










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSuma Psicológica
versión impresa ISSN 0121-4381
Suma Psicol. v.18 n.2 Bogotá jul./dic. 2011
ADQUISICIÓN DE LA RESPUESTA DE CONGELAMIENTO DE RATAS: DIFERENCIAS SEXUALES EN ADOLESCENTES Y ADULTOS
ACQUISITION OF FREEZING RESPONSE IN RATS: SEX DIFFERENCES IN ADOLESCENTS AND ADULTS
1Laboratorio de Análisis Experimental del Comportamiento Konrad Lorenz Fundación Universitaria, Colombia
2Centro de Investigaciones en Biomodelos
Proyecto realizado por el convenio Joven Investigador Becario Colciencias-Fundación Universitaria Konrad Lorenz y fue realizado con el financiamiento parcial de la Dirección de Investigaciones de la Konrad Lorenz al segundo autor. El proyecto fue presentado en el XXXIII Congreso Interamericano de Psicología. La correspondencia relacionada con este artículo debe ser dirigida a Lady Andrea Polanco. Correo electrónico: ladya.polancos@konradlorenz.edu.co
Recibido: Septiembre 9 2011 Aceptado: Noviembre 16 2011
Resumen
El Condicionamiento Pavloviano de Miedo es uno de los modelos preclínicos más comunes en el estudio del Trastorno de Estrés Post-traumático. El objetivo de la presente investigación consistió en utilizarlo como biomodelo para el estudio de las diferencias sexuales que caracterizan este síndrome, así como elaborar una descripción preliminar de las diferencias a lo largo de la trayectoria ontogenética. Se utilizaron 45 sujetos, de tres camadas diferentes de ratas ingenuas de ascendencia Wistar, 18 machos y 27 hembras, dos aproximadamente por camada para cada uno de los dos grupos experimentales: animales adolescentes y adultos. Los resultados señalan diferencias significativas en la segunda medición del estímulo condicionado en la interacción entre sexo y edad y al comparar las tres mediciones del estímulo condicionado. Se discuten los resultados en torno a las discrepancias en la literatura respecto al efecto de las variables evaluadas en la adquisición de miedo condicionado.
Palabras clave: diferencias sexuales, edad, miedo condicionado, congelamiento.
Abstract
Pavlovian fear conditioning is one of the most popular preclinical models in the study of Post-Traumatic Stress Disorder (PTSD). The aim of the present research was explore the sex differences that characterize PTSD by means of this experimental paradigm, as well as to offer a preliminary description of how these sex differences behave throughout development. Forty five native rats, of Wistar descent were used as subjects, with 18 males and 27 females approximately balanced by litter across the two experimental groups: adolescents and adults. The results show significant differences in the second measurement of the conditioned stimulus in the interaction between sex and age and to compare the tree measurements of the conditioned stimulus. Results are discussed regarding the discrepancies in the literature regarding the effect of the variables evaluated in the acquisition of Conditioned fear.
Keywords: sex differences, age, conditioned fear, freezing.
El Manual de Diagnóstico y Estadístico de los Trastornos Mentales (American Psychiatrich Association, 2000), define al Trastorno por Estrés Post Traumático (TEPT) como un trastorno de ansiedad que se caracteriza por una re-experimentación de acontecimientos altamente traumáticos, presentando síntomas como, el aumento de la activación y comportamiento de evitación a estímulos relacionados con el trauma. De acuerdo con el Estudio Europeo de Epidemiología de los Trastornos Mentales, el 13,6% de la población de seis países europeos (Bélgica, Francia, Alemania, Italia, Holanda y España) han padecido algún trastorno de ansiedad a lo largo de su vida y el 6,4% ha experimentado este cuadro en el último año (Alonso et al., 2004). Por otra parte, la prevalencia del TEPT fue estimada entre el 1 y el 4% de la población mundial, según la clasificación internacional de enfermedades por la Organización Mundial de la Salud (OMS) (2001).
Para Colombia, los datos del 2003 de la encuesta de la OMS, señalan que los trastornos de ansiedad son el tipo de trastorno de salud Mental de mayor prevalencia a nivel nacional, afectando al 19.3% de la población. De acuerdo con estos datos, Bogotá exhibe la prevalencia vitalicia más elevada de trastornos de ansiedad (46,7%) (Posada-Villa, Buitrago-Bonilla, Medina-Barreto & Rodríguez-Ospina, 2006). En lo que respecta al TEPT, el índice de prevalencia de vida es 1.8% pero esta cifra puede aumentar a 37% en poblaciones de alta exposición a la violencia, de acuerdo con los estudios de Pineda, Guerrero, Pinilla, & Estupiñán (2002) y el de Londoño et al., (2005), en donde las victimas de la guerra sufren trastornos emocionales que reflejan la dificultad del ser humano para adaptarse a la violencia, y en un país como Colombia, en donde las personas en las últimas dos décadas, han tenido como mínimo una vez en su vida un acontecimiento violento, el riego de desarrollar graves problemas de salud mental es mucho más alto.
Una de las variables más consistentemente asociadas a la ocurrencia de los trastornos de ansiedad es el sexo. En Colombia, tres de cada diez mujeres y dos de cada diez hombres han presentado trastornos de ansiedad alguna vez en la vida (Posada-Villa et al., 2006). Específicamente en el caso del TEPT, la prevalencia es mayor en mujeres (2.5%) que en hombres (0.8%) (Alonso et al., 2004; véase también Posada-Villa et al., 2006). No obstante, algunos estudios experimentales sobre la adquisición de la respuesta de miedo en humanos reportan que los hombres presentan significativamente mayor respuesta condicionada que las mujeres (Milad, et al., 2006), mientras que algunos estudios epidemiológicos reportan que no hay evidencia de una relación significativa entre el sexo ni la edad con los síntomas de TEPT (Echenique, Medina, medina & Ramírez, 2008; Kohn, Koenen & Jaffee, 2009). Uno de los análisis más comprehensivos hasta la fecha del papel de las diferencias sexuales en el TEPT, es el metanálisis de Tolin y Foa (2006), en el cual analizaron 290 artículos que investigaran la prevalencia y severidad de TEPT en mujeres y hombres reportados desde 1980 hasta julio de 2005, después de hacer una selección de artículos de acuerdo a los criterios de exclusión para el estudio. En el estudio utilizaron las siguientes categorías: agresiones sexual, abuso sexual infantil, accidente, agresiones no sexuales, desastres, abuso no sexual, combates, guerra o terrorismo, testigos de muestres o lesiones y enfermedad o lesión no especificada. En el análisis de los artículos, los autores señalan que los hombres están expuestos con mayor frecuencia a eventos amenazantes o traumáticos (accidentes, ataques no sexuales, combates o guerras, desastres entre otros), mientras quelas mujeres que están más expuestas a casos de abuso y ataque sexual. No obstante, se encuentran diferencias sexuales significativas en cada uno de los eventos traumáticos, indicando una mayor frecuencia y severidad de TEPT en las mujeres que en los hombres. Al controlar estadísticamente la incidencia de exposición a eventos traumáticos, tanto para hombres como para mujeres, la mayor ocurrencia de TEPT sigue siendo mayor en las mujeres. En vista de lo anterior, la alta prevalencia de TEPT en las mujeres no se debe únicamente a un mayor riesgo de vivir eventos traumáticos específicos.
Algunas de las variables que pueden influir en las diferencias sexuales en TEPT, son las respuestas neuroendocrinas y los procesos de afrontamiento de la situación traumática.
Las mujeres perciben el evento de forma más amenazante y con mayor pérdida de control que los hombres, contribuyendo a mayor TEPT en mujeres (Arenas & Puigcerver, 2009). Las bases biológicas del miedo condicionado han sido descritas con bastante detalle y constituyen una rica fuente de hipotesis para la explicación de las diferencias sexuales en la respuesta de ansiedad. Los humanos a nivel sexual presentan diferencias en la estructura y funcionalidad de la amigdala y la corteza prefrontal, al igual que en otras estructuras involucradas en la adquisición y extinción del condicionamiento pavloviano de miedo (Milad et al., 2006). Así mismo, la diferencia en la proporción de hormonas gonadales que distinguen a uno y otro sexo es también una fuente potencial de diferencias en la adquisición del miedo condicionado (Milad et al., 2010; Mitra y Sapolsky, 2010).
Por otra parte, desde los estudios de Pavlov (1927) de condicionamiento alimentario, en donde provocó la salivación de perros por la presentación de los estímulos que se habían asociado con la entrega de alimento, se han realizado estudios con animales no humanos para mostrar que frente a estímulos nocivos se presentan reflejos condicionados en respuesta a señales de advertencia que asemejan las condiciones originales de los humanos; a este mecanísmo, Pavlov lo denominó mecanismo de defensa y actualmente se conoce como condicionamiento pavloviano de miedo (Bouton & Bolles, 1980, Davis, 1992, Fanselow, 1980, Ledoux, 1995).
El Condicionamiento pavloviano de miedo es uno de los arreglos experimentales más utilizados como modelo del TEPT. Este arreglo consiste en administrar una leve estimulación eléctrica (Estímulo Incondicionado-EI), apareado a un Estímulo Condicionado (EC), que suele o bien ser el contexto de condicionamiento, un estímulo extereoceptivo como un tono o una luz (Domjan 2005; Fanselow & Tighe, 1988; Palanza, 2001). La presentación apareada del EC y EI, conlleva a que el EC elicite una respuesta condicionada (RC), como es el congelamiento, aún en ausencia del EI (Fanselow & Tighe, 1988, Palanza, 2001). La respuesta de congelamiento o inmovilización, se define como la ausencia de movimiento, excepto el necesario para respirar (Bolles, 1970; véase también Fanselow & Tighe, 1988). El congelamiento es una reacción adaptativa de los individuos a un estímulo ansiogénico y su estudio es común en laboratorios comportamentales (Rau & Fanselow, 2009). En muchas especies el congelamiento es la respuesta inicial a un peligro, ya que prepara al animal para luchar o huir en caso de ser atacado o reduce la probabilidad de ser detectado por un posible predador (Bolles, 1970). Otros comportamientos asociados a la respuesta de miedo incluyen cambio en tasa cardiaca, presión sanguínea, respiración, e incremento en la tasa de sobresalto y atención (Fanselow, DeCola & Young, 1993).
En cuanto a diferencias sexuales, en condicionamiento pavloviano de miedo en roedores, Pryce, Lehmann y Feldon (1999) evidenciaron como las hembras de las cepas Wistar y Fischer presentan una tendencia mayor de adquisición y extinción al condicionamiento contextual y al EC en relación a los machos, similares a los reportes de Kosten, Lee, y Kim, (2006) y Wiltgen, Sanders, Behne & Fanselow, (2001). No obstante, en otros estudios con ratones, jerbos y ratas (Ide et al., 2010; Starkey & Bridges, 2010; Verma, Hellemans, Choi, Yu & Weinberg, 2010 respectivamente) no se han encontrado diferencias sexuales claras. Aunque en otros estudios, se encuentra, igual que en la mayoría de reportes en humanos, que las hembras presentan niveles altos de congelamiento en comparación a los machos (Baran, Armstrong, Niren, Hanna, Conrad, 2009; Diehl et al., 2007).
Es posible que las discrepancias en los estudios de diferencias sexuales se deba a que los machos son más propensos a seleccionar y dar una respuesta inactiva, es decir de congelamiento y las hembras por el contrario una respuesta activa, como intentos de fuga; teniendo ambos niveles equivalentes de condicionamiento de miedo (Archer, 1975). Taylor et al., (2000), desde un modelo de mecanismo bioconductual, proponen que los machos adoptan la respuesta típica de lucha o huída, mientras que las hembras muestran una respuesta más defensiva y pasiva. Adicionalmente, se ha determinado que los estrógenos juegan un papel importante en la modulación del condicionamiento de miedo, al igual que el desarrollo ontogenético de los individuos y sus cambios morfológicos de estructuras neurológicas importantes en la respuesta de miedo que acompañan a estos cambios ontogenéticos (Andersen, 2003).
A diferencia del papel jugado por las diferencias sexuales en la adquisición de la respuesta de ansiedad, los determinantes asociados al desarrollo ontogenético han recibido menos atención en la literatura. Esto es especialmente cierto, en lo que respecta a la adolescencia, como etapa evolutiva, donde los cambios a nivel hormonal llevarían a pensar en alteraciones en la respuesta condicionada de miedo. La adolescencia es la etapa de transición y preparación para la edad adulta, en esta etapa, es más notable la expresión de trastornos comportamentales (Kessler, Chiu, Demler & Walters, 2005). Esta etapa se caracteriza también por una mayor prevalencia de conductas de riesgo y la búsqueda de novedad e interacción social con pares, lo cual ha llevado a suponer que la adolescencia es una época relativamente estresante, caracterizada por mayor ansiedad y mayor estrés en comparación con la edad adulta (Domerus-Fitzwater, Varlinskaya & Spear, 2009). Estos patrones comportamentales, se manifiestan en diferentes especies, como en roedores de laboratorio (Viveros, 2009).
Existen algunos estudios preclínicos que han explorado el efecto de la evolución ontogenética sobre el condicionamiento del miedo. Brasser y Spear (2004), por ejemplo, compararon el condicionamiento aversivo al contexto en ratas infantes, juveniles, adolescentes y adultas bajo presentaciones apareadas y explícitamente desapareadas del EC y el EI. Estos autores encontraron que el apareamiento EC-EI ante el contexto, los adultos y adolescentes se condicionan eficazmente, pero sólo en los infantes y juveniles este arreglo producía condicionamiento aversivo al contexto. Por el contrario, en adolescentes y adultos, el condicionamiento contextual era apreciable sólo en la presentación explícitamente desapareada del EC y el EI. Por tanto, las diferencias en el aprendizaje a lo largo del desarrollo, se deben a la forma en que se da este, de tal manera que los infantes tienden aprender más sobre el valor predictivo de un estímulo individual que si ese estímulo ha predicho el refuerzo, en otras palabras, aprenden más cuando un estímulo compuesto (presentación simultánea de dos estímulos) predicen un evento importe, mientras para los adultos esto genera dificultad, ya que dada la experiencia previa, aprenden mas si el estímulo solo predice el refuerzo que sobre el valor predictor de un estímulo individual.
Resultados similares fueron reportados por Esmoris-Arranza, Mendez y Spear (2008), quienes además evaluaron el efecto de las diferencias sexuales a lo largo del desarrollo, utilizando un procedimiento de condicionamiento pavloviano de miedo. Estos autores, encontraron que, las diferencias sexuales a lo largo del desarrollo son mas posibles y claras, en donde las hembras presentan mayor respuesta de congelamiento y supresión condicionada para con la intervención (procedimiento de extinción) que los machos en edades adultas más que los adolescentes.
Adicionalmente, estudios como los de Lynn y Brown (2010) muestran cómo los roedores en instrumentos como el campo abierto y laberinto elevado en cruz presentan mayor exploración (mayor actividad locomotora) y mayor tiempo en brazos abiertos, respectivamente, a medida que aumenta la edad. Este reporte estaría en concordancia con los resultados en donde se señala niveles bajos en la exploración de los adolescentes (Candland & Campbell, 1962) y opuesto a la postura en donde se asumen más riesgos y respuesta a la novedad en la adolescencia, en mayor medida que los adultos (Spear, 2000). Aunque en lo que respecta a la interacción edad–sexo, no se evidenciaron diferencias claras.
A partir de los anteriores reportes, la finalidad de este estudio fue evaluar la adquisición de la respuesta de miedo condicionado (congelamiento) entre hembras y machos en dos edades: adolescentes día Postnatal (DPN) 30 y adultos DPN 90, desde un modelo preclínico de estrés postraumático en roedores.
Método
Sujetos
Se utilizaron 45 ratas Wistar, 18 machos y 27 hembras, provenientes de 7 camadas, las cuales fueron balanceadas entre los grupos experimentales a fin de evitar las correlaciones espurias entre los miembros de un mismo grupo. Los animales fueron adquiridos en el bioterio de la empresa Inmunopharmos cuando tenían 21 días postnatales, llevados al bioterio de la Fundación Universitaria Konrad Lorenz en donde estuvieron alojados durante todo el experimento. El número de sujetos por edad y sexo se presentan en la tabla 1.

El lugar de alojamiento fue una habitación sin ventanas, en cajas de polipropileno opacas marca Tecniplast, con fase de luz de 12-h/12-h del ciclo luz/día (luces encendidas 6:00-18:00, la temperatura: 23.2°C y humedad relativa: 49.2% en promedio). Las ratas, tenían acceso ad libitum al alimento y al agua a lo largo de todo el experimento. En el día post-natal 28 fueron destetados y alojados en pares o tríos del mismo sexo pertenecientes a la misma camada que fueron asignados aleatoriamente a los grupos experimentales. Los experimentos fueron corridos en dos cohortes, por limitaciones en la cantidad de equipos experimentales, además las cohortes fueron balanceadas entre los grupos experimentales.
Cada animal se manipuló sólo para transportarlo a la sala experimental, para la toma de peso diario y la limpieza de la caja semanal. Las sesiones experimentales se llevaron a cabo una vez los animales cumplían 30 DPN o 90 DPN. Todos los procedimientos fueron aprobados y supervisados por el Comité Institucional para el Cuidado y Uso de Animales de Laboratorio (CICUAL) de la Fundación Universitaria Konrad Lorenz.
Instrumentos
Se utilizaron dos cajas de condicionamiento de miedo marca MED-Associates. Cada caja experimental consta de dos paredes laterales de aluminio y una pared frontal de plexiglás transparente, con dimensiones de 25 cm alto, 33 cm ancho y 32.5 cm de largo. Las cajas están dotadas de una iluminación de luz blanca, una iluminación cuasi-infra roja, un amplifcador y un parlante. El piso de las cajas están compuesto de varillas paralelas de hierro, ubicadas a 0.5 cm la una de la otra, permitiendo administrar descargas eléctricas alternadas. La presentación de los estímulos, las grabaciones de la sesión y los cálculos de los índices de movimiento, se realizaron de forma automática, mediante una computadora utilizando Video-Freeze Software (Asociados Med, Albany, VT) en tiempo real, a una velocidad de 30 fotogramas por segundo (véase figura 1).

Adjuntamente, las rejillas del suelo se lavaron con agua y jabón después de cada sesión con cada animal y las paredes de las cámaras experimentales se limpiaron con toallitas desinfectantes Clorox®.
Procedimiento
Todos los animales se pesaron al inicio de la sesión experimental y recibieron una sesión de condicionamiento Pavloviano de miedo de 7 minutos (min.) de duración. La sesión de condicionamiento iniciaba con una línea base de 120 segundos (s.), que sirvió como periodo de adaptación al instrumento. Seguidamente, se presentaron tres apareamientos del Estímulo Condicionado (EC–luz blanca intermitente que duró 10 s.) y el Estímulo Incondicionado (EI– Descarga eléctrica de 0.7 mA. y 1 s. de duración). El arreglo de condicionamiento fue de tipo demorado, en donde el EI y el EC terminaban simultáneamente. El intervalo entre ensayos fue de 90 s. entre apareamientos.
La cámara experimental se iluminó exclusivamente con la luz cuasi-infrarroja y cuando se presentaba el EC. Al finalizar las sesiones experimentales, todos los sujetos se sometieron a eutanasia mediante CO2, de acuerdo con el procedimiento estándar recomendado por el American Veterinary Medical Association (A.V.M.A., 2007) y realizado por la Dirección de Salud y Bienestar animal de la Fundación Universitaria Konrad Lorenz.
Resultados
Al evaluar la interacción de las variables sexo y edad sobre la adquisición de la respuesta de congelamiento, se identificaron los efectos de estas y su interacción sobre las tres mediciones de la variable dependiente (EC1, EC2 y EC2), por medio del análisis de un modelo de efectos. Adicionalmente, se evaluaron las diferencias en cada una de las mediciones del EC, permitiendo conocer la adquisición de la respuesta de congelamiento por sexo y edad.
La primera presentación del EC, previo al primer choque, se evidenció un porcentaje promedio de congelamiento bastante bajo para todos los sujetos, solo se evidenció una leve inmovilidad en las hembras adolescentes de 2% (ver figura 2a), los cual no resultó estadísticamente significativo. Adicionalmente las varianzas fueron iguales (F (3, 44) = 3.85, p < .16) entre sexo y edad en la respuesta de congelamiento, precisando que no existieron diferencian entre sexo y edad al inicio de la sesión, por lo cual los cambios que se presentaron más adelante en esta sesión se debieron a los apareamiento entre EC-EI y no a otras variables que pudieran influenciar el condicionamiento de miedo.
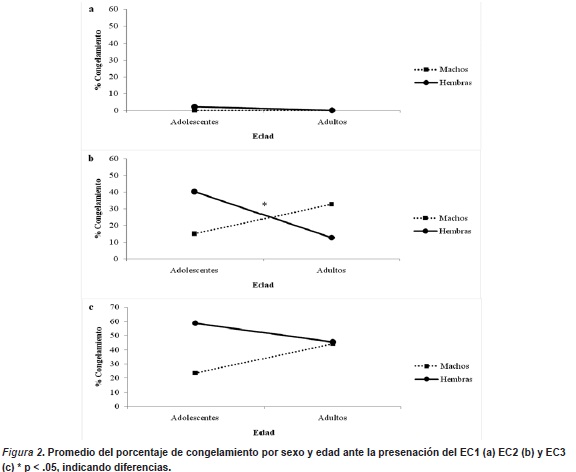
Para la segunda presentación del EC, se observó aumento en la respuesta de congelamiento en todos los sujetos. Siendo las hembras adolescentes las de mayor porcentaje de congelamiento (40%), seguido por los machos adultos (33%), luego los machos adolescentes (15%) y por último las hembras adultas (12%) (Ver figura 2b). Los efectos principales de la interacción entre sexo y edad son significativos (F (1, 44) = 5.43, p > .025), contrario a los efectos de cada variable por separado. Por tanto, la respuesta de miedo para la segunda presentación del EC dependió de los efectos de la interacción del sexo y edad de los sujetos.
Contrario en el tercer apareamiento, en donde no se evidenciaron efectos del sexo y de la edad, ni la interacción de ambas, sobre la respuesta de congelamiento. Aunque para todos los sujetos aumentó el nivel de congelamiento, las hembras adolescentes mantuvieron el mayor promedio de porcentaje de congelamiento (59%), seguido por hembras adultas (45%), muy cerca los machos adultos (44%) y en menor porcentaje los machos adolescentes (24%) (Ver figura 2c).
Con respecto a la adquisición de la respuesta de congelamiento en las tres mediciones, en interacción con el sexo y la edad, se evidenció que las hembras adolescentes adquirieron mayor respuesta en las tres mediciones, con respecto a los demás sujetos. Los machos adultos mostraron el segundo nivel de mayor adquisición del porcentaje, pero en la tercera presentación del EC su nivel de congelamiento fue menor que las hembras adultas, quienes iniciaron con un nivel de congelamiento inferior que todos los sujetos, pero a partir de la segunda presentación del EC, aumentó su porcentaje sobrepasando en 1 a los machos adultos y a los machos adolescentes en un 21 ante la tercera medición del EC. Los machos adolescentes fueron los de menor congelamiento en la última presentación del EC, pero se observaron niveles superiores de 2% en la segunda presentación del EC en comparación a las hembras adultas.
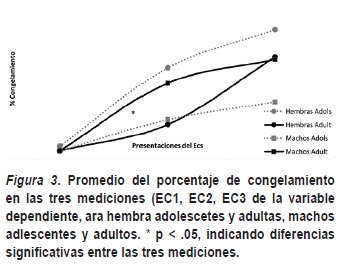
Con el ANOVA de mediciones repetitivas se encontraron diferencias significativas al comparar las tres mediciones de congelamiento ante el EC, F (2, 44) = 27.09, p > .00) (ver figura 3), mostrando que durante los tres apareamientos se presentó una adquisición de la respuesta de congelamiento, aumentando a partir del primer apareamiento, para todos los sujetos, aunque no se encuentran diferencias significativas a nivel sexual y edad entre las tres mediciones del EC. Por tanto, se puede concluir que existió adquisición de congelamiento para todos los sujetos durante la sesión experimental debido a los cambios evidenciados en cada uno de estas mediciones, aunque no se encontró diferencias sexuales y por edad en el total de la sesión de la adquisición de congelamiento.
Discusión
Las diferencias en la adquisición de la respuesta de congelamiento en ratas se puede explicar a través del aprendizaje dado por la interacción entre el ambiente y el individuo, por la ontogenia (Moye & Rudy, 1985) y el sexo del organismo (Gupta, Sen, Diepenhorst, Rudick & Maren, 2001). Los resultados resaltan que en las tres mediciones del EC las hembras adolescentes presentaron mayor respuesta de congelamiento que las hembras adultas y los machos adolescentes y adultos aunque solo fueron significativas en la segunda presentación del EC.
Los hallazgos son congruentes con los resultados de diferentes estudio en los que se reporta que las hembras presentan mayor ansiedad que los machos, evaluando la respuesta de congelamiento en condicionamiento pavloviano de miedo (Aguilar et al., 2003, Markus & Zecevic, 1997; Pryce et al., 1999; Wiltgen et al., 2001) y en otras medidas de ansiedad, tomadas a partir de pruebas como campo abierto (Mashoodh, Sinal, & Perrot-Sinal, 2009; Olivier et al., 2008) y laberinto elevado en cruz (Bowman, Micik, Gautreaux, Fernandez & Luine, 2009; Moreno, Lamprea, & Dueñas, 2009). Aunque contrario a los resultados que evidencian en los machos niveles más altos de la respuesta de congelamiento que las hembras (Chang et al. 2009; Graham, Yoon, Lee, & Kim, 2009; Maren, DeOca, & Fanselow, 1994; Sorg, Swindell, Tshirgi, 2004) o estudios en los que no se observó diferencias sexuales significativas durante la adquisición de miedo (Imanaka et al., 2008; Milad et al., 2009).
Adicionalmente, las diferencias podrían explicarse de acuerdo a Taylor et al. (2000) en cuanto a que los machos ante situaciones de peligro presentan mayor número de conductas de lucha/huida debido a la producción de andrógenos (como la testosterona) que se generan antes y/o después del nacimiento del individuo. Esto desarrolla a nivel cerebral la agresión, que se precipita ante contextos de amenaza y posibilita gran actividad, lo cual podría señalar que al no estar congelado los machos posiblemente su conducta de ansiedad se evidenciarían más por conductas de actividad más que quietud.
Contrario, las hembras por ser característico de ellas, la necesidad de búsqueda de protección, tienden a presentar un comportamiento diferente al de lucha/huida y más defensivo/pasivo como lo es la inmovilización (Collaer & Hines, 1995), explicando los niveles bajos de movimiento registrados después del segundo apareamiento y mucho más ante la tercera medición del EC para las hembras en las adolescentes y las hembras adultas. Por ello, es importante evaluar modelos animales y respuestas conductuales que puedan evaluar conductas activas y de pasividad que permitan evaluar el miedo condicionado.
Por otra parte, la influencia del ciclo hormonal en la respuestas de miedo puede generar diferencias sexuales debido al ciclo reproductivo en que se encuentren las hembras al momento de la prueba de condicionamiento pavloviano de miedo (Chang et al., 2009; Markus & Zecevic, 1997) como en otras pruebas (Baker et al., 2009; Liang, Byers &. Irwin, 2008).
Toufexis (2007) señala que las respuestas más altas de miedo en hembras se producen durante el final del proestro (sexualmente receptivas) cuando las hormonas ováricas, estrógenos y progesterona están en su pico y no en dioestrous cuando estas mismas hormonas están bajas. Aunque, Milad, et al. (2009) no reportan diferencias en el ciclo hormonal de las hembras en la adquisición de miedo. Y Gupta et al. (2001) señala que ratas hembras ovarectomizadas encontró un nivel de congelamiento similar a los machos y significativamente menor que el de ratas hembras sin operar. Estos estudios resaltan la importancia de identificar el ciclo hormonal de los sujetos, aunque en la presente investigación no se identificó el ciclo estral, se presumió que se encontraba sincronizado entre las hembras, debido a que eran de dos cohortes y fueron criadas en un mismo contexto y alojadas por camadas bajo las mismas condiciones medioambientales. Evidenciándose en la primera medición de la sesión experimental, en donde todos los sujetos presentaron patrones similares de respuesta de congelamiento, excepto por una leve inmovilidad de las hembras adolescentes, pero que no fue significativo para con los demás sujetos. Por lo cual, se podría afirmar que los cambios hormonales no influenciaron de manera significativa en las mediciones realizadas, aunque no se descarta que los cambios hormonales y por ende el ciclo estral influyan en el congelamiento, se recomienda en futuros estudios tener en cuenta esta variable en las hembras al momento de evaluarlas.
Con respecto a la edad, se ratifican los resultados de Brasser y Spear (2004) y Esmoris-Arranz et al. (2008), en los que resaltan que los adolescentes presentan mayor respuesta de miedo que los adultos, reportes similares se encuentran en pruebas exploratorias como campo abierto y laberinto elevado (Lynn & Brown, 2010). Esto se debe a que los adolescentes roedores presentan mayor ansiedad y estrés en comparación a los adultos (Domerus-Fitzwater et al., 2009). Igualmente Blumberg, Freeman y Robinson (2010), señalan que el congelamiento se observa más claro en edades tempranas.
Estudios que han evaluado la ontogenia, para determinar la adquisición de miedo condicionado han observado que ante estímulos condicionados como el tono, la retención se presentan a los 18 y 23 DPN incrementando significativamente a los 23 a 32 DPN (Rudy & Morledge, 1994) además se ha señalado que el aprendizaje en contexto y entre EC (tono)-EI en la infancia (17-19 DPN) es más fuerte que en la adolescencia (35 DPN) y en la adultez (71-89 DPN). En el estudio Brasser & Spear (2004) los adolescentes en comparación a los adultos presentan condicionamiento contextual cuando se apareó el EC (tono)-EI, acorde con lo encontrado en los resultados, en donde los adolescentes tienden a presentar mayor respuesta de congelamiento que los adultos.
Estudios en donde evalúen las diferencias en sexo y en edad en condicionamiento pavloviano de miedo son pocos los reportes, uno de los más recientes y con resultados similares a los del presente estudio es el de Esmoris-Arranz et al. (2008); quienes evaluaron la eficacia del condicionamiento de miedo contextual en ratas, en tres periodos de la ontogenia: infancia tardía (17 DPN), adolescentes (28-31 DPN) y adultez temprana (50-70 DPN), utilizando un estímulo olfativo como estímulo condicionado. Encontraron similitud con los resultados, en cuanto a que los adolescentes presentaron una fuerte disposición de aprendizaje contextual tanto en apareamiento como cuando no hay apareamiento, además las hembras presentaron menor actividad que los machos.
Teniendo en cuenta el tipo de EC, se encuentra el estudio de Moye y Rudy (1985), quienes hallaron que ante la presentación apareada luz-choque se establece una dependencia con la edad, evidenciándose que los sujetos de 17 DPN suprimen la actividad en presencia de la luz pero a mayor edad mayor la supresión. Estando en congruencia con los resultados encontrados, en cuanto a que todos los sujetos presentan supresión de la conducta aunque no aumenta con la edad.
Por otra parte, es de gran relevancia conocer la adquisición de miedo ante no asociaciones del EC-EI y las posibles diferencias sexuales y en edad que puedan aclarar y enriquecer más la literatura en modelos preclínicos, e igualmente evaluar la respuesta de congelamiento en otras especies que puedan evidenciar patrones más similares a los humanos de acuerdo al estudio de Taylor et al. (2000).
Además, es indispensable seguir abordando otras etapas del desarrollo, como edades mucho menor a 35 DPN, de acuerdo a los resultados de Esmoris-Arranz et al. (2008) y Moye y Rudy (1985), intermedias entre 35 y 90 DPN o superiores a 90 DPN, que permitan identificar claramente las edades de adquisición de miedo condicionado ante un estímulo condicionado.
Igualmente, es de gran importancia conocer el proceso de extinción en grupos con asociación y no asociación entre estímulos por edad y sexo, tema que poco se ha abordado hasta el momento (Esmoris-Arranz et al., 2008) y que es la explicación de los modelos terapéuticos para TEPT (Gómez et al. 2002; Milad et al. 2006).
Referencias
A.V.M.A. (2007). AVMA Gudelines on Euthanasia. EE.UU.: 36. [ Links ]
Aguilar, R., Gil, L., Gray, J. A., Driscoll, P., Flint, J., Dawson, G. R., Giménez-Llort, L., Escorihuela, R. M., Fernández-Teruel, A., & Tobeña, A. (2003). Fearfulness and sex in F2 Roman rats: males display more fear though both sexes share the same fearfulness traits. Physiology & Behavior, 78, 723-732. [ Links ]
Alonso, J., Angermeyer, M. C., Bernert, S., Bruffaerts, R., Brugha, T.S., Bryson, H., Girolamo, R., Graaf, K., Demyttenaere, I., Gasquet, J.M., Haro, S.J., Katz, R.C., Kessler, V., Kovess, J.P., Lèpine, J., Ormel, G., Polidori, L.J., Russo, G., Vilagut, J., Almansa, S., Arbabzadeh-Bouchez, J., Autonell, M., Bernal, M.A., Buist-Bouwman, M., Condony, A., Domingo-salvany, M., Ferrer, S.S., Joo, M., Martínez-Alonso, H., Matschinger, F., Mazzi, Z., Morgan, P., Morosini, C., Palacín, B., Romero, N., Taub, W.A., & Vollebergh, W.A.M. (2004). Prevalence of mental disorders in Europe: Results from the European study of the epidemiology of mental disorders (ESEM) Project. Acta Psychiatrica Scandinavica, 109 (suppl. 420), 21-27. [ Links ]
American Psychiatric Association. (2002). DSM-IV Manual diagnóstico y estadístico de los transtornos mentales. Barcelona: Masson. [ Links ]
Andersen, S. L. (2003). Trajectories of brain development: point of vulnerability or window of opportunity?. [ Links ]
Archer, J. (1975). Rodent sex differences in emotional and related behavior. Behavioral Biology, 14, 459–479. [ Links ]
Arenas, M. C. & Puigcerver, A. (2009). Diferencias entre hombres y mujeres en los trastornos de ansiedad: una aproximación psicobiológica. Escritos de Psicología, 3(1), 20-29. [ Links ]
Baker, S., Rees, S., Chebli, M., LeMarec, N., Godbout, R., Huta, V., & Bielajew C. (2009). Effects of gestational stress: 2. Evaluation of male and female adult offspring. Brain Research, 1302, 194-204. [ Links ]
Baran, S. E., Armstrong, C. E., Niren, D. C., Hanna, J. J., & Conrad, C. D. (2009). Chronic stress and sex differences on the recall of fear conditioning and extinction. Neurobiology of Learning and Memory, 91, 323-332. [ Links ]
Blumberg, M. S., Freeman, J. H., & Robinson, S. R. (2010). Ontogeny of gear conditioning, Oxford handbook of developmental behavioral neuroscience. New York: OXFORD University Press. [ Links ]
Bolles, R. C. (1970). Species-specific defense reactions and avoidance learning. Psychological Review, 77, 32–48. [ Links ]
Bouton, M. E. & Bolles, R. C. (1980). Conditioned fear assessed by freezing and by the suppression of three different baselines. Animal Learning Behavior, 8, 429-434. [ Links ]
Bowman, R. E., Micik, R., Gautreaux, C., Fernandez, L., & Luine, V. N. (2009). Sex-dependent changes in anxiety, memory, and monoamines following one week of stress. Physiology & Behavior, 97(1) 21-29. [ Links ]
Brasser, S.M., & Spear, N.E. (2004). Contextual conditioning in infants, but not older animals, is facilitated by CS conditioning. Neurobiology Learning Memory, 81, 46–59. [ Links ]
Candland, D. K., & Campbell, B. A. (1962). Development of fear in the rat as measured by behavior in the open field. Journal of Comparative and Physiological Psychology, 55, 593–598. [ Links ]
Chang, Y.J., Yang, C.H., Liang, Y.C., Yeh, C.M., Huang, C.C., & Hsu, K. S. (2009). Estrogen modulates sexually dimorphic contextual fear extinction in rats through estrogen receptor β. Hippocampus, 19, 1142-1150. [ Links ]
Collaer, M. L., & Hines, M. (1995). ¿Human behavioral sex differences: a role for gonadal hormones during early development?. Psychological Bulletin, 118(1), 55-107. [ Links ]
Davis, M. (1992). The role of the amygdala in conditioned fear. See aglleton, 255-306. [ Links ]
Diehl, L. A., Silveira, P. P., Leite, M. C., Crema, L. M., Portella, A. K., Billodre, M. N., Nunes, E., Henriques, T. P., Fidelix-Da-Silva, L. B., Heis, M. D., Goncalves, C. A., Quillfeldt, J. A., & Dalmaz, C. (2007). Long lasting sex-specific effects upon behavior and S100b levels after maternal separation and exposure to a model of post-traumatic stress disorder in rats. Brain Research, 1144, 107-116. [ Links ]
Domjan, M. (2005). Pavlovian conditioning: a functional perspective. Annual Review Psychology, 56, 179-206. [ Links ]
Doremus-Fitzwater T. L., Varlinskaya, E. I., & Spear, L. P. (2009). Effects of pretest manipulation on elevated plus-maze behavior in adolescent and adult male and female Sprague-Dawley rats. Pharmacology Biochemistry and Behavior, 92(3), 413-423. [ Links ]
Echenique, C., Medina, L. M., Medina, A. R., & Ramírez, A. (2008). Prevalencia del trastorno por estrés postraumático en población desplazada por violencia, en proceso de restablecimiento en Sincelejo. Psicología desde el Caribe, Universidad del Norte, 21, 122-135. [ Links ]
Esmoris-Arranz, F. J., Mendez, C., & Spear, N. E. (2008). Contextual fear conditioning differs for infant, adolescent, and adult rats. Behavioural Processes, 78(3), 340-350. [ Links ]
Fanselow, M. S. (1980). Conditional and unconditional components of postshock freezing. Pavlovian Journal Biological Sciences, 15, 177-82. [ Links ]
Fanselow, M. S., & Tighe, T. J. (1988). Contextual Conditioning with massed versus distributed unconditional stimuli in the absence of explicit conditional stimuli. Journal of Experimental Psychology, 14(2), 187-199. [ Links ]
Fanselow, M. S., DeCola, J. P., & Young, S. L. (1993). Mechanisms responsable for reduced contextual conditioning with massed unsignaled unconditional stimuli. Journal of Experimental Psychology, 19(2), 121-137. [ Links ]
Gómez, C., Saldívar-González, J. A., & Rodríguez, R. (2002). Modelos animales para el estudio de la ansiedad: una aproximación crítica. Salud Mental, 25(1), 14-24. [ Links ]
Graham, L. K., Yoon, T., Lee, H. J., & Kim J. J. (2009). Strain and sex differences in fear conditioning: 22 kHz ultrasonic vocalizations and freezing in rats. Psychology & Neuroscience, 2(2), 219-225. [ Links ]
Gupta R. R., Sen S., Diepenhorst, L. L., Rudick, C. N., & Maren, S. (2001). Estrogen modulates sexually dimorphic contextual fear conditioning and hippocampal long-term potentiation (LTP) in rats. Brain Research, 888(2), 356-365. [ Links ]
Ide S., Sora, I., Ikeda, K., Minami, M., Uhl, G. R., & Ishihara, K. (2010). Reduced emotional and corticosterone responses to stress in mu-opioid receptor knockout mice. Neuropharmacology, 58 (1), 241-247. [ Links ]
Imanaka, A., Morinobu, S., Toki, S., Yamamoto, S., Matsuki, A., Kozuru, T., & Yamawaki, S. (2008). Neonatal tactile stimulation reverses the effect of neonatal isolation on open-field and anxiety-like behavior, and pain sensitivity in male and female adult Sprague-Dawley rats. Behavioural Brain Research, 186(1), 91-97. [ Links ]
Kessler, R. C., Chiu, W. T., Demler, O., & Walters, E. E. (2005). Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the National Comorbidity Survey Replication. Archives of General Psychiatry, 62, 617-627. [ Links ]
Kohn, A., Koenen, K. C., & Jaffee, S. R. (2009). Post-traumatic stress symptoms and trajectories in child sexual abuse victims: an analysis of sex differences using the national survey of child and adolescent Well-Being. Journal Abnormal Child Psychology, 31, 727-737. [ Links ]
Kosten, T. A., Lee, H. J., & Kim, J. J. (2006). Early life stress impairs fear conditioning in adult male and female rats. Brain Research, 1087(1), 142–50. [ Links ]
LeDoux J. E. (1995). Emotion: Clues from the brain. Annual Review Psychology, 46, 209-235. [ Links ]
Liang S., Byers, D. M., & Irwin, L. N. (2008). Sex and diet affect the behavioral response of rats to chronic mild stressors. Physiology & Behavior, 93, 27-36. [ Links ]
Londoño, N. H., Muñiz, O., Correa, J. E., Patiño, C. D., Jaramillo, G., Raigoza, J., Toro, L., Restrepo, D. A., Rojas, C. (2005). Salud mental en víctimas de la violencia armada en Bojayá (Chocó Colombia). Revista Colombiana de Psiquiatría, XXXIV (004), 493-505. [ Links ]
Lynn, D. A. & Brown G. R. (2010). The ontogeny of anxiety-like behavior in rats from adolescence to adulthood. Developmental psychobiology, 1-9. [ Links ]
Maren, S., DeOca, B., & Fanselow, M. S. (1994). Sex differences in hippocampal long-term potentiation (LTP) and Pavlovian fear conditioning in rats: positive correlation between LTP, contextual learning. Brain Research, 661(1-2), 25–34. [ Links ]
Markus, E. J., & Zecevic M. (1997). Sex differences and estrous cycle changes in hippocampus-dependent fear conditioning. Psychobiology, 25(3), 246-252. [ Links ]
Mashoodh R., Sinal, C. J., & Perrot-Sinal, T. S. (2009). Predation threat exerts specific effects on rat maternal behaviour and anxiety-related behaviour of male and female offspring. Physiology & Behavior, 96, 693-702. [ Links ]
Milad M. R., Igoe, S. A., Lebron-Milad, K., & Novales, J. E. (2009). Estrous cycle phase and gonadal hormones influence conditioned fear extinction. Neuroscience, 164 (3), 887-895. [ Links ]
Milad, M. R., Goldstein, J. M., Orr, S.P., Wedig, M.M., Klibanski, A., Pitman, R. K., & Rauch, S. L. (2006). Fear conditioning and extinction: influence of sex and mestrual cycle in healthy humans. Behavioral Neuroscience, 120(5), 1196-1203. [ Links ]
Milad, M. R., Zeidan, M. A., Contero, A. Pitman, R. K., Klibanski, A., Rauch, S. L., & Goldstein, J. M. (2010). The influence of gonadal hormones on conditioned fear extinction in healthy humans. Neuroscience, 168, 652-658. [ Links ]
Mitra, R. & Sapolsky, R. M. (2010). Expression of chimeric estrogen-glucorticoid-receptor in the amygdala reduces anxiety. Brain Research, 1342, 33-38. [ Links ]
Moreno L, Lamprea, M & Dueñas, Z. (2009). Diferencias en los comportamientos asociados con la ansiedad de ratas macho y hembras expuestas a un protocolo de estrés crónico por separación maternal temprana. Suma Psicológica, 16(1), 31-43. [ Links ]
Moye, T.B., & Rudy, J.W. (1985). Ontogenesis of learning. VI. Learned and unlearned responses to visual stimulation in the infant hooded rat. Developmental Psychobiology, 18, 395–409. [ Links ]
Organización Mundial de la Salud (World Health Organization (WHO)). (2001). The world health report 2001, mental health: New understanding. New Hope: Geneva. [ Links ]
Palanza, P. (2001). Animal models of anxiety and depression: how are females different? Neuroscience and Biobehavioral Reviews, 25, 219-233. [ Links ]
Pavlov, I. P. (1927). Conditioned reflexes: An investigation of the physiological activity of the cerebral cortex (G. V. Anrep, Trans.). Oxford: Oxford University Press. [ Links ]
Pineda, Guerrero, Pinilla, & Estupiñán (2002). Utilidad de un cuestionario para rastreo del estrés postraumático en una población colombiana. Acta Neurológica Colombiana, 18 (3), 132-138. [ Links ]
Posada-Villa, J. A., Buitrago-Bonilla, J. P., Medina-Barreto, Y., & Rodiguez-Ospina, M. (2006). Trastorno de ansiedad según distribución por edad, género, variación por regiones, edad de aparición, uso de servicios, estado civil y funcionamiento/discapacidad según el tipo de estudio nacional. Red de Revistas Científicas de América Latina y el Caribe, España y Portugal, 4(006), 33-41. [ Links ]
Pryce, C. R., Lehmann, J., & Feldon, J. (1999). Effect of sex on fear conditioning is similar for context and discrete CS in Wistar, Lewis and Fischer rat strains. Pharmacology Biochemistry and Behavior, 64(4), 753–9. [ Links ]
Rau, V., & Fanselow, M. S. (2009). Exposure to a stressor produces a long lasting enhancement of fear learning in rats. Stress, 12(2), 125-133. [ Links ]
Rudy, J., & Morledge, P., (1994). Ontogeny of contextual fear conditioning in rats: implications for consolidation, infantile amnesia, and hippocampal system function. Behavior Neurosciences, 198, 227–234. [ Links ]
Sorg B. A., Swindell, S., & Tschirgi, M. L.(2004). Repeated low level formaldehyde exposure produces enhanced fear conditioning to odor in male, but not female, rats. Brain Research, 1008(1). [ Links ]
Spear, L. P. (2000). Neurobehavioral Changes in Adolescence. Current directions in psychological science, 9(4), 111-114. [ Links ]
Starkey N.J., & Bridges, N.J. (2010). The effects of acute, chrinic and withdrawn progesterone in male and female mongolian gerbils (meriones unguiculatus) in two tests of anxiety. Behavioural Brain Research, 207, 490-499. [ Links ]
Taylor, S. E., Cousino-Klein, L., Lewis, B. P., Gruenewald, T. L., Gurung, R. A. R., & Updegraff, J. A. (2000). Biobehavioral responses to stress in females: Tend-and-Befriend, not fight-or-fight. Psychological Review, 107(3), 411-429. [ Links ]
Tolin, D. F., & Foa, E. B. (2006). Sex differences in trauma and post-traumatic stress disorder: a quantitative review of 25 years of research. Psychological Bulletin, 132(6), 959-992. [ Links ]
Toufexis, D. (2007). Region-and sex-specific modulation of anxiety behaviours in the rat. Journal of neuroendocrinology, 19, 461-473. [ Links ]
Verma P., Hellemans, K. G.C., Choi, F. Y., Yu, W., & Weinberg, J. (2010). Circadian phase and sex effects on depressive/anxiety-like behaviors and HPA axis responses to acute stress. Physiology & Behavior, 99(3), 276-285. [ Links ]
Viveros, M. P. (2009). Presenta la mujer aspectos etiológicos y epidemiológicos diferenciales? Efectos sexodimórficos de los cannabinoides: ¿Qué nos dicen los modelos animales?. Jornada Cannabis, Mujer y Patología Dual. [ Links ]
Wiltgen B. J., Sanders, M. J., Behne, N. S., & Fanselow, M. S. (2001). Sex differences, contex preesposure, and the immediate shock deficit in pavlovian context conditioning with mice. Behavioral Neuroscience, 115(1), 26-32. [ Links ]

