










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSuma Psicológica
versión impresa ISSN 0121-4381
Suma Psicol. v.18 n.1 Bogotá ene./jun. 2011
Estimación temporal en seres humanos mediante el procedimiento pico con interrupciones
Human Timing in the peak procedure with GAPS
1Facultad de Psicología, Universidad Nacional Autónoma de México, México.
Esta investigación fue financiada por el proyecto 82990-H de CONACYT e IN 305109 de PAPIIT. Agradecemos los valiosos comentarios y críticas hechas a este trabajo por parte del doctor Florente López y el doctor Arturo Bouzas.
La correspondencia relacionada con este artículo debe dirigirse a Ramsés Vázquez-Lira. Correo electrónico: chchemo@gmail.com
Recibido: Noviembre 19 2010 Aceptado: Marzo 12 2011
Resumen
Diversos procedimientos empleados en el estudio del aprendizaje animal han sido adaptados para investigación con humanos; específicamente, los procedimientos de intervalo fijo, bisección y generalización temporal, y el procedimiento pico, originalmente empleados para estudiar el proceso de estimación temporal en animales no humanos, han sido también utilizados con humanos, brindando importantes conocimientos para la comprensión del comportamiento humano. El presente trabajo reporta la adaptación para humanos del procedimiento pico con interrupciones, utilizado en las últimas décadas para dilucidar aspectos atencionales y mnémicos del proceso de estimación temporal. Se encontró que los 35 participantes (22.3 ± 3.1 años de edad) tienden a detener su estimación temporal durante la interrupción, conservando en memoria temporal el tiempo previo a la interrupción; los presentes resultados sugieren que la tasa de decaimiento de memoria temporal en humanos es menor que en cualquiera de las otras especies que han sido evaluadas en este procedimiento.
Palabras clave: estimación temporal; interrupción; procedimiento pico; teoría de expectancia escalar; humanos.
Abstract
Several procedures used in the study of animal learning have been adapted for human research; specifically, the fixed interval schedule, temporal bisection, temporal generalization, and the peak procedure, originally used in research on animal timing, have also been used in humans, providing important insights for understanding human behavior. This paper reports the adaptation to humans of the peak procedure with gaps, which was used in the past decades to elucidate attentional and mnemonic aspects of the timing process. It was found that the 35 participants (22.3 ± 3.1 years old) tend to stop their timing during the gap, keeping a representation of the pre-gap duration in working memory; these results suggest that the rate of decay of temporal memory in humans is lower than in any other species that have been evaluated in this procedure.
Keywords: Gap; Scalar Expectancy theory; peak procedure; timing; humans.
La estimación temporal es fundamental para muchos procesos motores y cognitivos. En el nivel motor, los individuos coordinan temporalmente sus movimientos, para llevar a cabo acciones, por ejemplo patear una pelota o detener su caminar ante un semáforo con luz roja. Del lado cognitivo, la estimación temporal es crítica para el aprendizaje asociativo y para la representación de relaciones secuenciales entre estímulos en el ambiente, entre muchas otras actividades. Debido a que no existen receptores sensoriales explícitos para el tiempo, la información temporal posiblemente se derive a través de la operación de algunos mecanismos neuronales, cuyas propiedades están sujetas a gran controversia.
Para explicar el comportamiento de animales y humanos en procedimientos de estimación temporal, se han propuesto diversas teorías que tratan de dar cuenta de dicho fenómeno. Una de las principales es la teoría de expectancia escalar (SET por sus siglas en inglés; para una revisión véase Gibbon, 1977), la cual propone un constructo que ha dominado la literatura de estimación temporal en animales y seres humanos: un dispositivo generador-acumulador de pulsos, el cual sirve como reloj ubicuo, y en conjunto con otros procesos permite al individuo discriminar entre diferentes duraciones temporales. Aunque esta teoría emerge de la literatura animal como una explicación de las regularidades encontradas en múltiples investigaciones (Church, 1984; Gibbon, 1977, 1991), también es capaz de modelar el comportamiento de seres humanos en tareas de percepción y estimación temporal (Allan, 1998).
Existen diferentes procedimientos que han sido utilizados para estudiar el proceso de estimación temporal, entre ellos se incluyen el programa intervalo fijo (Ferster & Skinner, 1957), el procedimiento pico (Catania, 1970), y el procedimiento de pico con interrupciones (S. Roberts & Church, 1978), la bisección temporal (Church & Deluty, 1977), y el reforzamiento diferencial de tasas bajas (Ferster & Skinner, 1957), entre otros. De acuerdo con la SET, el desempeño en el programa intervalo fijo es un indicador del proceso de estimación temporal. Sin embargo, debido a que dicho procedimiento solo nos permite cuantificar la conducta del sujeto hasta el final del intervalo a estimar, ya que una vez que se ha obtenido el reforzador no se genera información adicional sobre la conducta de estimación temporal, Catania (1970) desarrolló el procedimiento de pico, el cual ha sido extensamente empleado (Balsam, Drew & Gallistel, 2010; Cabeza de Vaca, Brown & Hemmes, 1994; Morrissey, Wogar, Bradshaw & Szabadi, 2000; S. Roberts, 1981). Este procedimiento consiste en la combinación de dos tipos de ensayos, aquellos que son idénticos a los presentados en los programas intervalo fijo (ensayos intervalo fijo) y otros sin reforzamiento (ensayos pico) que además se distinguen por una mayor duración, que suele ser por lo menos tres veces mayor al valor del programa intervalo fijo en curso. El resultado principal en este procedimiento es que la distribución temporal de respuestas en los ensayos pico tiene la forma de una curva normal, con un máximo cercano al valor del intervalo fijo de los ensayos reforzados (tiempo pico).
A pesar de que el procedimiento pico ha sido una herramienta importante en la evaluación de la habilidad de palomas (Cabeza de Vaca, et al., 1994), ratas (S. Roberts, 1981), peces (Talton, Higa & Staddon, 1999), abejas (Boisvert & Sherry, 2006) y seres humanos (Rakitin, et al., 1998) para estimar intervalos de tiempo, existe una pregunta de investigación de gran valor teórico, que no puede ser respondida por el procedimiento pico: El efecto de una interrupción sobre el estímulo a estimar. Ante este escenario surge el procedimiento pico con interrupciones (S. Roberts, 1981; S. Roberts & Church, 1978; W. A. Roberts, Cheng & Cohen, 1989) el cual agrega al procedimiento pico un tipo de ensayo adicional, llamado ensayo con interrupción. Estos ensayos son similares a los ensayos pico, pero el estímulo a estimar es apagado por un breve periodo de tiempo y reencendido de nuevo. El objetivo principal del procedimiento pico con interrupciones es cuantificar si existe un cambio en el tiempo pico debido a la interrupción del estímulo a estimar (Buhusi & Meck, 2000; S. Roberts, 1981; S. Roberts & Church, 1978). Existen tres posibles resultados: (a) se retiene la duración del intervalo previa a la interrupción, y por lo tanto al final de la interrupción se continúa la estimación del intervalo hasta completar el tiempo criterio; (b), la estimación del intervalo comienza de nuevo al final de la interrupción. Estos posibles resultados presumiblemente implican diferentes modos de operación del reloj interno (S. Roberts, 1981), detención y reinicio, respectivamente. Finalmente, es también posible que (c), el sujeto continúe su estimación temporal durante la interrupción, por lo que no se genera un desfase en el tiempo pico. Se ha realizado una cantidad considerable de estudios para entender los determinantes del empleo de estas estrategias, sin que hasta la fecha exista un acuerdo unánime acerca de ellos, (para una revisión véase, Buhusi, Perera & Meck, 2005; W. A. Roberts, et al., 1989).
Las dos principales explicaciones teóricas sobre el comportamiento en este procedimiento son las hipótesis de ambigüedad instruccional y la de decaimiento de memoria. La primera de ellas asume que en el transcurso del entrenamiento en un procedimiento pico típico, los animales han sido entrenados a reiniciar el reloj al final de los ensayos, el cual es indicado por la entrega del reforzador y/o la presentación del intervalo entre ensayos (IEE). Debido a este entrenamiento, cuando las interrupciones en el estímulo cuya duración se estima son iguales al IEE, los animales harán lo que han sido entrenados a hacer, es decir, reiniciar su estimación temporal; esta hipótesis implica que si las interrupciones son notablemente diferentes al IEE los animales no reiniciarán. Por lo tanto, esta hipótesis predice que las manipulaciones que hacen ver a la interrupción semejante al IEE, generan en los sujetos un reinicio de su reloj. En contraste, manipulaciones en la interrupción que la hacen distinta al IEE, dan como resultado que los sujetos detengan su estimación temporal y sea reanudada tras la interrupción (Kaiser, Zentall & Neiman, 2002).
Por otro lado, la hipótesis de decaimiento de memoria fue sugerida por Cabeza de Vaca et al. (1994), con base en el hallazgo de que entre más larga sea la interrupción, mayor es el desplazamiento en el tiempo pico. Esto es compatible con la idea de que el tiempo subjetivo almacenado en la memoria de trabajo decae pasivamente durante la interrupción, dando lugar a un decaimiento mínimo para interrupciones cortas, y a un decaimiento mayor cuando la interrupción incrementa su duración. Es decir, a medida que la duración de la interrupción aumenta, el modo de operación aparentemente cambia de detención a reinicio, por lo que este modelo nos explica los aparentes cambios en el modo de operación del reloj interno ante diferentes duraciones en la interrupción.
La revisión presentada deja claro que el modo de operación del reloj interno está en función de una diversidad de variables; sin embargo, ante la nula investigación en seres humanos con este procedimiento, parece importante evaluar los efectos de la presencia, duración (corta/ larga) y localización (temprana/tardía) de una interrupción en el estímulo a medir.
El procedimiento empleado en este trabajo combina al procedimiento pico con interrupciones utilizado en animales (Cabeza de Vaca, et al., 1994; S. Roberts, 1981) con la versión computarizada del procedimiento de pico utilizada en seres humanos (Hinton & Rao, 2004; Rakitin, et al., 1998).
Método
Participantes
35 estudiantes (11 hombres y 24 mujeres) de licenciatura de semestres iniciales (segundo a cuarto semestre) de diversas carreras de la Universidad Nacional Autónoma de México, con edades entre 18 y 25 años (22.3 ± 3.1 años). Todos los participantes tenían vista normal o corregida. Dichos participantes asistieron voluntariamente por puntos extra en sus materias.
Aparatos
10 computadoras con procesador Pentium 4® y Sistema operativo Windows XP®, con resolución en monitor de 1280*1024 pixeles, ratón y teclado.
Procedimiento
Cada participante se asignó a la misma computadora, y realizó la tarea a la misma hora del día en cada una de las cuatro sesiones experimentales. Las sesiones se realizaron entre las 12 y las 15 horas, con una duración aproximada de 45 minutos. En el monitor aparecieron las instrucciones para la tarea experimental (véase el Apéndice). Una vez que los participantes completaron la lectura de las instrucciones en la pantalla, en caso de tener alguna duda recibieron instrucciones verbales adicionales, las cuales tenían como fin dejar claro el objetivo de la tarea; al expresar la comprensión del objetivo de la tarea, iniciaron la sesión.
Entrenamiento
Consistió en 20 ensayos en los cuales en el centro del monitor aparecía un cuadro azul (100*100 pixeles) en un fondo gris claro; dentro de dicho cuadro azul aparecían dígitos con una duración de 0.5 segundos y con un intervalo entre estímulos variable con una media de 1.5 segundos. Los dígitos distractores se emplearon con el fin de que los participantes no contaran. El cuadro azul en el centro de la pantalla indicaba el tiempo a estimar (10 segundos) y su cambio a color rojo al final del intervalo, indicaba al participante el final del mismo, este nuevo color permanecía durante 2 segundos. Posteriormente, se presentaron 10 ensayos pico en los cuales debían emitir por lo menos 5 respuestas en el momento en que juzgaran que el cambio del color ocurriría.
Sesión 1. Después de terminar el entrenamiento daba inicio esta sesión. La tarea experimental consistió en 80 ensayos totales. Los 10 primeros ensayos fueron ensayos iguales a los presentados en el entrenamiento. Los 70 ensayos restantes estaban distribuidos semi-aleatoriamente, los cuales tenían la restricción de aparecer por lo menos al quinto ensayo pico. Los 10 ensayos eran iguales a los presentados en el entrenamiento y los 60 ensayos restantes fueron ensayos pico, que iniciaban a partir de que el cuadro aparecía en la pantalla. La particularidad de estos ensayos radicaba en la supresión del cambio a color rojo transcurrido el intervalo a estimar. En estos ensayos el participante debía emitir sus respuestas lo más cercanas a la duración del intervalo a estimar. Las respuestas se emitían con la tecla Espacio, mientras que para finalizar cada ensayo el sujeto debía oprimir la tecla Enter, lo que daba inicio a un IEE variable de 10 segundos en promedio. En el momento que el IEE terminaba, se habilitaba en la pantalla la posibilidad de oprimir la tecla Enter para comenzar un nuevo ensayo. En caso de no registrarse por lo menos 5 respuestas en un periodo de 40 segundos el ensayo se repetía.
Sesiones 2-4. Estas sesiones constaron de 80 ensayos. Los 10 primeros fueron iguales a los presentados en el entrenamiento, de los 60 ensayos pico restantes, 30 ensayos pico fueron sustituidos por ensayos pico con interrupciones. En este tipo de ensayos el estímulo a estimar desapareció por un breve periodo de tiempo (duración de la interrupción), para después volver a aparecer. De estos 30 ensayos, en 10 ensayos la interrupción fue temprana (duró 2 segundos y comenzó a los 2 segundos de iniciado el ensayo), en 10 ensayos la interrupción fue tardía (duró 2 segundos, comenzando a los 5 segundos) y en 10 ensayos la interrupción fue larga (duró 5 segundos y se inició a los 2 segundos transcurrido el ensayo). En cada sesión se presentaron de manera semi-aleatoria 10 ensayos de recordatorio (tiempo fijo), iguales a los del entrenamiento e indicados desde el inicio del ensayo.
Análisis de datos
Los datos se ordenaron en bines en donde se examinó la distribución de respuestas a través de un periodo de tiempo. Para todos los tipos de ensayo, se generaron 120 bines de 0.25 segundos equivalentes a 30 segundos por ensayo. Se registró en qué periodo ocurrió cada una de las respuestas para generar la distribución de frecuencia de respuestas por bin en cada sesión. Posteriormente se transformó la frecuencia a tasa de respuestas por minuto, para que finalmente a cada distribución de cada participante se le ajustara la siguiente ecuación:
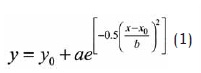
En la que α representa la tasa pico, b la desviación estándar y x0 el tiempo pico; finalmente la fracción de Weber se calculó dividiendo la desviación estándar entre el tiempo pico. El valor de estos parámetros fue analizado por medio de un ANOVA de medidas repetidas, con Tipo de Ensayo (pico, interrupción temprana, tardía y larga) y Sesión (2-4) como factores intrasujeto y Género como factor entre sujetos. La significancia de todos los análisis fue α < 0,05.
Resultados
La estimación temporal se evaluó al analizar los parámetros obtenidos del ajuste de la ecuación 1 a los datos de cada participante, en cada tipo de ensayo en cada sesión. En la Tabla 1 se muestra el valor promedio y el error estándar de cada parámetro analizado.

El promedio de las funciones de pico de todos los participantes en cada una de las condiciones evaluadas y su correspondiente ajuste gaussia-no se muestran en la Figura 1. Las funciones muestran un patrón de incremento súbito en la densidad de respuesta cercana a la duración del intervalo a estimar (10 segundos), seguido por un decremento simétrico hasta una tasa de respuesta cercana a cero. De manera general se observa una distribución de respuestas normal (con un ajuste promedio R2> 0.95) con una baja dispersión.
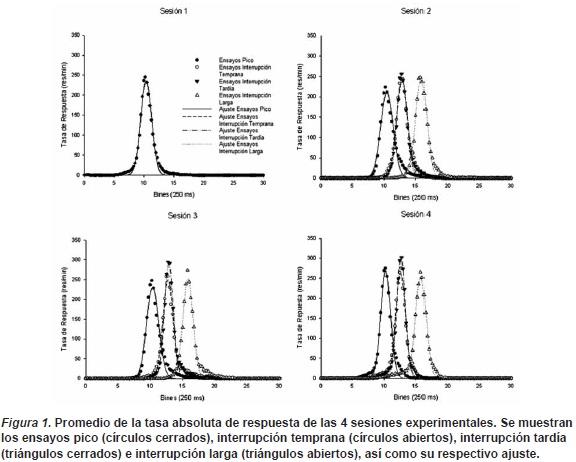
El primer ANOVA realizado demostró que el factor género no provocó diferencias significativas en ningún parámetro (F(1,27)=3.41, p=0,07 F(1,27)=0.63, p=0,48 y F(1,27)=0.10, p=0,40 para tiempo pico, desviación estándar y tasa pico, respectivamente), por lo que para subsecuentes análisis ambos géneros se agruparon.
Al analizar las diferencias en el tiempo pico con el factor tipo de ensayo, se obtuvo una F(3,102)=1243.60, p<0,001, η2=0.97. En los análisis Post-Hoc se encontró un incremento significativo en los ensayos con interrupciones tempranas, tardías y largas (p<0,001), respecto a los ensayos pico sin interrupción. No hubo diferencia en el tiempo pico entre los ensayos en donde las interrupciones fueron tempranas y tardías. Al evaluar el tiempo pico con el factor sesión, se obtuvo una F(2,68)=10.49, p<0,001, η2=0.24, encontrando una disminución significativa entre la sesión 2 y la sesión 4 (p<0,001).
Se analizaron las diferencias en la desviación estándar en los distintos tipos de ensayos. Se obtuvo una F(3,102)=7.93, p<0,001, η2=0.19, encontrando por medio del análisis Post-Hoc decrementos significativos entre los ensayos pico y los ensayos con interrupción tardía y larga (p<0,001). Analizando el factor sesión, se obtuvo una F(2,68)=3.93, p=0,02, η2=0.10; se encontró una disminución significativa entre la sesión 2 y 4 (p=0,01).
En relación con la evaluación de la tasa pico con el factor tipo de ensayo se obtuvo una F(3,102)=12.78, p<0,001, η2=0.27. En el análisis Post-Hoc se encontraron incrementos significativos entre los ensayos pico y cada uno de los ensayos con interrupciones (p<0,001). Para el análisis del factor sesión se asocia una F(2, 68)=4.71, p=0,01, η2=0.12, encontrando de igual manera un incremento significativo entre la Sesión 2 y la Sesión 4 (p=0,01). Dado que la tasa de respuestas por minuto tuvo cambios significativos a lo largo de las sesiones y en conjunción con el decremento significativo del tiempo pico, se observa que los participantes emitieron más respuestas en el momento adecuado, conforme las sesiones avanzaron.
La Figura 2 presenta el diagrama de caja del tiempo pico para cada tipo de ensayo; se observan diferencias entre los ensayos pico y los ensayos pico con interrupciones, por lo que se evaluó si esta diferencia en el tiempo pico ajustaba a alguna de las dos principales hipótesis de operación del reloj interno: Detención o reinicio.
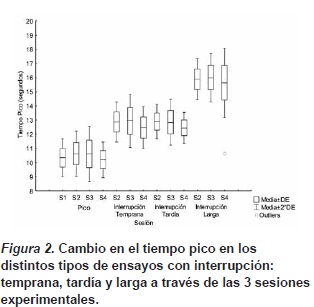
El valor obtenido en cada tipo de interrupción en cada sesión fue contrastado con el valor del tiempo pico de los ensayos pico sin interrupción más el valor del desfase esperado por la hipótesis de detención (2 segundos para las interrupciones tempranas y tardías, y 5 segundos para las interrupciones largas). Al evaluar las diferencias entre el desfase encontrado en cada uno de los tipos de interrupción y el predicho por la hipótesis de detención se encontraron diferencias significativas, lo que demuestra que el desfase encontrado fue mayor al predicho por la hipótesis de detención; mediante un ANOVA de medidas repetidas se obtuvo una F(1, 34)=9.16, p<0,05, η2=0.21 para los ensayos interrupción temprana, un valor F(1, 34)=9.33, p<0,05, η2=0.22 para los ensayos interrupción tardía y finalmente para los ensayos interrupción larga una F(1, 34)=5.32, p=0,02, η2=0.14.
Al realizar este mismo análisis con la hipótesis de reinicio se esperaba que el desfase en los ensayos con interrupción temprana fuera de 4 segundos y en los ensayos con interrupción tardía y larga fuera de 7 segundos, debido a que dicho desfase sería equivalente a la duración del ensayo antes de la interrupción más la duración de la interrupción misma. De manera semejante se halló que el desfase en el tiempo pico no ajustó a esta hipótesis en ningún tipo de ensayo. Los ANOVA realizados muestran diferencias significativas entre el cambio en el tiempo pico predicho por la hipótesis de reinicio y el cambio en el tiempo pico encontrado en la condición interrupción temprana F(1, 34)=362.20, p<0,001, η2=0.91, interrupción tardía F(1, 34)=3287.20, p<0,001, η2=0.99, e interrupción larga F(1, 34)=68.84, p<0,001, η2=0.67.
Debido a restricciones en la cantidad de equipo de cómputo disponible, el horario de aplicación de las sesiones experimentales no fue uniforme para todos los participantes, por lo que existe la posibilidad de que existieran diferencias en la ejecución debido al impacto de ritmos circadianos (Adan, 1993). A pesar de que las sesiones experimentales se realizaron en un pequeño rango (12 a 15 horas), realizamos análisis estadísticos adicionales para evaluar el impacto del horario, encontrando que el horario no ejerció un efecto significativo en ninguna de las variables relacionadas con estimación temporal (todas las razones F < 1.32 todas las p > 0, 28).
Discusión
La presente investigación tuvo como principal objetivo estudiar en seres humanos bajo la perspectiva de SET (Gibbon, 1977) una de las propiedades del reloj interno, el modo de operación. Por esta razón, utilizamos el procedimiento pico con interrupciones. Las funciones de distribución de respuesta durante los ensayos pico (incluyendo aquéllos con interrupciones) para los datos individuales de este experimento fueron descritas apropiadamente por una ecuación Gaussiana de 3 parámetros, que ha sido ampliamente utilizada en la literatura animal (S. Roberts, 1981).
El primer hallazgo significativo de la tarea experimental fue que las funciones de respuesta en los ensayos pico resultaron semejantes a las reportadas por Rakitin, et al. (1998). Una vez demostrada la validez del procedimiento pico, se procedió a analizar el impacto de las interrupciones en la estimación temporal. Lo más importante de los datos en los ensayos pico con interrupciones es el análisis del ajuste hacia alguna de las dos principales hipótesis de operación del reloj interno. Las comparaciones de los tiempos pico obtenidos en los ensayos con los 3 tipos de interrupciones y el tiempo pico obtenido en los ensayos pico (cambio en el tiempo pico), nos sugiere un mejor ajuste de los datos para la hipótesis de detención, lo cual se observa en el desfase de los tiempos pico de la Figura 2. A pesar de que en cada uno de los tres tipos de ensayos con interrupción (temprana, tardía y larga), los análisis estadísticos nos indican que existen diferencias significativas entre el tiempo pico de los ensayos con interrupciones y el tiempo pico esperado de acuerdo a la hipótesis de detención, el desfase es sumamente cercano al predicho por la hipótesis de detención, sugiriendo que la estimación del intervalo fue detenida durante la interrupción, conservando el tiempo previo a la interrupción en memoria.
Los resultados del presente experimento también permiten comparar el desempeño entre especies en una tarea de estimación temporal inmediata. Existe una amplia línea de investigación en múltiples especies animales, que abarca peces (Talton, et al., 1999), ratas (S. Roberts, 1981), palomas (Cabeza de Vaca, et al., 1994) y evidentemente seres humanos (Rakitin, et al., 1998), encontrando que todas las especies estudiadas exhiben en general la misma característica: estimación temporal de un intervalo de tiempo muy cercana a la duración del intervalo presentado. Teniendo como referencia estos antecedentes en múltiples especies animales, el comparar los modos de operación del reloj interno entre especies por medio del procedimiento pico con interrupciones puede generar especial interés en el dominio de la cognición animal, ya que los datos obtenidos en seres humanos se aproximan cercanamente hacia la hipótesis de detención. En términos de diferencias entre especies, se ha encontrando que las palomas tienden a reiniciar (W. A. Roberts, et al., 1989), mientras que las ratas pueden detener o reiniciar, siendo una especie flexible en este ámbito, dependiendo de la manipulación experimental (Buhusi, et al., 2005; S. Roberts, 1981). Este estudio complementa a la investigación animal con el reporte que arroja claros indicios de que el modo de operación del reloj interno en seres humanos se aproxima hacia la estrategia de detención.
De esta manera esta investigación representa la primera aproximación al estudio de una de las propiedades del reloj interno que ha sido normalmente abordada desde el modelo SET. Por otra parte, bajo la perspectiva de la hipótesis de decaimiento de memoria (Cabeza de Vaca, et al., 1994) los presentes resultados sugieren que la tasa de decaimiento de memoria temporal en seres humanos es menor que en cualquiera de las otras especies que han sido evaluadas en este procedimiento. Los resultados presentados abren un amplio panorama para continuar estudiando variables relacionadas con el modo de operación del reloj interno.
Referencias
Adan, A. (1993). Circadian variations in psychological measures: A new classification. Choronobiologia, 20(3-4), 145-162. [ Links ]
Allan, L. G. (1998). The influence of the scalar timing model on human timing research. Behavioural Processes, 44, 101-117. [ Links ]
Balsam, P. D., Drew, M. R. & Gallistel, C. R. (2010). Time and associative learning. Comparative Cognition & Behavior Reviews, 5, 1-22. [ Links ]
Boisvert, M. J. & Sherry, D. F. (2006). Interval timing by an invertebrate, the bumble bee Bombus impatiens. Current Biology, 16(16), 1636-1640. [ Links ]
Buhusi, C. V. & Meck, W. H. (2000). Timing for the absence of a stimulus: The gap paradigm reversed. Journal of Experimental Psychology: Animal Behavior Processes, 26(3), 305-322. [ Links ]
Buhusi, C. V., Perera, D. & Meck, W. H. (2005). Memory for timing visual and auditory signals in albino and pigmented rats. Journal of Experimental Psychology: Animal Behavior Processes, 31(1), 18-30. [ Links ]
Cabeza de Vaca, S., Brown, B. L. & Hemmes, N. S. (1994). Internal clock and memory processes in animal timing. Journal of Experimental Psychology: Animal Behavior Processes, 20(2), 184-198. [ Links ]
Catania, A. C. (1970). Reinforcement schedules and psychophysical judgments: A study of some temporal properties of behavior In W. N. Schoenfeld (Ed.), The theory of reinforcement schedules (pp. 1-42). New York: Appleton-Century-Crofts. [ Links ]
Church, R. M. (1984). Properties of the internal clock. Annals of the New York Academy of Sciences, 423, 566-582. [ Links ]
Church, R. M. & Deluty, M. Z. (1977). Bisection of temporal intervals. Journal of Experimental Psychology: Animal Behavior Processes, 3, 216-228. [ Links ]
Ferster, C. B. & Skinner, B. F. (1957). Fixed ratio Schedules of Reinforcement (pp. 39-132). New York: Appleton Century Crofts. [ Links ]
Gibbon, J. (1977). Scalar expectancy theory and Weber's Law in animal timing. Psychological Review, 84, 279-325. [ Links ]
Gibbon, J. (1991). Origins of scalar timing. Learning and Motivation, 22, 3-38. [ Links ]
Hinton, S. C. & Rao, S. M. (2004). "One-thousand one... one-thousand two...": Chronometric counting violates the scalar property in interval timing. Psychonomic Bulletin Review, 11(1), 24-30. [ Links ]
Kaiser, D. H., Zentall, T. R. & Neiman, E. (2002). Timing in pigeons: Effects of the similarity between intertrial interval and gap in a timing signal. Journal of Experimental Psychology: Animal Behavior Processes, 28, 416-422. [ Links ]
Morrissey, G., Wogar, M. A., Bradshaw, C. M. & Szabadi, E. (2000). Effect of lesions of the ascending 5-hydroxytryptaminergic pathways on timing behaviour and the circadian time-keeping system revisited. Brain Research Reviews, 33, 34-77. [ Links ]
Rakitin, B. C., Gibbon, J., Penney, T. B., Malapani, C., Hinton, S. C. & Meck, W. H. (1998). Scalar expectancy theory and peak-interval timing in humans. Journal of Experimental Psychology: Animal Behavior Processes, 24(1), 15-33. [ Links ]
Roberts, S. (1981). Isolation of an internal clock. Journal of Experimental Psychology: Animal Behavior Processes, 7, 242-268. [ Links ]
Roberts, S. & Church, R. M. (1978). Control of an internal clock. Journal of Experimental Psychology: Animal Behavior Processes, 4, 318-337. [ Links ]
Roberts, W. A., Cheng, K. & Cohen, J. S. (1989). Timing light and tone signals in pigeons. Journal of Experimental Psychology: Animal Behavior Processes 15, 23-35. [ Links ]
Talton, L. E., Higa, J. J. & Staddon, J. E. R. (1999). Interval schedule performance in the goldfish Carassius auratus. Behavioural Processes, 45, 193-206. [ Links ]


