










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSuma Psicológica
versión impresa ISSN 0121-4381
Suma Psicol. vol.19 no.1 Bogotá ene./jun. 2012
Instituto de Investigaciones Médicas (IDIM) - CONICET y Universidad Abierta
Interamericana, Argentina
Instituto de Investigación Médica Mercedes y Martin Ferreyra - CONICET y
Universidad Nacional de Córdoba, Argentina
Instituto de Investigaciones Médicas (IDIM) - CONICET y Universidad Abierta
Interamericana, Argentina.
La correspondencia relacionada con este artículo puede ser dirigida a Giselle Kamenetzky. Dirección postal: Combatientes de Malvinas 3150 (CP 1427), Buenos Aires, Argentina. Correo electrónico: yoselevich@hotmail.com
Recibido: Febrero 11 2012 Aceptado: Mayo 24 2012
Resumen
Este estudio evaluó la respuesta consumatoria de ratas infantes ante diferentes magnitudes de soluciones azucaradas y luego de su devaluación o extinción, con el objetivo de detectar la preferencia de las soluciones y la existencia de efectos de contraste y de magnitud del reforzamiento en la extinción. En el Experimento 1 se expuso ratas de 8-10 días postnatales (DPN) a diferentes concentraciones de sacarosa (12%, 10%, 5% ó 2%; fase de precambio) a las que se las devaluó al 2% (fase de postcambio). En el Experimento 2, dos grupos de 10-14 DPN recibieron soluciones al 12% ó 2% en la fase de precambio y 2% en la de postcambio. En ambos experimentos se observó que, durante la primera fase, el grupo 12% exhibió un mayor porcentaje de ganancia de peso que los demás grupos, pero no hubo diferencias entre los grupos en la fase de post-cambio. En el Experimento 3 se evaluó el efecto de magnitud del refuerzo en la extinción en ratas de 7-12 DPN, utilizando agua en la segunda fase. Se halló un efecto de magnitud del refuerzo en la adquisición, sin embargo, no hubo diferencias entre grupos en la segunda fase. En el Experimento 4 se utilizó un procedimiento de contraste anticipatorio (CA) con ratas de 10-16 DPN. Los animales exhibieron nuevamente un efecto de magnitud del refuerzo pero no se observó CA. Estos resultados indican que: (a) al menos desde los 8 DPN las ratas discriminan diferentes concentraciones de sacarosa y que prefieren la del 12%, y que (b) hasta la segunda semana de vida los animales consumen los reforzadores en función de su valores absolutos. Los resultados se discuten en términos de la ontogenia del aprendizaje de los efectos paradójicos del reforzamiento y su vinculación con la teoría de Amsel.
Palabras clave: Magnitud, devaluación, ratas, ontogenia.
Abstract
This study evaluated consummatory responses in infant rats exposed to different magnitude of reward, and after the devaluation (i.e., consummatory successive negative contrast) or omission (i.e., extinction) of reward. In Experiment 1, 8-10 post-natal days (PND) pups were intraorally infused with 12%, 10%, 5% or 2% sucrose (preshifit phase, 2 daily trials). Subsequently, all groups received 2% sucrose (postshift phase). In Experiment 2, 10-14 day-old pups received 12% or 2% sucrose in 4 daily trials in the preshift phase, followed by 2% in a postshift trial. Both experiments indicated that during preshifit, animals exposed to 12% sucrose exhibited higher sucrose consumption than those receiving lower concentration solutions. This phenomenon, indicative of a magnitude of reinforcement effect was not accompanied by evidence of successive negative contrast. In Experiment 3 we evaluated the magnitude of reinforcement extinction effect in 7-12 PND rats. Animals received 12% sucrose or water in preshifit phase and both groups received a neutral solution (i.e., water) in the second phase. A magnitude of reinforcement acquisition effect was again observed, yet there were no differences between groups in extinction phase. In Experiment 4 we used an anticipatory contrast procedure in 10-16 PND pups. A magnitude of reinforcement, but not an anticipatory contrast effect was observed. Overall the results indicate that: (a) rats from - at least- 8 PND discriminate between different concentrations of sucrose, and (b) until the second week of life the response to rewards is mainly regulated by their absolute value and not by their relative value. Results are discussed terms of the ontogeny of paradoxical effects of reward and its relationship to Amsel's theory.
Keywords: Magnitude, devaluation, rats, ontogeny.
Existen ciertos fenómenos del aprendizaje que acaparan el interés de los investigadores debido a sus implicancias teóricas y aplicadas. Se trata de los efectos paradójicos del reforzamiento (EPR) o efectos de contraste, desarrollados ampliamente por la teoría de la frustración de Amsel (1992) y por Gray (1987). Los EPR constituyen un conjunto de fenómenos del aprendizaje que tienen en común la pérdida o variación sorpresiva de recompensas. Se los llama "paradójicos" porque los sujetos que recibieron magnitudes o frecuencias mayores de incentivos exhiben, al recibir la omisión del reforzador o uno de menor magnitud, una respuesta conductual significativamente menor que aquellos que siempre recibieron el reforzador de menor magnitud. Las teorías clásicas del aprendizaje no pueden explicar estos fenómenos, ya que postulan que cuanto mayor sea el refuerzo recibido, mayor será la respuesta que se emite (Amsel, 1992; Amsel, & Stanton, 1980). Estos efectos sugieren que durante el aprendizaje los animales anticipan la clase de reforzador que van a recibir y cuando éste cambia, su respuesta está controlada no sólo por el valor absoluto del reforzador, sino por la relación entre lo que recibe y lo que esperaba recibir. Es por ello que también se los denominan relatividad de los refuerzos (Flaherty, 1996), aludiendo a que los reforzadores tienen un valor absoluto y otro relativo a la historia previa de reforzamiento.
Los EPRs se estudiaron ampliamente en ratas adultas con procedimientos instrumentales, en los cuales se evaluó los cambios en la conducta instrumental (e.g., velocidad de recorrido en el corredor lineal o palanqueo en la caja de condicionamiento) y con procedimientos consumatorios (e.g., consumo del reforzador, lameteo o tiempo en contacto con un bebedero que contiene reforzadores líquidos). En este último paradigma la repuesta aprendida es el consumo o medidas indirectas del mismo porque describe una curva ascendente y asintótica en función de los ensayos, de lo que se infiere que los animales optimizan la respuesta consumatoria. En ratas infantes los ERPs sólo se estudiaron con procedimientos instrumentales. En este trabajo se presentarán los primeros experimentos con ratas infantes que evaluarán la presencia o no de cuatros fenómenos de aprendizaje usando procedimientos consumatorios: el efecto de magnitud del refuerzo en la adquisición consumatoria (EMRAc), el efecto de magnitud del reforzamiento en la extinción consumatoria (EMREc), el contraste sucesivo negativo consumatorio (CSNc) y el contraste anticipatorio (CA). El EMRA es un fenómeno no paradójico que consiste en la observación de una respuesta diferencial en función de la magnitud del refuerzo (Stanton & Amsel, 1980). Por ejemplo las ratas corren más velozmente en el corredor lineal cuando reciben altas magnitudes de refuerzo respecto de las bajas o consumen más de un reforzador preferido en procedimientos consumatorios. El EMRE consiste en observar una mayor resistencia a la extinción en sujetos que previamente fueron expuestos a magnitudes bajas del reforzador cuando se los compara con aquellos que recibieron magnitudes altas en la fase de adquisición (Brudette, Brake, Chen & Amsel, 1976; Chen, Gross & Amsel, 1981).
En el contraste sucesivo negativo (CSN) se observa una disminución abrupta de la respuesta tras experimentar un cambio sorpresivo de un reforzador de alta magnitud/calidad por otro de baja magnitud/calidad. Usualmente, la disminución se produce por debajo del nivel de un grupo control que siempre recibió el reforzador de menor valor (Chen, Gross & Amsel, 1981; Stanton & Amsel, 1980). El contraste anticipatorio (CA) consiste en cambiar la magnitud del reforzador en una misma sesión. En este caso, los animales que reciben una magnitud baja del reforzador (e.g., 4% de solución azucarada) consumirán menos en sucesivas sesiones si inmediatamente después se exponen a una más preferida (e.g., 32% de solución azucarada), comparado con sujetos que en las dos fases reciben 4% de solución azucarada (Flaherty, 1996). Existen evidencias de que el CA está causado por mecanismos asociativos pavlovianos, donde la sustancia inicial funciona como un estímulo condicionado que señala la presentación de otra sustancia preferida o estímulo incondicionado (Flaherty, Turovsky & Krauss, 1994).
Algunos antecedentes realizados con ratas adultas sugieren divergencias en los resultados de ERPs instrumentales y consumatorios por lo que podrían estar regulados por diferentes vías neurales (Mustaca et al., 2005). Por ejemplo, en nuestro laboratorio mostramos un efecto invertido del EMREc y del efecto de reforzamiento parcial con un procedimiento consumatorio (Mustaca, Freidin & Papini, 2002). Por otra parte, las lesiones hipocampales eliminan el CSN instrumental (CSNi) (Salinas & White, 1998) pero no el CSNc (Ver Flaherty, 1996) y las lesiones en el tálamo gustativo eliminan el CSNc pero no el CSNi (Sastre & Reilly, 2006).
Los antecedentes sobre la ontogenia del desarrollo cerebral de la rata indican que las células granulares del giro dentado (sistema septohipocampal) comienzan el proceso de diferenciación con mayor rapidez entre los 12-14 DPN, alcanzando los niveles adultos alrededor de los 25-30 DPN (Amsel & Stanton, 1980). En cambio el complejo amigdalino alcanzaría el punto máximo de desarrollo neuronal alrededor del DPN 14. Luego de esta edad no se observan importantes modificaciones (Berdel, Moryś & Maciejewska, 1997). Es probable que en los EPR consumatorios esté involucrada la memoria de reconocimiento y en los instrumentales la memoria de evocación (Mustaca et al., 2009), por lo que los ERPs consumatorios podrían aparecer antes que los EPR instrumentales.
Desde una perspectiva ontogenética, los EPR han sido estudiados escasamente en bebés humanos y ratas infantes. Las evidencias con humanos muestran que habría una regulación emocional de las respuestas a edades tempranas. Mast et al. (1980) hallaron la existencia de CSNi cuando a bebés de 82-112 días le cambiaron un móvil colgante de 10 ó 6 sonajeros a uno de 2. Por otra parte, Kobre y Lipsitt (1972) hallaron un efecto de CSNc sobre el chupeteo en bebés de 4 y 10 hs. Estos resultados sugieren que la primera reacción a la frustración en humanos es una respuesta no aprendida.
La evaluación de la ontogenia del aprendizaje en ratas permite manipulaciones y un control experimental imposibles de realizar con sujetos humanos (Kamenetzky & Mustaca, 2005). Además, tiene la ventaja de que el desarrollo cerebral no está completo al momento del nacimiento, lo que brinda la posibilidad de inferir las estructuras cerebrales necesarias para la existencia de determinados fenómenos de aprendizaje.
Los estudios ontogenéticos del aprendizaje en general y de los ERP en particular en la rata fueron llevados a cabo principalmente por Amsel entre 1950 a 1980 aproximadamente, usando exclusivamente procedimientos instrumentales con el corredor lineal, y la velocidad de recorrido como medida dependiente. Sus trabajos muestran que los EPRs aparecen entre los 10 y 63 días post natales (DPN) de la rata, expresándose en un orden específico que parece estar relacionado con el desarrollo cerebral. A edades más tempranas aparecen fenómenos relativamente más simples, no paradójicos, como el condicionamiento clásico e instrumental (Amsel & Stanton, 1980). El EMRA lo halló a los 11 DPN (Stanton & Amsel, 1980) y el EMRE, a partir de los 18 DPN, utilizando alimento sólido en diferentes magnitudes (Brudette, Brake, Chen & Amsel, 1976). En cuanto al CSNi, Chen, Gross y Amsel (1981) hallaron que se expresa de forma contundente a los 34-35 DPN utilizando comida sólida como reforzador en la caja meta y a los 25-26 DPN si se utiliza leche en un recipiente. El CA no fue evaluado en ratas infantes.
Para que se expresen los ERPs es necesario que los animales discriminen y prefieran distintas magnitudes de los reforzadores, lo cual se infiere a través de las respuestas que emiten. El valor hedónico de los incentivos se puede inferir a través de las expresiones faciales (Ganchrow, Steiner & Daher, 1983) y por los ritmos de succión o consumo. La discriminación de sabores se estudió con neonatos humanos, quienes parecen distinguir entre diferentes soluciones al momento del nacimiento. Se halló que los ritmos de succión frente al agua y al cloruro de sodio son significativamente diferentes (Crook, 1978). También se encontró que la respuesta es diferencial en función de distintas concentraciones de sacarosa (15% vs. 5%) y glucosa, midiendo la presión de la lengua sobre un pezón artificial (Nowlis & Kessen, 1976), la cantidad de succiones por minuto (Engen & Lipsitt, 1974) o la duración de los episodios de succión usando un pezón artificial (Crook & Lipsitt, 1976). Desor, Maller y Turner (1976) mostraron además que los bebés discriminaban el agua y diferentes concentraciones de solución de sacarosa, glucosa, fructosa o lactosa midiendo su consumo en mililitros. En ratas infantes se observó que la succión de un pezón artificial es mayor cuando permite el acceso a soluciones dulces que cuando dispensa soluciones saladas o agridulces (Nizhnikov, Petrov, Varlinskaya & Spear, 2002). Los procedimientos de aprendizaje aversivo gustativo también sugieren que las ratas son capaces de procesar sabores a edades tempranas (Gemberling & Domjan, 1982; Gemberling, Domjan & Amsel, 1980) e incluso durante la vida prenatal (Mickley, Remmers-Roeber, Crouse & Peluso, 2000). Estos antecedentes sugieren que tanto en ratas como en humanos las capacidades de discriminar y mostrar prefierencia por sabores aparecen en estadios tempranos del desarrollo.
Teniendo en cuenta los antecedentes presentados, este trabajo tuvo como objetivo analizar la ontogenia de los efectos paradójicos del aprendizaje usando procedimientos consumatorios. La comparación entre los datos ya obtenidos en adultos y los obtenidos en infantes, sumados a los conocimientos sobre ontogenia del sistema nervioso central, pueden permitir inferir las estructuras neurales necesarias para la expresión de ERPs. En otras palabras, el abordaje ontogenético permite correlacionar un determinado fenómeno con la emergencia de cambios en el patrón normal de desarrollo neural. Esta estrategia se ha utilizado para analizar fenómenos tan diversos como la expresión diferencial de proteínas en abejas que cursan desarrollo normal de cuidadoras a forrajeras (García et al., 2009), y la correlación entre la ontogenia de ciertos neurotransmisores y la prefierencia por alcohol en ratas infantes (véase Pautassi et al., 2009). Específicamente, los siguientes experimentos evaluaron la existencia de EMRAc, el cual permitió detectar (i) si los animales discriminaban y preferían diferentes concentraciones de sacarosa, (ii) si la omisión del reforzador (EMREc) provoca un EMRE invertido similar al observado en las ratas adultas y (iii) si la presentación de una solución de concentración de sacarosa más baja produce CSNc. Se utilizó un paradigma de infusión intraoral que permite el consumo voluntario de fluidos en ratas infantes (ver Pautassi et al., 2008) y la ganancia porcentual de peso corporal en cada uno de los ensayos como variable dependiente. Esta variable toma en cuenta el consumo absoluto de las soluciones infundidas, relativizado al peso de cada animal, y ha sido extensamente utilizada en numerosas investigaciones sobre la ontogenia del aprendizaje y los procesos de discriminación y aceptación de reforzadores (Hall & Rosenblatt, 1977; Gregg, Kittrell, Domjan & Amsel, 1978; Gemberling, Domjan, & Amsel, 1980; Miller, Hunt, & Spear, 1989; Hoffmann, Hunt & Spear, 1991; Nizhnikov, Petrov, Varlinskaya, & Spear, 2002; Chotro & Alonso, 2003; Arias & Chotro, 2005; Pautassi, Arias, Molina, & Spear 2008; Pautassi, Molina & Spear, 2008; Arias, Pautassi, Molina, & Spear, 2010). El Experimento 1 evaluó ratas de 8 a 10 DPN en el EMRAc y CSNc, el Experimento 2, CSNc en ratas de 10-14 DPN, el Experimento 3 EMRAc y EMREc a los 7-12 DPN y el Experimento 4, el CA a los 10-16 DPN.
Experimento 1. EMRAc y CSNc - DPN 8 a DPN 10.
Método
Sujetos. Se utilizaron 19 ratas hembras Wistar, representativas de 5 camadas, de 8 DPN al comienzo del experimento nacidas y criadas en el Instituto de Investigaciones Médicas A. Lanari (IDIM), Buenos Aires, Argentina, bajo condiciones de temperatura (23°C ±1) y humedad constantes. El ciclo de luz/oscuridad fue de 12 horas (fase de luz a partir de las 7 am). El día de nacimiento se consideró DPN 0.
Aparatos y Materiales. Se utilizó una bomba de infusión (KD Scientific, Holliston, MA) equipada con 8 jeringas Prexajet de 5 ml cada una. El volumen total de infusión fue, siguiendo parámetros de trabajos previos (Pautassi, Arias, Molina Spear, 2008), equivalente al 2.5% del peso de cada animal. La infusión se realizó de modo continuo durante 10 minutos. Las jeringas contenían solución azucarada al 12%, 10%, 5% ó 2%. Todas las concentraciones se prepararon diluyendo azúcar comercial en agua de red hasta alcanzar 100 ml de solución total. Para la solución al 12% se diluyó 12 g de azúcar, 10 g de azúcar para la de 10 %, 5 g de azúcar para la de 5 % y 2 g de azúcar para la de 2 %. Cada jeringa se conectaba a un tubo de polietileno PE 50 y ésta a su vez a una cánula PE10 (Clay Adams, Parsippany, NJ) colocada previamente en la mejilla del animal. Las cánulas fueron construidas utilizando el calor de un soldador con el fin de aplanar uno de sus extremos sin que se obstruya. Las cajas de entrenamiento consistían en 2 cajas de 28 x 15 x 15 cm cada una, construidas en acrílico transparentes. Cada una estaba subdividida en 4 compartimientos de 7 x 15 x 15 cm cada uno. Debajo de ellas se colocó una almohadilla térmica para mantener la temperatura constante de las crías (35.0 ± 0.5 °C), con una toalla de papel secante colocada arriba de la almohadilla.
Procedimiento. Se formaron 4 grupos: 12%, 10%, 5% ó 2% (la nomenclatura remite al porcentaje de solución azucarada recibido durante la primera fase del estudio). Tres horas antes de comenzar cada ensayo, las crías fueron separadas de sus madres y canuladas a fin de administrar las soluciones azucaradas dentro de la cavidad oral. Durante la privación materna las crías fueron mantenidas en grupos en una caja de acrílico negro de 24,5 x 20 x 22 cm, mantenida a 22-23 °C por medio de una almohadilla térmica que se colocaba debajo de la caja. Para canular a los animales, una aguja de metal (30G C-KJECT, CK Dental Industries, Buenos Aires, Argentina) se unió al extremo libre de una cánula PE10 y se colocó en el interior de la cavidad bucal del animal, en la porción media de la mejilla, quedando el extremo aplanado dentro de la misma. La aguja posteriormente se retiraba. En cada ensayo se alternaba el orden de canulación entre la mejilla izquierda y derecha para preservar el tejido del animal. Este procedimiento requiere aproximadamente 15 seg por sujeto y no induce un estrés significativo en la crías (Spear, Specht, Kirstein, & Kuhn, 1989).
Luego de 3hs las ratas se estimulaban suavemente con un algodón en la zona anogenital para inducir micción y/o defecación. A continuación se las pesaban e inmediatamente la cánula se conectaba a la bomba de infusión comenzando el ensayo. El entrenamiento consistió de dos fases: 2 ensayos de precambio (DPNs 8-9) y 1 de postcambio (DPN 10), con un intervalo entre ensayos de 24 hs. En la primera fase las crías recibían una solución azucarada al 12% (n=6, Grupo 12-2), 10%,(n=5, Grupo 10-2), 5% (n=4, Grupo 5-2), ó 2% (n=4, Grupo 2-2), según el grupo asignado, en tanto que en la fase de postcambio todos los animales fueron infundidos con una solución al 2%. Las nomenclaturas de los grupos remiten a la solución recibida en cada fase. Por ejemplo, el Grupo 12-2 recibió la solución azucarada al 12% en la fase de precambio, y al 2% en el postcambio. Al finalizar cada ensayo se registraba nuevamente el peso de cada animal y las cajas de entrenamiento se limpiaban con un trapo humedecido en agua para neutralizar los olores. El entrenamiento se realizó entre las 13 y las 17 hs. La variable dependiente fue el porcentaje de ganancia de peso, el cual se obtuvo usando la siguiente fórmula: [(peso postinfusión - peso preinfusión) / peso preinfusión] x 100. (Hall & Rosenblatt, 1977; Gregg, Kittrell, Domjan, & Amsel, 1978; Gemberling, Domjan, & Amsel, 1980; Miller, Hunt, & Spear, 1989; Hoffmann, Hunt & Spear, 1991; Nizhnikov, Petrov, Varlinskaya & Spear, 2002; Chotro & Alonso, 2003; Arias & Chotro, 2005; Pautassi, Arias, Molina, & Spear 2008; Pautassi, Molina, & Spear, 2008; Arias, Pautassi, Molina, & Spear, 2010).
Resultados y discusión
En la Figura1 se grafica el porcentaje de ganancia de peso en función de los ensayos. Se observa que durante la fase de precambio los animales que recibieron reforzadores de mayor magnitud mostraron un porcentaje de ganancia de peso significativamente mayor que los que recibieron soluciones de menor concentración. Los resultados del ANOVA confirman esta observación. En la fase de precambio se realizó un ANOVA con un factor intersujeto (Grupo) y un factor intrasujeto (Sesiones 1 y 2). Se halló un efecto principal de Grupo, F (3, 15) = 5.47, p < .009. No se halló efecto principal de Sesión ni la interacción de ambos factores (p > .05). Para conocer las fuentes de las diferencias del factor grupo, se realizaron comparaciones poshoc con LSD Fisher. Este análisis arrojó diferencias estadísticamente significativas entre el Grupo 12-2 vs Grupo 10-2 (p < .04), así como con el Grupo 2-2 (p < .001) y marginalmente significativa con el Grupo 5-2 (p < .06) en el ensayo 2. El resto de los grupos no mostraron diferencias estadísticamente significativas entre sí (p > .05). Para la fase de postcambio se realizó un Anova de un factor intersujeto (Grupo12-2, 10-2, 5-2 y 2-2). No se hallaron diferencias estadísticamente significativas entre los grupos (p > .05).
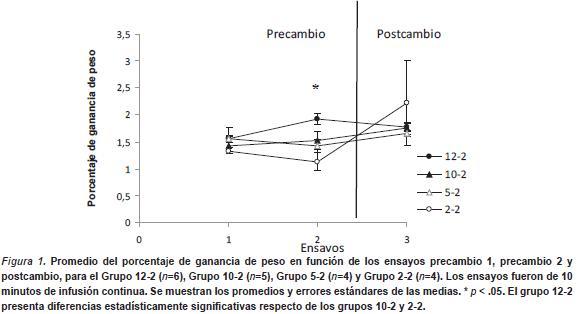
En resumen, este experimento muestra la existencia del EMRA, que aparece en el segundo ensayo de adquisición. También indica que los animales prefieren la solución al 12% y la diferencian entre las otras concentraciones. La falta de diferencias entre grupos durante el postcambio indica la ausencia de CSNc en ratas infantes de 8-10 DPN, al menos bajo los parámetros utilizados. Este hecho no puede atribuirse a una incapacidad para discriminar las diferentes concentraciones ya que se constató un consumo diferencial en función de las concentraciones de azúcar. Estos resultados sugieren que en esta edad los animales responden en función del valor absoluto del incentivo.
Experimento 2. CSNc - DPN 10 a DPN 14
La ausencia del CSNc en el experimento anterior puede deberse a una escasa cantidad de ensayos en la fase de precambio o que su expresión emerja unos días más tarde en el desarrollo ontogenético. Por estas razones, en el Experimento 2 se extendió la fase de precambio y se utilizaron animales de mayor edad. Además se usaron las dos concentraciones de azúcar que mostraron diferencias más acentuadas entre sí en el Experimento 1, 12% y 2%.
Método
Sujetos e instrumentos. Se utilizaron 28 ratas hembras Wistar de 10 DPN al comienzo del experimento, representativas de 7 camadas, criadas en las condiciones descritas para el Experimento 1. Los instrumentos utilizados fueron los mismos que en el Experimento 1.
Procedimiento. Fue esencialmente el mismo que en el Experimento 1, con excepción que se emplearon 2 grupos: Grupo 12-2 (n=14) y Grupo 2-2 (n=13) y la fase de precambio constó de 4 ensayos, a razón de uno por día. La fase de postcambio se realizó 24 hs. después del último ensayo de la fase de precambio en el cual los dos grupos recibieron 2% de solución de sacarosa.
Resultados y discusión.
En la Figura 2 se muestra el porcentaje de ganancia de peso en función de los ensayos de pre y postcambio. Durante la fase de precambio los animales parecen incrementar su consumo de manera gradual a lo largo de las sesiones, siendo el consumo aparentemente mayor en los animales que reciben 12% hacia el final de la fase de precambio. No parece observarse un efecto de contraste negativo. Estas impresiones fueron confirmadas por un ANOVA, el cual estuvo compuesto de un factor intersujeto (Grupo) y un factor intrasujeto (Ensayos 1 a 4) en la fase de precambio. En la fase de precambio se halló un efecto principal de Grupo F (1, 19) = 4.45, p < .04, y de Sesión F (3, 57) = 6.71, p < .0005. No se halló un efecto de interacción entre ambos factores (p > .05).
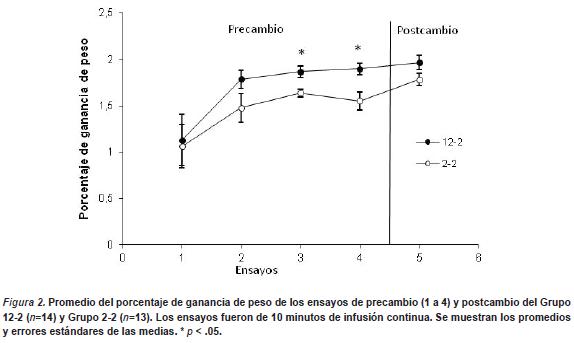
En la fase de postcambio, un ANOVA de un factor intersujeto (Grupo 12-2 vs. Grupo 2-2) no arrojó diferencias estadísticamente significativas entre los grupos (p > .05). Este resultado replica el EMRA en la fase de precambio y muestra un efecto de sesión, de lo que se infiere que los animales optimizan el consumo en función de los ensayos, la falta de diferencias entre los grupos durante la fase de postcambio, indica ausencia de CSNc. Al igual que en el Experimento 1, el grupo experimental no mostró un decremento del consumo durante el ensayo de devaluación del reforzador.
Experimento 3. EMRA y EMRE - DPN 7 a DPN 12
Stanton y Amsel (1980) hallaron la expresión de EMRA a los 11 DPN y Brudette, Brake, Chen y Amsel (1976) hallaron el EMRE a los 18 DPN, con un procedimiento instrumental. El siguiente experimento se realizó con el objetivo de explorar si los animales de 7 y 8 DPN son capaces de discriminar entre reforzadores de distinto valor (EMRAc) y si se expresa el EMREc a la edad de 11 y 12 DPN. Para evaluar el EMREc los animales recibieron agua durante la extinción.
Método
Sujeto e instrumentos. Se utilizaron 31 ratas hembras Wistar de 7 DPN al comienzo del experimento, representativas de 9 camadas. Las demás condiciones y los instrumentos utilizados se mantuvieron iguales al Experimento 1.
Procedimiento. Los sujetos se dividieron en dos grupos en función del valor del incentivo recibido en la fase de adquisición: 12% (n=15, Grupo 12-0) y 2% (n=16, Grupo 2-0). La fase de adquisición constó de 4 ensayos, uno por día, en los cuales los animales recibieron soluciones de sacarosa de 12% ó 2%, de acuerdo al grupo al que habían sido asignados. La fase de extinción constó de 2 ensayos diarios de 10 min y se realizó 24 hs. después del último ensayo de adquisición. Durante la extinción los animales recibieron una solución neutra (i.e., agua), debido a que al tratarse de un procedimiento consumatorio no sería posible retirar completamente el acceso a un fluido y aun así contar con una medida dependiente. Estudios no publicados de nuestro laboratorio mostraron que la presencia o no de agua en un bebedero no alteraba los patrones de extinción, cuando se utilizaba tiempo de contacto con el bebedero como medida dependiente. Los restantes detalles de procedimiento son iguales que en el Experimento 1.
Resultados y discusión.
En la Figura 3 se muestra el porcentaje de ganancia de peso en función de los ensayos de las fases de adquisición y extinción. Se observa que durante la fase de adquisición los animales consumen más solución a lo largo de los ensayos, observándose nuevamente mayor consumo en aquellos que reciben 12% (i.e., EMRA). Durante la extinción se observa una caída abrupta del porcentaje de ganancia de peso en los dos grupos durante el primer ensayo y un aumento en el segundo ensayo de extinción. El ANOVA de la fase de adquisición arroja un efecto principal de Grupo F (1, 27) = 11,67, p < .002, y de Sesión F (3, 81) = 18,67, p < .0001. No se halló un efecto de interacción entre ambos factores (p > .05). El ANOVA de la fase de extinción arroja un efecto significativo principal de Sesión, F (1, 29) = 15,97, p < .0005 y la interacción entre ambos factores es también significativa, F (1, 29) = 4,94, p < .03. Un análisis de cada sesión por separado no mostró diferencia significativa en la primera sesión, pero sí se observó una diferencia marginalmente significativa en la sesión 2 (p < .055). El grupo 2% exhibió mayor consumo de la solución neutra que el grupo 12% en la segunda sesión.
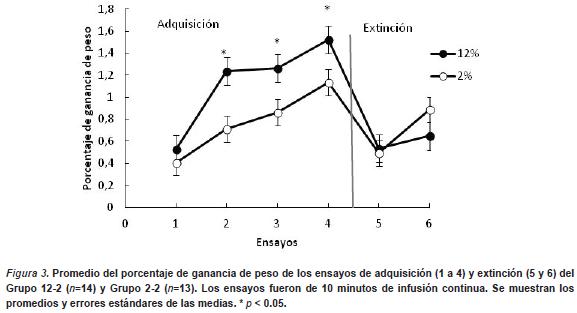
Experimento 4. CA - DPN 10 a DPN 16
En este experimento se evaluó si las ratas de 10 - 16 DPN discriminan entre soluciones azucaradas de diferentes concentraciones y si son capaces de anticipar la presentación de un reforzador de magnitud más elevada y, por lo tanto, suprimir el consumo de una solución menos preferida que la antecede inmediatamente (i.e., CA).
Método
Sujeto e instrumentos. Se utilizaron 27 ratas hembras Wistar de 10 DPN al comienzo del experimento, representativas de 8 camadas. Las demás condiciones y los instrumentos utilizados se mantuvieron iguales que en el Experimento 1.
Procedimiento. Los sujetos se dividieron en dos grupos en función de la concentración de sacarosa presente en la solución 2: 12% (n=14, Grupo 2-12) o 2% (n=13, Grupo 2-2). El procedimiento constó de 7 sesiones de 2 ensayos cada una. En el Ensayo 1, todos los animales recibieron una infusión continua de 5 min de solución de sacarosa al 2%. En el Ensayo 2 de cada sesión, los animales fueron infundidos de forma continua con soluciones de sacarosa al 12% ó 2%, de acuerdo al grupo al que pertenecían. El intervalo entre ensayos era de aproximadamente 1 min. Los restantes detalles de procedimiento fueron iguales que en el Experimento 1.
Resultados y discusión
En la Figura 4.a. se muestra el porcentaje de ganancia de peso en función de los ensayos de la primera solución. Se observa que ambos grupos presentan en todos los ensayos un porcentaje de ganancia de peso similar y que se incrementa a lo largo de los ensayos. Un ANOVA de dos factores (Grupo x Sesión) arroja un efecto principal de Sesión, F (6, 144) = 14,45, p < .0001, pero no un efecto de Grupo ni la interacción entre ambos factores (p > .05).
En la Figura 4.b. se muestra el porcentaje de ganancia de peso durante los segundos ensayos de cada sesión. Se observa que el Grupo 2-12 muestra un mayor porcentaje de ganancia de peso, respecto del Grupo 2-2, en todas las sesiones. El ANOVA de la solución 2 arroja un efecto significativo principal de Grupo, F (1, 23) = 67,62, p < .0001, Sesión, F (6, 138) = 20,83, p < .0001 y la interacción entre ambos factores es también significativa, F (6, 138) = 3,06, p < .007. Un análisis de cada sesión por separado mostró diferencias estadísticamente significativas entre los grupos en todas las sesiones (p < .0001).
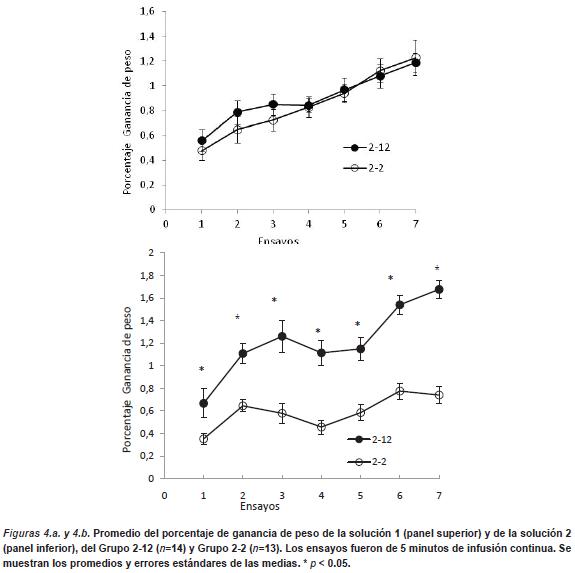
Estos resultados indican la ausencia de contraste anticipatorio, ya que el consumo de la primera solución no se modifica a lo largo de los ensayos, aún cuando es seguida inmediatamente por una solución más preferida. Sin embargo, nuevamente se observó que los animales eran capaces de discriminar entre diferentes concentraciones de la solución azucarada y mostrar prefierencias por la solución de más concentración de azucar (12%).
Discusión general
En conjunto, los resultados de estos experimentos indican que los animales de 8 DPN, 9 DPN, 10 DPN y 12 DPN manifiestan el EMRAc, ya que son capaces de discriminar y responder de manera diferencial en función de distintas concentraciones de agua azucarada, prefriendo la solución al 12%. Además se infiere que aprenden una respuesta consumatoria, ya que la ganancia de peso va aumentando en función de los ensayos. Según el Experimento 1 logran discriminar entre soluciones al 12% de las del 10%, 5% y 2% de sacarosa. En estas edades los animales requirieron de experiencia previa con el reforzador para evidenciar EMRAc, ya que este fenómeno fue más marcado a partir del segundo ensayo de adquisición. Es posible también que los animales necesiten habituarse al procedimiento de infusión intrabucal, el cual puede tener elementos aversivos propios (véase Pautassi et al., 2005).
De los que conocemos, ésta es la primera evidencia de EMRA consumatorio en ratas infantes con distintas concentraciones de soluciones azucaradas y con diferentes paradigmas. Stanton y Amsel (1980) encontraron EMRA instrumental a los 11 DPN. Sin embargo los resultados del presente estudio evidencian que tal efecto aparece ya a los 8 DPN con el paradigma consumatorio.
Nuestra hipótesis de encontrar efectos paradójicos consumatorios a etapas más tempranas que en los paradigmas instrumentales no fue confirmada, al menos en estas edades y procedimientos. En los Experimentos 1 y 2 no se halló CSNc (DPN 10 y DPN 14 respectivamente) y los Experimento 3 y 4 tampoco reflejan la aparición del EMREc y del CA (DPN 11 - 12 y DPN 10 - 16 respectivamente). Estos resultados pueden deberse a razones paramétricas de la metodología utilizada, o bien a que en las edades estudiadas los animales evalúan los incentivos en función de su valor absoluto y no es aún un factor relevante en su conducta el valor relativo de los reforzadores. Respecto de la primera conjetura, los EPR bajo análisis pueden requerir más ensayos de entrenamiento, menor intervalo entre ensayos o el uso de otros reforzadores. Sin embargo, la ausencia de estos fenómenos no podría deberse a una incapacidad para detectar las diferentes concentraciones de sacarosa, tal como queda evidenciado por el efecto de EMRAc en todos los experimentos presentados.
Es interesante mencionar que en el Experimento 3 los animales a los que se les devaluó el refuerzo exhibieron, durante el segundo ensayo de extinción, un consumo marginalmente menor que los sujetos que no habían experimentado devaluación del reforzador. Esto podría constituir un esbozo de EMREc, si bien la ausencia de diferencias significativas entre los grupos, la caída abrupta en ambos grupos en la primera sesión de extinción y la imposibilidad de realizar una extinción total (i.e., sin la presentación de una solución neutra durante la fase de devaluación) impide aventurar conclusiones más firmes. Más en detalle, en el Experimento 3 los animales muestran una disminución abrupta del consumo cuando reciben agua en la primera sesión y en la siguiente ambos grupos consumen más de la misma solución con una tendencia mayor en el grupo 2%. Este patrón de respuesta no es representativo de una extinción. Más bien sugiere que el agua es un incentivo menos preferido para estos infantes y que el primer día de devaluación sufrieron un efecto de contraste negativo que se recuperó en el segundo ensayo. Esta hipótesis es posible, ya que el protocolo incluye 3 horas de privación materna y los infantes pueden estar motivados por el agua. Estas especulaciones deberán ponerse a prueba realizando nuevos experimentos en los cuales al menos se agregue un grupo que reciba agua durante las dos fases del entrenamiento.
Otra explicación para los resultados hallados es que en las edades estudiadas los animales no respondan en función del valor relativo de los reforzadores (Flaherty, 1996). Esto podría deberse a que los factores neurales que modulan las respuestas de los ERPs de los animales en edades más avanzadas, como el hipocampo y la amígdala, no alcanzaron la maduración suficiente en las edades estudiadas. De todos modos es de destacar que estos son los primeros trabajos de EPR consumatorios, por lo cual antes de concluir ausencia del EMREc, CSNc y CA en las edades presentadas se debe realizar variaciones en la metodología utilizada.
Agradecimientos
Este trabajo fue parcialmente financiado por el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), la Agencia Nacional de Promoción Científica y Tecnológica, UBACYT-UBA y la Universidad Abierta Interamericana (UAI). Agradecemos al Dr. Juan Carlos Molina por poner a nuestra disposición los aparatos, insumos y asesoramiento para iniciar esta investigación.
Referencias
Amsel, A. (1992). Frustration Theory. Cambridge, UK: Cambridge University Press. Appleton. Traducción al castellano en Madrid: Alianza, 1984. [ Links ]
Amsel, A. & Stanton, M. (1980). Ontogeny and Phylogeny of Paradoxical Reward Effects. Advances in the Study of Behavior, 11, 227-267. [ Links ]
Amsel, A., Letz, R., & Burdette, D.R. (1977). Appetitive Learning and Extinction in 11-Day-Old Rat Pups: Effects of Various Reinforcement Conditions. Journal of Comparative and Physiological Psychology, 81, 1166-1167. [ Links ]
Arias, C. & Chotro, M. G. (2005). Increased palatability of ethanol after prenatal ethanol exposure is mediated by the opioid system. Pharmacology Biochemistry and Behavior, 82, 434-442. [ Links ]
Arias, C., Pautassi, R. M., Molina, J. C., & Spear, N. E. (2010). A Comparison Between Taste Avoidance and Conditioned Disgust Reactions Induced by Ethanol and Lithium Chloride in Preweanling Rats. Developmental Psychobiology, 52, 545-557 [ Links ]
Berdel, B., Morys, J., & Maciejewska, B. (1997). Neuronal changes in the basolateral complex during development of the amygdala of the rat. International Journal of Developmental Neuroscience, 15, 755-765. [ Links ]
Brake, S., Burdette, D.R., Chen, J., & Amsel, A. (1980). Retention of Response Persistence in Weanling and Adolescent Rats. Journal of Comparative and Physiological Psychology, 94, 1060-1068. [ Links ]
Burdette, D.R., Brake, S., Chen, J., & Amsel, A. (1976). Ontogeny of persistence: Immediate extinction effects in preweanling and weanling rats. Animal Learning & Behavior, 4, 131-138. [ Links ]
Chen, J., Gross, K., & Amsel, A. (1981). Ontogeny of Successive Negative Contrast and Its Dissociation From Other Paradoxical Reward Effects in Preweanling Rats. Journal of Comparative and Physiological Psychology, 95, 146-159. [ Links ]
Chen, J., Gross, K., Stanton, M., & Amsel, A. (1980). The partial reinforcement acquisition effect in preweanling and juvenile rats. Bulletin of the Psychonomic Society, 16, 239-242. [ Links ]
Chen, J., Gross, K., Stanton, M., & Amsel, A. (1981). Adjustment of weanling and adolescent rats to a reward condition requiring slow responding. Developmental Psychobiology, 14, 139-45. [ Links ]
Chotro, M. G., & Alonso, G. (2003). Stimulus preexposure reduces generalization of conditioned taste aversions between alcohol and non-alcohol favors in infant rats. Behavioral Neuroscience, 117, 113-122. [ Links ]
Crook, C. K. (1978). Taste perception in the newborn infant. Infant Behavior and Development, 1, 52-69. [ Links ]
Crook, C. K., & Lipsitt, L. P. (1976). Neonatal nutritive sucking: effects of taste stimulation upon sucking rhythm and heart. Child Development, 47, 518-52. [ Links ]
Desor, J. A., Maller, O., & Turner, R. E. (1976). Taste in acceptance of sugars by human infants. Journal of Comparative and Physiological Psychology, 84, 496-501. [ Links ]
Domjan, M. (1998). Principios de aprendizaje y conducta. Cuarta edición. D. F., México: International Thomson Editores. [ Links ]
Engen, T., & Lipsitt, L. P. (1974). Ability of Newborn Infants to Discriminate Sapid Substances. Developmental Psychology, 10, 741-744. [ Links ]
Flaherty, C. F. (1996). Incentive relativity. Cambridge, UK: Cambridge University Press. [ Links ]
Flaherty, C. F., Turovsky, J., & Krauss, K. L. (1994). Realtive hedonic value modulates anticipatory contrast. Physiology & Behavior, 55, 1047-1054. [ Links ]
Ganchrow, J. R., Steiner, J. E., & Daher, M. (1983). Neonatal facial expressions in response to different qualities and intensities of gustatory stimuli. Infant Behavior and Development, 6, 473-484. [ Links ]
Garcia, L., Garcia, C.H.S., Calábria, L.K., Da Cruz, G.C.N., Puentes, A.S., Bao, S.N., Fontes, W., Ricart, C.A.O., Espindola, F.S., & De Sousa, M.V., (2009). Proteomic analysis of honey bee brain upon ontogenetic and behavioral development. Journal of Pro-teome Research, 8, 1464-1473. [ Links ]
Gemberling, G. A., & Domjan, M. (1982). Selective associations in one-day-old rats: taste-toxicosis and texture-shock aversion learning. Journal of Comparative Physiological and Psychology, 96, 105-113. [ Links ]
Gemberling G. A., Domjan, M., & Amsel, A. (1980). Aversion learning in 5-day-old rats: taste-toxicosis and texture-shock associations. Journal of Comparative Physiological and Psychology, 94, 734-745. [ Links ]
Gray, J. A. (1987). The Psychology of fear and stress. Cambridge, UK: Cambridge University Press. [ Links ]
Gregg, B. Kittrell, M. W. Domjan, M., & Amsel, A. (1978). Ingestional Aversion Learning in Preweanling Rats. Journal of Comparative and Physiological Psychology, 92, 785-795. [ Links ]
Hall, W. G., & Rosenblatt, J. S. (1977). Suckling Behavior and Intake Control in the Developing Rat Pup. Journal of Comparative aanndd PPhysiological Psychology, 91, 1232-1247. [ Links ]
Hoffmann, H., Hunt, P., & Spear, N. E. (1991). Ontogenetic Differences in CS Palatability following Conditioned Taste Aversion Training. Learning and Motivation, 22, 329-352. [ Links ]
Kamenetzky, G. V., & Mustaca, A. E. (2005). Modelos animales para el estudio del alcoholismo. Terapia Psicológica, 23, 65-72. [ Links ]
Kobre, K.R., & Lipsitt, L.P. (1972). A Negative Contrast Effect in Newborns. Journal of Experimental Child Psychology, 14, 81-91 [ Links ]
Letz, R., Burdette, D.R., Gregg, B., Kittrell, E.M., & Amsel, A. (1978). Evidence for a Transitional Period for Development of Persistence in Infant Rats. Journal of Comparative and Physiological Psychology, 92, 856-866. [ Links ]
Mast, V.K., Fagen, J.W., Rovee-Collier, C.K., & Sullivan, M.W. (1980). Immediate and longterm memory for reinforcement context: The development of learned expectancies in early infancy. Child Development, 51, 700-707. [ Links ]
Mickley, G. A., Remmers-Roeber, D. R., Crouse, C., & Peluso, R. (2000). Ketamine blocks a taste-mediated conditioned motor response in perinatal rats. Pharmacology, Biochemestry and Behavior, 66, 547-552. [ Links ]
Miller, H., & Spear, N. E. (1989). Age-Dependent Facilitation of Taste-Footshock Conditioning by Prior Exposure to the Training Context. Behavioral and Neural Biology, 52, 123-130. [ Links ]
Mustaca, A., Freidin, E. & Papini, M. (2002). Extinction of Consummatory Behavior in Rats. International Journal of Comparative Psychology, 15, 1-10. [ Links ]
Mustaca, A., Bentosela, M., Pellegrini, S., Ruetti, E., Kamenetzky, G y Cuello, M. (2005). Aportes para la comprensión de la frustración. En: "Las Ciencias del Comportamiento en los albores del Siglo XXI". Compilación: Jorge Vivas. Editorial Universitaria. Bs. As. [ Links ]
Mustaca, A. E., Bentosela, M., Ruetti, E., Kamenetzky, G., Cuenya, L., Justel, N. Lopez Seal, F., Fosacheca, S., & Papini, M. (2009). Similitudes y discrepancias en dos modelos animales de frustración. En: Recientes desarrollos iberoamericanos en investigación en Ciencias del Comportamiento. Ed. CIIPME-CONICET. Libro editado por el 20 aniversario de la Asociación Argentina de Ciencias del Comportamiento. Bs.As. Argentina. Tomo II, 921-940. [ Links ]
Nizhnikov, M. E., Petrov, E. S., Varlinskaya, E. I., & Spear, N. E. (2002). Newborn rats' first suckling experience: Taste differentiation and suckling plasticity. Physiology & Behavior, 76, 181-198. [ Links ]
Nowlis, G. H., & Kessen, W. (1976). Human newborns differentiate differing concentrations of sucrose and glucose. Science, 191, 865-866. [ Links ]
Pautassi, R.M., Arias, C., Molina J. C., & Spear, N.E. (2008) Domperidone interferes with conditioned disgust reactions but not taste avoidance evoked by a LiCl paired taste in infant rats. Developmental Psychobiology, 50, 343-352. [ Links ]
Pautassi R. M., Melloni C., Ponce L. F., & Molina J. C. (2005). Acute ethanol counteracts the acquisition of an aversive olfactory learning in infant rats. Alcohol: an International Biomedical Journal, 36, 2, 99-105. [ Links ]
Pautassi, R. M., Molina J. C., & Spear, N. E. (2008). Infant rats exhibit aversive learning mediated by ethanol's orosensory effects but are positively reinforced by ethanol's postingestive effects. Pharmacology, Biochemistry, and Behavior, 88, 393-402. [ Links ]
Pautassi, R.M., Nizhnikov, & Spear, N.E. (2009) Assessing appetitive, aversive, and negative ethanolmediated reinforcement through an immature rat model. Neuroscience and Biobehavioral Reviews, 33, 953-974. [ Links ]
Salinas, J.A., & White, N.M. (1998). Contributions of the Hippocampus, Amygdala, and Dorsal Striatum to the Response Elicited by Reward Reduction. Behavioral Neuroscience, 112, 812-826. [ Links ]
Sastre, A., & Reilly, S. (2006). Excitotoxic lesions of the gustatory thalamus eliminate consummatory but not instrumental successive negative contrast in rats. Behavioural Brain Research, 170, 34-40. [ Links ]
Spear, L. P., Specht, S. M., Kirstein, C. L., & Kuhn, C. M. (1989). Anterior and posterior, but not cheek, intraoral cannulation procedures elevate serum corticosterone levels in neonatal rat pups. Developmental Psychobiology, 22, 401-411. [ Links ]
Stanton, M., & Amsel, A. (1980). Adjustment to Reward Reduction (but No Negative Contrast) in Rats 11, 14, and 16 Days of Age. Journal of Comparative and Physiological Psychology, 94, 446-458. [ Links ]
Stanton, M., Dailey, W., & Amsel, A. (1980). Patterned (Single) Alternation in 11-and 14-Day-Old Rats Under Various Reward Conditions. Journal of Comparative and Physiological Psychology, 94, 469-471. [ Links ]
Tinbergen N. (1963). En Aims and Methods in Ethology. Zeitschrift für Tierpsychologie, 1963, 20: 410-433. [ Links ]
