que ninguno de estos sistemas de comunicación animal se equipare al lenguaje humano en su capacidad de transmitir información. Más concretamente, el lenguaje humano difiere de los otros sistemas de comunicación animal en su forma de utilizar un número finito de elementos para generar un rango infinito de expresiones (Hauser, Chomsky & Fitch, 2002). La pregunta entonces es ¿cómo puede un bebé desarrollar este complejo sistema de comunicación que es el lenguaje? La clave para resolver esta cuestión parece estar en el estudio de los mecanismos involucrados en la extracción inicial de información de los estímulos lingüísticos.
Existen tres niveles de análisis de la señal del habla que son especialmente relevantes en la adquisición temprana de estructuras lingüísticas: el prosódico, el de la frecuencia con que aparecen ciertos elementos en el habla (conocidas como regularidades distribucionales), y el del aprendizaje de reglas abstractas ligadas a la gramática. El procesamiento de la señal del habla a estos tres niveles parece ser fundamental en el proceso de adquisición de una lengua por parte de los bebés humanos. Sin embargo, como intentaré demostrar a continuación, investigaciones recientes hechas desde una perspectiva comparada muestran que podemos encontrar procesos muy similares en otros animales. Lo que esto sugiere es que varios de los fundamentos cognitivos que son importantes en la aparición de una habilidad típicamente humana, como es el lenguaje, son compartidos a través de diferentes especies.
El uso de claves prosódicas
La prosodia es una combinación de ritmo y entonación que se encuentra en el habla. El ritmo a su vez se define en función de la estructura silábica de una lengua y de su distribución relativa de consonantes y vocales (Ramus, Nespor & Mehler, 1999). Estudios en psicolingüística hacen cada vez más énfasis en la importancia que tienen las claves prosódicas en el procesamiento lingüístico en todos los niveles. Los bebés son sensibles a cambios en la prosodia desde muy temprana edad (Gleitman & Wanner, 1982) y esta habilidad podría ayudar en la adquisición temprana de algunos patrones gramaticales del lenguaje (Christophe, Guasti, Nespor, Dupoux & Van Ooyen, 1997). Más concretamente, el contorno prosódico de una frase puede señalar una característica sintáctica de las lenguas tan importante como su orden de palabras (el orden canónico que se sigue en una lengua para construir frases, que puede ser SVO (Sujeto-Verbo-Objeto) o SOV (Sujeto-Objeto-Verbo); (véase Nespor, Shukla, Vijner, Avesani, Schraudolf & Donati, 2008).
Un estudio comparando el comportamiento de bebés humanos y monos tamarines encontró que las dos especies podían utilizar el ritmo lingüístico para discriminar entre diferentes lenguas (Ramus, Hauser, Miller, Morris & Mehler, 2000). Sin embargo, no se podía determinar hasta qué punto esta habilidad podía encontrarse en especies diferentes a los primates. Más aún, no se sabía hasta qué punto el uso de claves prosódicas dependía de la capacidad para producir vocalizaciones complejas como lo hacen los humanos y los monos tamarines. Para resolver estas cuestiones desarrollamos un estudio en el que pusimos a prueba la capacidad de discriminar lenguas con el uso del ritmo lingüístico en ratas (Toro, Trobalon & Sebastian-Galles, 2005).
En estos experimentos se utilizaron 40 frases en holandés y japonés, dos lenguas que difieren radicalmente en su ritmo lingüístico. Todas las frases fueron grabadas por hablantes nativos de las dos lenguas y su duración media era de 2.5 segundos. En el Experimento 1 se presentaron a las ratas frases naturales, esto es, tal como fueron grabadas por los hablantes y sin ningún tipo de modificación acústica. En el experimento participaron 16 ratas Long Evans de 3 meses de edad.
Antes de comenzar el experimento, se entrenó a las ratas a presionar la palanca para obtener comida (una bola de leche azucarada). Una vez alcanzaron una tasa de respuesta adecuada, se comenzó el entrenamiento en discriminación de lenguas. Las ratas fueron divididas en dos grupos: el Grupo holandés que recibiría refuerzo por las frases en holandés, y el Grupo japonés que recibiría refuerzo por las frases en japonés. El experimento consistió en dos fases. Una fase de entrenamiento y una de prueba. La fase de entrenamiento tuvo 20 sesiones, una sesión por día. En cada sesión se presentaban a través de un altavoz 8 frases en holandés y 8 frases en japonés de forma balanceada. Luego de la presentación de cada frase había un intervalo de 2 minutos. Si la frase presentada era de la lengua reforzada (holandés para el Grupo holandés y japonés para el Grupo japonés), la rata recibía comida por sus presiones de palanca bajo un programa de razón variable más o menos diez (RV-10) durante todo el intervalo. Por el contrario, si la frase presentada era de la lengua no reforzada, la rata no recibía comida durante el primer minuto del intervalo, sin importar sus presiones de palanca. Para evitar una extinción de la respuesta, la rata sí podía recibir comida por presionar la palanca durante el segundo minuto del intervalo.
Después de las 20 sesiones de entrenamiento se hizo una sesión de prueba. Esta sesión era similar a las anteriores excepto por dos diferencias. Primero, se presentaban nuevas frases de holandés y de japonés, que no se habían presentado durante el entrenamiento. Se utilizaron frases nuevas para evaluar si las ratas podían generalizar lo aprendido durante el entrenamiento a nuevas instancias. Segundo, se desconectaron los dispensadores de comida. Así, las ratas no recibían comida ante ninguna de las frases. Con esto se controlaba la posibilidad de que diferencias en las tasas de presión de palanca fueran solo una consecuencia de un refuerzo inicial durante el intervalo de dos minutos, y que no tuvieran nada que ver con la discriminación efectiva de los estímulos. Para normalizar a través de diferencias individuales en las presiones de palanca, se calculó una tasa de discriminación.
Los resultados en la sesión de prueba del Experimento 1, en el que se presentó a las ratas frases naturales de dos lenguas diferentes, mostraron que las ratas pueden extraer suficiente información de la señal acústica como para poder discriminar entre ellas. Esto es, hubo una mayor frecuencia de presiones de palanca luego de frases de la lengua reforzada, que luego de frases de la lengua no reforzada (véase la Figura 1). Esta discriminación es posible luego de un entrenamiento relativamente corto y se observa a pesar de que durante la sesión de prueba las ratas no recibían ningún refuerzo. Más importante aún es el hecho de que esta discriminación se generaliza a nuevas instancias no presentadas durante la primera fase del experimento.
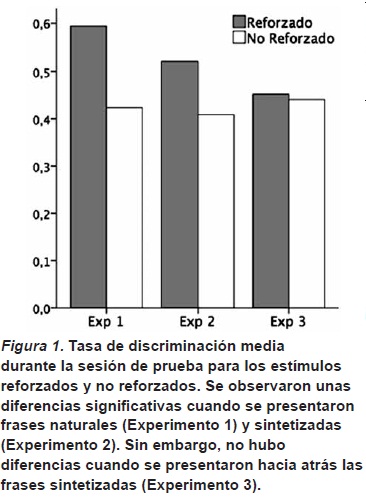
Estos datos, sin embargo, no permiten concluir que las ratas estuvieran utilizando necesariamente las claves prosódicas para la discriminación de lenguas. El habla natural contiene otras claves que pueden ayudar a diferenciar una lengua de otra. Por ejemplo, está el repertorio fonético, esto es el conjunto de sonidos que pertenecen a una lengua y no a otra. Es posible que las ratas se estuvieran basando en solo unos pocos sonidos para discriminar el holandés del japonés. Para evaluar esta posibilidad hicimos el Experimento 2, en el que presentamos a las ratas frases sintetizadas de tal forma que sólo se conservaba la prosodia de las dos lenguas.
En el Experimento 2 se presentaron las mismas frases que en el Experimento 1, pero luego de ser re-sintetizadas. El objetivo de esta síntesis era eliminar todas las claves fonéticas que pudieran servir en la discriminación de las lenguas. Así, eliminando estas claves, sólo quedaba el ritmo lingüístico como fuente de discriminación. Para la síntesis de las frases originales se siguió un procedimiento conocido como saltanaj. En este procedimiento se reemplazan todas las vocales en las frases originales con /a/, todas las consonantes fricativas con /s/, las laterales con /l/, las oclusivas con /t/, las nasales con /n/ y las africadas con /j/. Así se mantiene la estructura y la entonación original de la lengua, pero se normalizan las diferencias fonéticas, al usar exactamente los mismos fonemas en todas las frases.
El procedimiento en el Experimento 2 fue el mismo que en el Experimento 1; la única diferencia fue la presentación de frases sintetizadas, en vez de frases naturales. Los resultados durante la sesión de prueba fueron similares a los observados en el Experimento 1. Esto es, las ratas mostraron una mayor frecuencia de respuestas para frases pertenecientes a la lengua reforzada que para frases de la lengua no reforzada (véase la Figura 1). Lo que sugieren estos resultados es que las ratas detectaron las diferencias en ritmo lingüístico que caracterizan el holandés y el japonés y las usaron para discriminar entre ellas. El hecho de que este resultado se observe con frases sintetizadas excluye la posibilidad de que la discriminación se haga con base en otras claves, como las fonéticas.
Estos resultados con ratas revisten especial importancia por su similitud con lo observado en bebés humanos y monos tamarines (Ramus et al., 2000). A través de las tres especies se observó que podían detectar y utilizar la prosodia del habla para distinguir dos lenguas diferentes, totalmente desconocidas. A su vez, cuando se eliminaba esta información prosódica al presentar los estímulos invertidos, ninguna de las tres especies podía hacer la discriminación. Este patrón sugiere que las tres especies están procesando de forma similar los estímulos. Esto es, que la capacidad para procesar los aspectos prosódicos del habla que ha sido documentada en bebés también está presente en otros mamíferos tan alejados filogenéticamente de los humanos, como son las ratas.
Aprendizaje estadístico
Además de las claves prosódicas, otra fuente potencial de información sobre la estructura de un lenguaje se encuentra en la forma como se distribuyen los fonemas, sílabas y palabras dentro de éste. Por ejemplo, en español, la vocal /a/ es más frecuente que la vocal /u/; su distribución a través de la lengua es diferente. A su vez, si en una frase encontramos el pronombre /nosotros/, lo más probable es que le siga un verbo, no un sustantivo (como en nosotros diseñamos el experimento), porque la estructura sintáctica del español marca preferencialmente un orden de tipo Sujeto-Verbo-Objeto. Así, un mecanismo que pueda detectar este tipo de regularidades en la distribución de los diferentes elementos que componen un lenguaje permitiría descubrir muchos aspectos de su estructura.
Saffran, Aslin & Newport (1996) demostraron que bebés de tan sólo 8 meses de edad podían extraer este tipo de información del habla. Los investigadores familiarizaron durante dos minutos a los bebés con un lenguaje artificial. Este lenguaje estaba compuesto por cuatro palabras trisilábicas sin sentido. Lo más importante de este lenguaje inventado es que no había ninguna pausa, ni ninguna otra señal acústica que señalara donde terminaba una palabra y donde comenzaba la otra. La única información que señalaba cuáles eran las palabras del lenguaje artificial era la distribución de las sílabas en éste. A pesar de lo difícil de la tarea, los investigadores encontraron que los bebés registraban esta distribución de sílabas y luego identificaban las palabras que componían el lenguaje presentado en función de esta distribución. A este tipo de aprendizaje se le llamó aprendizaje estadístico. Este aprendizaje estadístico despertó un enorme interés porque abría las puertas a la posibilidad de demostrar que un mecanismo general de asociación entre elementos diferentes (en esta caso, sílabas) estaba implicado directamente en tareas muy especializadas, como la segmentación del habla. De hecho, numerosos estudios posteriores han demostrado que el aprendizaje estadístico se extiende a través de modalidades (visual, auditiva, táctil) y de estímulos (fonemas, sílabas, tonos, ruidos, dibujos, figuras sin sentido, etcétera; para una revisión reciente, véase Aslin & Newport, 2008). Al ser un mecanismo tan general, aparece pues la pregunta si las ratas también podrían hacer uso de él para extraer regularidades del habla en forma de distribuciones de sílabas tal como lo hacen los bebés y los adultos humanos.
Siguiendo el diseño de Saffran y colaboradores (1996), en este experimento se creó un lenguaje artificial compuesto por cuatro palabras trisilábicas sin sentido (e.g., tupiro, golabu, bidaku, padoti). Para una descripción más detallada del estudio, véase a Toro y Trobalon, 2005. Estas palabras estaban formadas por doce sílabas diferentes y cada una de ellas se sintetizó usando el conversor de texto a habla MBROLA (disponible en http://tcts.fpms.ac.be/synthesis/), con la base de difonos españoles es2. La duración de cada sílaba fue de 232 ms. Lo más importante es que la frecuencia fundamental de cada sílaba se mantuvo constante a 200 Hz, así que no había cambios acentuales que marcaran ninguna sílaba. Las cuatro palabras se concatenaron luego de forma que no existiera ningún silencio entre ellas ni ninguna otra señal acústica que marcara el comienzo o el final de cada palabra. Así, la única fuente de información que distinguía una palabra de otra era la secuencia exacta de las sílabas que la formaban. Esto es, la información de la distribución de las sílabas en el lenguaje artificial. Por ejemplo, en la palabra tupiro la probabilidad de que después de la sílaba /tu/ venga /pi/ es de 1.0 (siempre que aparezca una, vendrá inmediatamente después la otra). Lo mismo de la sílaba /pi/ a /ro/. Sin embargo, la probabilidad de que después de la sílaba / ro/ venga cualquier otra sílaba es de solo 0.33. Esto es porque después de tupiro puede venir cualquiera de las otras 3 palabras del lenguaje artificial, con lo que la probabilidad de que a una sílaba final de una palabra siga una sílaba específica de otra palabra desciende a sólo un tercio (véase la Figura 2). Es precisamente esta caída en las probabilidades de una sílaba a la siguiente la que marca el final y el comienzo de una palabra en ausencia de claves acústicas. En ausencia de claves acústicas que separen las palabras, se ha demostrado que un oyente es capaz de extraer la secuencia de sílabas de cada palabra usando su distribución en el lenguaje artificial (Saffran et al., 1996).

En este experimento se usaron 32 ratas Long Evans de 3 meses de edad. Antes de comenzar el experimento se entrenó a las ratas a presionar una palanca para recibir comida. Una vez las ratas alcanzaron tasas de respuestas adecuadas se comenzó el experimento que consistía en dos fases, una de familiarización y otra de prueba. Cada fase tenía sólo una sesión. En la sesión de familiarización se dejaba que la rata presionara la palanca y se le daba comida bajo un programa de refuerzo de razón variable más o menos 10 (RV-10). De forma importante, durante toda esta sesión, que duraba 20 minutos, se presentaba auditivamente a la rata el lenguaje artificial. Al día siguiente se realizaba la sesión de prueba.
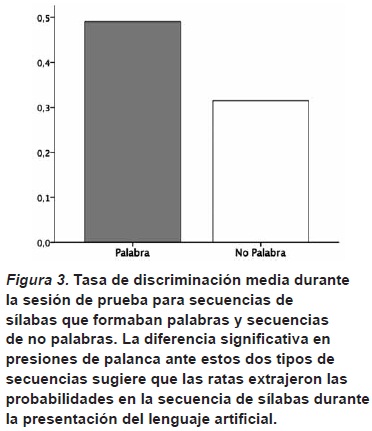
En ésta, los dispensadores de comida estaban desconectados con lo que las ratas no recibían refuerzo por sus presiones de palanca. Durante la sesión de prueba se presentaban tres sílabas de las que formaban el lenguaje artificial. Estas sílabas podían formar una palabra (p.e., tupiro), o ser la última sílaba de una palabra unida a las dos primeras sílabas de otra (p.e., rogola). Luego de la presentación de estas tres sílabas había un intervalo de un minuto. Durante este intervalo se registraban las presiones de palanca de la rata. Al finalizar el minuto de intervalo se presentaban otras tres sílabas y se volvía a registrar las presiones de palanca.
Los resultados mostraron que había una mayor frecuencia de presiones de palanca durante el intervalo posterior a la presentación de sílabas que formaban una palabra, que de las que no la formaban (véase la Figura 3). Esto sugiere que las ratas asociaron el refuerzo recibido durante la sesión de familiarización con las palabras que formaban el lenguaje artificial. Como estas palabras se presentaron durante la familiarización como un flujo constante de sílabas, sin separación entre ellas, la única forma de separarlas era detectando la información en términos de probabilidades estadísticas. Por lo tanto, estos resultados apoyan la idea de que la capacidad para detectar este tipo de información en un flujo continuo de habla está presente en estos roedores. Así, el mecanismo fundamental que pueden utilizar los bebés para segmentar inicialmente palabras del habla está ya presente en otras especies que no poseen un sistema de comunicación acústico especialmente complejo.
Extracción de estructuras abstractas
Una de las características más importantes del lenguaje es su gramática, esto es, la forma como se estructuran fonemas, sílabas y palabras para crear frases con sentido. La mejor forma de describir los aspectos gramaticales del lenguaje es a través de estructuras más o menos abstractas (es decir, no ligadas directamente a elementos específicos). Por ejemplo, en inglés, la forma de construir el pasado de un verbo regular es agregarle el sufjo /ed/ al final del infinitivo (como en /worked/ o /played/), lo que se puede describir en términos de X-ed (siendo X cualquier verbo regular). En español, la estructura por defecto de una frase afirmativa es SVO. Una vez hemos aprendido esta regla, podemos reemplazar las variables con casi cualquier palabra y tenemos una frase regular en español (p.e., la rata presionó la palanca, Ana escribió un artículo, la gramática es compleja). Diferentes estudios han demostrado que los bebés pueden aprender ciertas reglas simples a partir de la presentación de una serie de palabras que siguen la regla (Marcus, Vijayan, Bandi Rao & Vishton, 1999). Pero si los límites en los sistemas de comunicación animal se encuentran en su incapacidad de hacer uso de este tipo de estructuras abstractas, es posible que otros animales no puedan llegar a procesarlas de forma similar a los humanos.
Con el fin de estudiar este tema realizamos un experimento sobre la posibilidad de aprendizaje de reglas lingüísticas simples en ratas. En este experimento creamos una serie de 16 palabras trisilábicas que seguían una regla de tipo AAB (donde A y B son diferentes sílabas). Esto significa que la primera y la segunda sílaba siempre eran iguales y la tercera sílaba era diferente (p.e., lalabe, dodotu, kikina). Las palabras fueron sintetizadas de igual forma que las descritas en el experimento anterior de aprendizaje estadístico. El procedimiento también fue el mismo que el utilizado en los experimentos descritos anteriormente.
El experimento tenía dos fases, una de familiarización y otra de prueba. Cada fase tenía una sesión de entrenamiento en que la rata podía presionar la palanca para obtener comida bajo un programa de refuerzo RV-10. Durante este tiempo se presentaban acústicamente las palabras con pausas de 500 milisegundos entre palabras. Esta sesión duraba 20 minutos. Al día siguiente se hacía la sesión de prueba en la cual los dispensadores de comida estaban desconectados; se presentaban nuevas palabras, formadas por nuevas sílabas (no presentadas durante la familiarización) que podían seguir la misma regla que las de la familiarización (AAB), o una regla diferente (p.e. ABA). Las palabras de la prueba se presentaban con un intervalo de un minuto entre ellas. Durante este intervalo se registraban las presiones de palanca de las ratas.
Los resultados no mostraron ninguna diferencia en presiones de palanca luego de nuevas palabras que seguían la regla de familiarización y palabras que no la seguían (Toro & Trobalon, 2005). Esto es, en este experimento no obtuvimos ninguna evidencia de que las ratas hubieran extraído la regla abstracta que era común a las palabras durante la sesión de familiarización. ¿Significa esto que los roedores no puede aprender este tipo de reglas? Es poco probable, pues hay indicios que sugieren que otras especies animales sí pueden extraer y generalizar reglas simples del tipo AAB.
Murphy, Mondragón y Murphy (2008) observaron que, luego de un entrenamiento muy prolongado, las ratas llegaban a discriminar secuencias de tres tonos que seguían determinada regla, de secuencias que no la seguían. Por su parte, Hauser y Glynn (2009) reportaron que los monos rhesus aprenden reglas similares si se presentan compuestas por vocalizaciones propias de la especie. Parece pues que el aprendizaje de este tipo de estructuras abstractas sí se puede observar en otras especies de animales, pero que para hacerlo se necesitan procedimientos más intensivos y características especiales en los estímulos para lograrlo.
En los últimos años, sin embargo, se han realizado numerosos esfuerzos para explorar el punto hasta el que otras especies de animales pueden aprender estructuras abstractas, cada vez más complejas, relacionadas con formas gramaticales del lenguaje. Herbranson y Shimp (2008) entrenaron durante más de 180 días a un grupo de palomas en la discriminación de una gramática artificial compuesta por entre 5 y 7 elementos. Los resultados sugerían que luego de un entrenamiento masivo, las palomas podían discriminar secuencias que seguían las reglas de esa gramática artificial de las que no lo hacían. Otras series experimentales han mostrado que aves con cantos muy elaborados (como los pinzones cebras) pueden aprender gramáticas un poco más complejas con menos entrenamiento (véase Gentner, Fenn, Margoliash & Nusbaum, 2006; Van Heijningen, De Visser, Zuidema & Ten Cate, 2009). A su vez, se ha observado que el aprendizaje de ciertas reglas en los chimpancés se guía por principios similares a los observados en humanos (Endress, Carden, Versace & Hauser, 2010), y que monos tamarines pueden aprender reglas morfológicas simples como la afjación (Endress, Cahill, Block, Watumull & Hauser, 2009). Estos resultados sugieren que otras especies animales pueden llegar a procesar algunas estructuras tradicionalmente ligadas a la organización gramatical del lenguaje. Ya que los sistemas de comunicación de estas especies no utilizan este tipo de estructuras, a lo que apuntan estos estudios es que el lenguaje humano hace uso de una serie de mecanismos preexistentes que muy probablemente no aparecieron como respuesta a necesidades comunicativas. En la actualidad, la investigación comparada está orientada hacia la dirección de comprender los límites que tiene el aprendizaje de estructuras complejas en otros animales y así explorar en qué medida el lenguaje hace uso de mecanismos ya presentes en otras especies.
Posibles Diferencias entre Humanos y otros animales
A través de este artículo he intentado mostrar cómo una aproximación comparada al estudio del lenguaje puede ayudar a comprender mejor algunos de sus aspectos durante la adquisición y el procesamiento. He querido resaltar las coincidencias que se han encontrado entre humanos y otros animales en su capacidad para extraer información de las claves prosódicas del habla y de la distribución de elementos dentro de ella. Sin embargo, estos son sólo dos ejemplos de casos donde se han encontrado similitudes entre especies. Queda por fuera del alcance del presente artículo hacer una revisión más detallada de todos los casos que apuntan a este tipo de similitudes. De todas maneras, es interesante observar cómo los estudios con animales no sólo han servido para mostrar que los humanos compartimos con otras especies determinados mecanismos utilizados en el procesamiento del lenguaje. Otras investigaciones apuntan a algunas divergencias. Un ejemplo reciente se encuentra en los estudios sobre diferencias funcionales entre consonantes y vocales.
Estudios con humanos sugieren que tendemos a asignar distintos roles a las consonantes y a las vocales, (p.e. Bonatti, Peña, Nespor & Mehler, 2005; Toro, Nespor, Mehler & Bonatti, 2008) y que estas diferencias aparecen pronto en el desarrollo (Pons & Toro, 2010). Por el contrario, resultados con monos indican que para éstos las consonantes y las vocales son procesadas de manera equivalente (Newport, Hauser, Spaepen & Aslin, 2004). A pesar de que no se ha hecho todavía un estudio sistemático de este tema, los datos que se tienen hasta el momento apuntan hacia la idea que una vez se posee un sistema lingüístico, se da una transformación en el procesamiento de los estímulos que caen dentro del campo de este sistema. Así, en los humanos, las consonantes y las vocales dejan de ser estímulos equivalentes, como lo son para otros animales, y cada uno de estos elementos fonológicos adquiere un rol especial. Es posible pues que muchos de los mecanismos implicados en el procesamiento lingüístico sean compartidos a través de las especies, pero una vez estos mecanismos han dado lugar al lenguaje propiamente dicho, éste transforma la forma de procesar determinados estímulos.
Otro ejemplo de un aspecto del lenguaje potencialmente interesante para estudios comparados es la capacidad para crear ciertas frases subordinadas. Si se toma una frase como El experimento salió mal, se puede poner otra frase dentro como El experimento, aquel del que hablamos ayer, salió mal y la frase original seguirá siendo comprensible. Este tipo de frases resaltan el carácter jerárquico y no lineal del lenguaje y es una cuestión abierta el punto hasta el cual otras especies pueden procesar secuencias que tengan este tipo de características. Se necesitan experimentos con otras especies que resuelvan esta cuestión.
Así, la investigación con animales de procesos relacionados con el lenguaje ha aportado datos novedosos sobre características que hasta hace poco se consideraban únicamente humanas. Una de sus contribuciones más importantes ha sido la demostración que podemos encontrar en otros animales algunos de los mecanismos que los humanos utilizamos en la extracción de información del habla. Más específicamente, se ha demostrado que la habilidad para usar las claves prosódicas y estadísticas presentes en el habla está ya presente en algunos roedores. Por lo tanto, gracias a la investigación comparada se ha dado un fundamento empírico a la hipótesis de que los humanos utilizamos con fines lingüísticos, algunas habilidades ya existentes en otros animales. Sin embargo, como se señaló anteriormente, existe todavía un amplio abanico de temas relacionados con la adquisición y el procesamiento del lenguaje a estudiar desde una perspectiva comparada. La complejidad propia del lenguaje y la idea de que su aparente exclusividad pueda ser explicada en función de una serie de mecanismos compartidos con otros animales, deja la puerta abierta a un campo entero de investigación. Gracias a estudios experimentales que señalen los puntos que compartimos con otras especies y aquellos en los que nos distanciamos, tenemos la oportunidad de ampliar nuestra comprensión del lenguaje y de los mecanismos involucrados en su adquisición.
Referencias
Aslin, R. & Newport, E. (2008). What statistical learning can and can't tell us about language acquisition. In J. Colombo, P. McCardle & L. Freund (Eds.), Infant pathways to language: Methods, models, and research directions. Mahwah, NJ: Erlbaum. [ Links ]
Bonatti, L. L., Peña, M., Nespor, M. & Mehler, J. (2005). Linguistic constraints on statistical computations: The role of consonants and vowels in continuous speech processing. Psychological Science, 16, 451-459. [ Links ]
Christophe, A., Guasti, T., Nespor, M., Dupoux, E. & Van Ooyen, B. (1997). Reflections on phonological bootstrapping: Its role for lexical and syntactic acquisition. Language and Cognitive Processes, 12, 585-612. [ Links ]
Doupe, A. J. & Kuhl, P. K. (1999). Birdsong and human speech: Common themes and mechanisms. Annual Review of Neuroscience, 22, 567-631. [ Links ]
Endress, A. D., Cahill, D., Block, S., Watumull, J. & Hauser, M. D. (2009). Evidence of an evolutionary precursor to human language affixation in a non-human primate. Biology Letters, 5, 749-751. [ Links ]
Endress, A. D., Carden, S., Versace, E. & Hauser, M. D. (2010). The apes' edge: positional learning in chimpanzees and humans. Animal Cognition, 13, 483-495. [ Links ]
Gentner, T. Q., Fenn, K. M., Margoliash, D. & Nusbaum, H. C. (2006). Recursive syntactic pattern learning by song birds. Nature, 440, 1204-1207. [ Links ]
Gleitman, L. & Wanner, E. (1982). The state of the state of the art. In L. Gleitman & E. Wanner (Eds.), Language acquisition: The state of the art. Cambridge: Cambridge University Press. [ Links ]
Hauser, M. D., Barner, D. & O'Donnell, T. (2007). Evolutionary Linguistics: A New Look at an Old Landscape. Language Learning and Development, 3, 101-132. [ Links ]
Hauser, M. D., Chomsky, N. & Fitch, W. T. (2002). The faculty of language: What is it, who has it, and how did it evolve? Science, 298, 1569-1579. [ Links ]
Hauser, M. D. & Glynn, D. (2009). Can free-ranging rhesus monkeys (Macaca mulatta) extract artificially created rules comprised of natural vocalizations? Journal of Comparative Psychology, 123, 161-167. [ Links ]
Herbranson, W. & Shimp, C. (2008). Artificial grammar learning in pigeons. Learning & Behavior, 36, 116-137. [ Links ]
Janik, V. M., Sayigh, L. S. & Wells, R. S. (2006). Signature whistle shape conveys identity information to bottlenose dolphins. Proceedings of the National Academy of Sciences, 103, 8293-8297. [ Links ]
Marcus, G. F., Vijayan, S., Bandi Rao, S. & Vishton, P. M. (1999). Rule learning by seven-month-old infants. Science, 283, 77-80. [ Links ]
Murphy, R., Mondragón, E. & Murphy, V. (2008) Rule learning by rats. Science, 319, 1849-1851. [ Links ]
Nespor, M., Shukla, M., Vijner, R., Avesani, C., Schraudolf, H. & Donati, C. (2008). Different phrasal prominence realizations in VO and OV languages. Lingue e Linguaggio, 2, 139-168. [ Links ]
Newport, E. L., Hauser, M. D., Spaepen, G. & Aslin, R. N. (2004). Learning at a distance II. Statistical learning of non-adjacent dependencies in a non-human primate. Cognitive Psychology, 49, 85-117. [ Links ]
Pons, F. & Toro, J. M. (2010). Structural generalizations over consonants and vowels in 11-month-old infants. Cognition, 116, 361-367. [ Links ]
Ramus, F., Hauser, M. D., Miller, C., Morris, D. & Mehler, J. (2000). Language discrimination by human newborns and by cotton-top tamarin monkeys. Science, 288, 349-351. [ Links ]
Ramus, F., Nespor, M. & Mehler, J. (1999). Correlates of linguistic rhythm in the speech signal. Cognition, 73, 265-292. [ Links ]
Saffran, J. R., Aslin, R. N. & Newport, E. L. (1996). Statistical learning by 8-month-old infants. Science, 274, 1926-1928. [ Links ]
Toro, J. M., Nespor, M., Mehler, J. & Bonatti, L. L. (2008). Finding words and rules in a speech stream: Functional differences between vowels and consonants. Psychological Science, 19, 137-144. [ Links ]
Toro, J. M. & Trobalon, J. B. (2005). Statistical computations over a speech stream in a rodent. Perception y Psychophysics, 67, 867-875. [ Links ]
Toro, J. M., Trobalon, J. B. & Sebastian-Galles, N. (2005). Effects of backward speech and speaker variability in language discrimination by rats. Journal of Experimental Psychology: Animal Behavior Processes, 31, 95-100. [ Links ]
Van Heijningen, C. A. A., De Visser, J., Zuidema, W. & Ten Cate, C. (2009). Simple rules can explain discrimination of putative recursive syntactic structures by a songbird species. Proceedings of the National Academy of Sciences, 106, 20538-20543. [ Links ]
Zuberbuhler, K. (2006). Language evolution: The origin of meaning in primates. Current Biology, 16, R123-125. [ Links ]

