










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Psicología
versión impresa ISSN 0121-5469
Rev. colomb. psicol. v.20 n.2 Bogotá jul./dic. 2011
Modulación Contextual en la Extinción: Recuperación de una Aversión Condicionada al Sabor en un Tercer Contexto*
Contextual Modulation of Extinction: Recovery of Taste-Aversion Conditioning in a Third Context
Modulação Contextual na Extinção: Recuperação de uma Aversão Condicionada ao Sabor em um Terceiro Contexto
RODOLFO BERNAL-GAMBOA
RODRIGO CARRANZA-JASSO
GABRIELA GONZÁLEZ-MARTÍN
YECTIVANI JUÁREZ
JAVIER NIETO
LIVIA SÁNCHEZ-CARRASCO
Universidad Nacional Autónoma de México, México D. F.
La correspondencia en relación con este artículo puede dirigirse a Javier Nieto, e-mail: janigu@unam.mx.
División de Investigación y Posgrado, Facultad de Psicología, Universidad Nacional Autónoma de México,
Av. Universidad 3004, Col. Copilco-Universidad, Coyoacán, C. P. 04510, México, D. F.
ARTÍCULO DE INVESTIGACIÓN CIENTÍFICA
RECIBIDO: 15 de junio del 2010 - ACEPTADO: 22 de julio del 2011
Resumen
Este experimento se diseñó para evaluar la renovación ABC con un procedimiento de condicionamiento de aversión al sabor (CAS) en tres fases: adquisición, extinción y prueba. Durante la adquisición, 15 ratas Wistar, asignadas a los grupos ABC y ABB, tuvieron acceso a una solución con sacarosa y posteriormente recibieron una inyección de cloruro de litio (LiCl) en el Contexto A. En la extinción, ambos grupos bebieron la solución con sacarosa sin la administración del LiCl en el Contexto B. En la prueba, los grupos tuvieron acceso a la solución con sacarosa sin administrarles LiCl en el contexto de extinción para el grupo ABB, y en un tercer contexto para el grupo ABC. Los resultados mostraron la recuperación de la aversión condicionada al sabor únicamente en el grupo ABC.
Palabras clave: renovación, extinción, condicionamiento de aversión al sabor, ratas.
Abstract
This experiment was designed to evaluate ABC renewal in a conditioned taste aversion (CTA) procedure consisting of three phases: acquisition, extinction and test. During the acquisition phase, 15 Wistar rats assigned to groups ABC and ABB had access to a sucrose solution, after which they were injected with lithium chloride (LiCl) in Context A. In the extinction phase, both groups of rats had access to the sucrose solution in Context B, but no LiCl was administered. Finally, during the test, group ABB received sucrose solution without LiCl in the extinction context, while group ABC was tested in a third context. Results showed renewal of conditioned taste aversion only in the ABC group.
Keywords: renewal, extinction, conditioned taste aversion, rats.
Resumo
Este experimento foi elaborado para avaliar a renovação ABC com um procedimento de condicionamento de aversão ao sabor (CAS) em três fases: aquisição, extinção e prova. Durante a aquisição, 15 ratos Wistar, designados aos grupos ABC e ABB, tiveram acesso a uma solução com sacarose e posteriormente receberam uma injeção de cloreto de lítio (LiCl) em Contexto A. Na extinção, ambos grupos beberam a solução com sacarose sem a administração do LiCl em Contexto B. Na prova, os grupos tiveram acesso à solução com sacarose sem administrar-lhes LiCl num contexto de extinção para o grupo ABB, e num terceiro contexto para o grupo ABC. Os resultados mostraram a recuperação da aversão condicionada ao sabor unicamente no grupo ABC.
Palavras-chave: renovação, extinção, condicionamento de aversão ao sabor, ratos.
ACTUALMENTE, SE acepta que el decremento en la respuesta condicionada (RC) observado durante la extinción no significa la destrucción de las asociaciones entre el estímulo condicionado (EC) y el estímulo incondicionado (EI) que se establecieron en la adquisición (Delamater, 2004; Rescorla, 2001). Un ejemplo de ello, es el fenómeno de renovación contextual, que muestra la reaparición de la RC extinguida como consecuencia de un cambio en el contexto experimental (Bouton, Westbrook, Corcoran & Maren, 2006). Por ejemplo, cuando se empareja un EC con un EI o se refuerza una respuesta instrumental en un contexto particular (denominado A), y luego se extingue a través de la omisión del EI o del reforzamiento instrumental en un segundo contexto (denominado B), regresar a los sujetos al contexto original (A) durante la fase de prueba produce la reaparición de la RC o la respuesta instrumental. Específicamente, este tipo de renovación contextual se conoce como renovación ABA (e. g. Bouton & Bolles, 1979; Nakajima, Tanaka, Urushihara & Imada, 2000). De la misma forma, se ha reportado la recuperación de la respuesta cuando la adquisición y la extinción se conducen en el mismo contexto, y la fase de prueba se realiza en un contexto diferente (i. e. renovación AAB) (e. g. Bouton & Ricker, 1994; Rescorla, 2008), así como, cuando la adquisición, la extinción y la prueba se conducen en tres contextos distintos (i. e. renovación ABC)(e. g. Bouton & Swartzentruber, 1986; Thomas, Larsen & Ayres, 2003).
En la década de los 90s, Bouton (1993, 1994, 1997) desarrolló uno de los modelos asociativos más influyentes dentro de esta área con el propósito de explicar la renovación contextual, así como otros fenómenos en los que se observa la recuperación de respuestas extinguidas, como son la recuperación espontánea, el restablecimiento, entre otros. Este modelo se conoce como modelo de recuperación de la información y asume que la memoria está formada por nodos o unidades que representan los eventos del mundo, así como las asociaciones que se establecen entre ellos. De acuerdo con dicho modelo, durante la fase de adquisición pavloviana se forma una asociación excitatoria entre la representación del EC y del EI. Posteriormente, en la fase de extinción, esta asociación no se rompe ni se destruye; por el contrario, se genera una nueva asociación entre la representación del EC y del EI, cuyo carácter es inhibitorio (Konorski, 1948). Asimismo, el modelo supone que durante la extinción los organismos representan el contexto y esta representación es capaz de modular la activación de la asociación inhibitoria a través de una compuerta lógica AND. Así, cuando las representaciones del EC y del contexto se activan simultáneamente, la asociación inhibitoria atenúa la activación del nodo del EI observándose una ejecución similar a la de extinción. Por otro lado, si la representación del contexto no se activa simultáneamente con la representación del EC, se activará la asociación excitatoria y se observará la renovación contextual, es decir, la ejecución será similar a la de la fase de adquisición.
Una de las principales predicciones del modelo de recuperación de información es que la RC extinguida se observará siempre que se presente el EC en un contexto distinto al utilizado en la extinción. Así, los tres diseños de renovación contextual ABA, AAB y ABC son manipulaciones equivalentes en cuanto a su potencial para producir renovación, y aun más se considera que el mecanismo subyacente a los tres tipos de renovación es el mismo. Se han publicado varios reportes de renovación ABA, AAB y ABC consistentes con esta predicción, a través de diferentes arreglos experimentales como supresión condicionada (Bouton & King, 1983; Harris, Jones, Bailey & Westbrook, 2000; Tamai & Nakajima, 2000), condicionamiento pavloviano apetitivo (Bouton & Peck, 1989; Rescorla, 2008), condicionamiento instrumental (Nakajima, Urushihara & Masaki, 2002; Sánchez-Carrasco, Bernal-Gamboa & Nieto, 2011) y en tareas de juicios causales con humanos (Pineño & Miller, 2004; Rosas & Callejas-Aguilera, 2006; Vila & Rosas, 2001); mientras que con el condicionamiento de aversión al sabor (CAS) solo se ha reportado con los diseños de renovación ABA y AAB (Rosas & Bouton, 1997; Rosas, García-Gutiérrez & Callejas-Aguilera, 2007).
Cuando se realizan experimentos de renovación contextual empleando CAS, es particularmente importante tener presente las observaciones metodológicas señaladas por Rosas y Bouton (1997) a investigaciones realizadas por Archer y colaboradores: primero, los resultados de Archer, Sjödén, Nilsson y Carter (1979) (ver también Archer, Sjödén, Nilsson & Carter, 1980) muestran un efecto similar a la renovación ABA; sin embargo, el cambio de contexto entre el condicionamiento (i. e. Contexto A) y la extinción (i. e. Contexto B) produjo un decremento en la RC, por lo que el hallazgo de renovación ABA puede atribuirse a la falta de extinción o al decremento en la generalización. Segundo, en el experimento descrito por Archer et al. (1980) se expuso a los sujetos a uno de los contextos cada día, iniciando con el Contexto A, así al iniciar la fase de extinción, el Contexto B resultaba novedoso, situación que favorece el establecimiento de asociaciones directas contexto-sabor.
Experimentos recientes que evalúan la renovación contextual en CAS han utilizado estrategias que eliminan las deficiencias metodológicas antes descritas; específicamente, exponen a los sujetos a todos los contextos a lo largo del experimento, de tal manera que se elimina su novedad en las sesiones de cambio contextual y se reducen las probabilidades de establecer asociaciones directas contexto-sabor.
Hasta donde los autores del presente trabajo conocen, a la fecha únicamente se han publicado dos trabajos de investigación sobre el CAS (Rosas & Bouton, 1997; Rosas et al., 2007) en los que se demostraron los diseños de renovación ABA y AAB, y se emplearon los controles metodológicos descritos. La demostración de renovación ABC es particularmente relevante porque, como se señaló previamente, el modelo de recuperación de la información predice renovación siempre que se retire a los sujetos del contexto de extinción. Así, en términos teóricos, la demostración de este tipo de renovación sugiere que la recuperación de la respuesta depende de la presentación del EC fuera del contexto de extinción, y no de las propiedades asociativas del contexto o del estímulo. Este resultado daría sustento a las predicciones realizadas por el modelo de recuperación de información.
En conclusión, el presente experimento tuvo como propósito demostrar la renovación ABC, eliminando los problemas metodológicos señalados por Rosas y Bouton (1997). Para ello, se entrenaron dos grupos de ratas en una preparación del CAS en la que se igualó la exposición y familiaridad de los contextos. A lo largo del experimento, se expuso a los sujetos de manera contrabalanceada a tres ensayos por día (mañana, tarde y noche), en los cuales consumieron agua o una solución de sacarosa. En la fase de adquisición, que se condujo en un contexto denominado A, se asoció el consumo de sacarosa con la administración de una inyección intraperitoneal (IP) de LiCl, posteriormente la fase de extinción se condujo en el Contexto B y se permitió a los sujetos consumir la sacarosa en ausencia del LiCl. Finalmente, en la fase de prueba los dos grupos tuvieron acceso a la solución con sacarosa en ausencia del LiCl. Se probó al grupo ABB en el Contexto B y al grupo ABC en el Contexto C.
Método
Sujetos
Se utilizaron 16 ratas macho, cepa Wistar, experimentalmente ingenuas, con un peso promedio de 293 g. Al inicio del experimento una de las ratas del grupo ABC enfermó y se eliminó del experimento.
Instrumentos
Se emplearon 12 cajas de Plexiglás, las cuales se acondicionaron para representar tres contextos experimentales distintos. Ocho de las cajas medían 22 x 20 x 43 cm, de las cuales cuatro tenían las paredes cubiertas con papel verde oscuro, el piso estaba cubierto con empaques de cartón para huevo, debajo de los cuales se colocó un trozo de algodón humedecido con 10 ml de vinagre blanco (Clemente Jacques, Sabormex S.A. de C. V., México). Las cuatro cajas restantes tenían las paredes cubiertas con papel cuadriculado en rojo y blanco, cada cuadro media 7 mm por lado; el piso estaba cubierto con una tabla de perfocel debajo de la cual se colocó un trozo de algodón humedecido con 10 ml de esencia de vainilla (McCormick & Company Inc., Maryland). Las cuatro cajas que representaron el tercer contexto medían 18 x 15.5 x 28 cm, tenían las paredes cubiertas con papel bond marcado con líneas diagonales blancas y negras de 8 mm de ancho; el piso se cubrió con etileno acetato de vinil blanco (foamy) debajo del cual se colocó un trozo de algodón humedecido con 10 ml de Windex (S.C. Johnson and Son, S.A. de C.V., México).
Las cajas se colocaron en cubículos separados, con condiciones de iluminación y temperatura similares. La asignación de los sujetos a los contextos y a los grupos se contrabalanceó de la siguiente forma: para dos de los sujetos del grupo ABC el Contexto A estuvo constituido por las cajas con papel verde en la mañana, el Contexto B por las cajas con papel de cuadros rojos en la tarde, y el Contexto C por las cajas con líneas diagonales en la noche. Para los siguientes dos sujetos, el Contexto A correspondió a las cajas con papel de cuadros rojos en la tarde, el Contexto B a las cajas con líneas diagonales en la noche, y el Contexto C a las cajas con papel verde en la mañana. Finalmente, para las últimas tres ratas, el Contexto A estuvo conformado por las cajas con las líneas diagonales en la noche, el Contexto B por las cajas con papel verde en la mañana, y el Contexto C por las cajas con papel de cuadros rojos en la tarde.
Por otra parte, para dos de los sujetos del grupo ABB, el Contexto A correspondió a las cajas con papel verde en la mañana, el Contexto B a las cajas con papel de cuadros rojos en la tarde, y el Contexto C a las cajas con líneas diagonales por la noche. Para las siguientes tres ratas, el Contexto A se constituyó por las cajas con papel de cuadros rojos en la tarde, el Contexto B por las cajas con las líneas diagonales en la noche, y el Contexto C estuvo constituido por las cajas con papel verde en la mañana. Finalmente, para las últimas tres ratas, el Contexto A estuvo conformado por las cajas con líneas diagonales en la noche, el Contexto B por las cajas con papel verde en la mañana, y el Contexto C se constituyó por las cajas con papel de cuadros rojos en la tarde.
Se utilizó como EC una solución al 15 % de sacarosa diluida en agua destilada. El EI fue una inyección IP al 0.3 Molar de LiCl, al 0.5 % del peso corporal. Los líquidos se administraron en probetas de plástico, graduadas, de 50 ml, y se les colocó un chupón de acero inoxidable.
Procedimiento
Las ratas recibieron acceso libre a comida y agua durante dos semanas antes de iniciar el experimento, y se mantuvieron en un ciclo de luz-oscuridad normal, en el que las luces del bioterio se apagaban a las 22:00 h y se encendían a las 06:00 h.
Antes de iniciar el experimento, se familiarizó durante cinco días a los sujetos con el procedimiento de acceso a líquidos y al contexto experimental. Se impidió a las ratas el acceso al agua 24 horas antes de comenzar el experimento y se mantuvieron en un programa de privación de agua excepto en tres periodos de 10 min cada uno, con un intervalo entre periodos de consumo de 8 h. El primer periodo se produjo a las 06:00 h, el segundo a las 14:00 h y el tercero tuvo lugar a las 22:00 h. El uso de estos periodos de consumo permitió manipular la exposición a los tres contextos, haciendo que los sujetos tuvieran una experiencia similar con cada uno de ellos. Las fases experimentales de interés iniciaron el sexto día.
Días 1-3: las ratas recibieron agua destilada en cada una de las tres sesiones diarias en sus cajas habitación. Al finalizar la última sesión del tercer día, se asignó a los sujetos a los grupos ABC y ABB igualando el consumo promedio de agua, en ml, registrado durante los tres horarios en los tres días previos, de la misma forma en que fue descrito por Rosas et al. (2007). Durante estos días el consumo promedio para el grupo ABC en las sesiones de la mañana, tarde y noche fue de 8.5, 7.9 y 6.5 ml respectivamente, mientras que para el grupo ABB fue de 9.0, 7.8 y 6.1 ml respectivamente.
Días 4-5: todas las ratas recibieron agua en las tres sesiones, a la vez que fueron expuestas a los tres contextos de acuerdo con el patrón de asignación descrito previamente. Durante estos días el consumo promedio para el grupo ABC en las sesiones de la mañana, tarde y noche fue de 6.5, 6.0 y 5.4 ml respectivamente, mientras que para el grupo ABB fue de 5.7, 5.1 y 5.9. ml respectivamente.
Una vez concluida la familiarización se inició con el experimento, que constó de tres fases (ver Tabla 1).
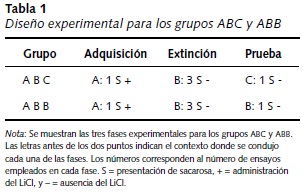
Día 6 (Adquisición): en esta fase los grupos ABB y ABC recibieron un ensayo de adquisición. Se dio acceso a los sujetos a la sacarosa en el Contexto A, durante 10 minutos; posteriormente se retiraron las probetas mientras las ratas permanecieron en dicho contexto. Diez minutos después, se retiraron de los contextos para inyectarles IP, la solución con LiCl, e inmediatamente después se regresaron al contexto por 10 minutos más, antes de colocarlos en sus cajas habitación. En las otras dos sesiones del día, que se condujeron en los Contextos B y C, se permitió a los sujetos beber agua destilada durante 10 minutos.
Día 7: todas las ratas recibieron agua destilada en sus cajas habitación en las tres sesiones del día.
Días 8-10 (Extinción): todos los sujetos recibieron tres ensayos de extinción; es decir, se les dio acceso a la solución de sacarosa durante 10 minutos en el Contexto B sin inyectarlas con LiCl, mientras que en las sesiones conducidas en los Contextos A y C se les permitió beber agua destilada por 10 minutos.
Día 11 (Prueba): se sometió a todas las ratas a un tratamiento idéntico al empleado en la fase anterior, de extinción, con la siguiente excepción: las ratas en el grupo ABC recibieron el acceso a la sacarosa en el Contexto C, mientras los sujetos del grupo ABB recibieron la solución de sacarosa en el mismo contexto en donde se realizó la extinción (i. e. Contexto B).
Resultados
Fase de Adquisición
En el panel izquierdo de la Figura 1 se muestra el consumo promedio (en ml) de sacarosa durante el único ensayo de adquisición. El consumo promedio fue de 7.6 y 7.9 ml para los grupos ABB y ABC, respectivamente. Una prueba t, confirmó la ausencia de diferencias significativas entre los grupos, t (13) = 0.15, p > .05.
Fase de Extinción
En el panel central de la Figura 1 se muestran los consumos promedio de sacarosa, en ml, durante cada uno de los tres ensayos de extinción. Como era de esperar, se observa un incremento paulatino en el consumo de sacarosa en ambos grupos conforme progresa la extinción. El primer ensayo de esta fase se utilizó como un indicador del condicionamiento de aversión al sabor, es decir, consumos bajos de sacarosa son indicativos de que se produjo la aversión condicionada al sabor. Un ANOVA de medidas repetidas que incluyó el factor Grupo (i. e. ABB y ABC) así como el factor Fase (i. e. ensayo de adquisición vs. primer ensayo de extinción), confirmó la adquisición de la aversión al sabor al resultar significativo para el factor Fase, F (1,13) = 32.085, p < .05, mientras el factor principal Grupo, F (1,13) = 0.002, p > .05, y la interacción, Grupo x Fase, F (1,13) = 0.092, p > .05, no resultaron significativos.
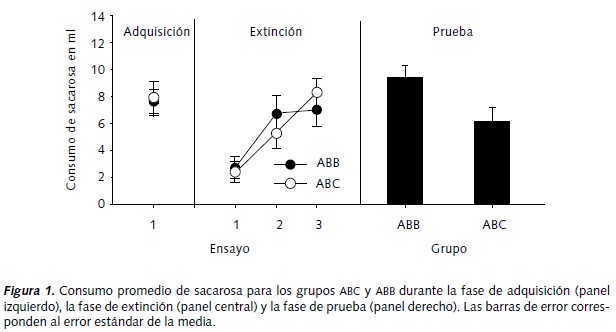
En el primer ensayo de la fase de extinción, el consumo promedio de sacarosa fue de 2.4 y 2.8 ml para los grupos ABB y ABC, respectivamente, e incrementó a 7.0 y 8.3 ml al finalizar el tercer ensayo. Un ANOVA mixto de dos factores Grupo x Ensayo indicó un efecto significativo del factor Ensayo, F (2, 26) = 20.800, p < .05, lo cual confirma que en el transcurso de los ensayos de extinción el consumo de sacarosa se incrementó. Ni el factor Grupo, F (1, 13) = 0.018, p > .05, ni la interacción Grupo x Ensayo, F (2, 26) = 1.491, p > .05, fueron significativas, indicando que ambos grupos mostraron niveles similares de consumo durante esta fase.
Fase de Prueba
El panel derecho de la Figura 1 muestra el consumo promedio de sacarosa en ml, para cada grupo durante el ensayo de prueba. Se puede observar que el consumo promedio de sacarosa presentado por el grupo ABC (6.1 ml) fue menor que el del grupo ABB (9.4 ml). Una prueba t confirmó la renovación de la aversión condicionada al sabor en un tercer contexto t (13) = -2.50, p < .05.
Discusión
El presente experimento mostró la reaparición de una respuesta de aversión al sabor cuando las fases de adquisición, extinción y prueba se condujeron en tres contextos distintos, pero igualmente familiares; es decir, se observó renovación ABC. Este resultado se suma a otras investigaciones que aportan evidencia de que la extinción no refleja el desaprendizaje o la ruptura de las asociaciones establecidas durante la adquisición (Bouton, 2004; Sánchez-Carrasco & Nieto, 2009). Asimismo, es consistente con hallazgos que muestran renovación ABC en una gran variedad de preparaciones con animales humanos y no humanos (Bouton & Swartzentruber, 1986; Harris, et al., 2000; Neumann, 2006; Pineño & Miller, 2004; Rescorla, 2008; Sánchez-Carrasco et al., 2011; Thomas et al., 2003). Por tanto, los resultados del presente experimento son consistentes con las predicciones del modelo de Bouton (1994), el cual sugiere que se observará la renovación de la respuesta cuando se retire a los sujetos del contexto de extinción, ya que únicamente las asociaciones establecidas durante esta fase muestran dependencia contextual.
Con respecto a las interpretaciones teóricas de estos resultados, es importante discutir algunas explicaciones alternativas al fenómeno de renovación. Por ejemplo, Pavlov (1927) reportó el efecto de desinhibición, en el cual se observa la recuperación de la RC extinguida como consecuencia de la presentación de un estímulo novedoso. Asimismo, supuso que las asociaciones inhibitorias son más lábiles que las asociaciones excitatorias y, por lo tanto, son más fáciles de alterar con la presencia de un estímulo nuevo. Sin embargo, los resultados reportados en la presente investigación no pueden explicarse como desinhibición, ya que durante todo el experimento los sujetos tuvieron una exposición equivalente a todos los contextos, evitando así que en el momento de la prueba el Contexto C fuese novedoso.
Por otro lado, existen investigaciones que muestran la participación de una asociación directa entre el contexto y el EI en el control de la aversión gustativa (Bonardi, Honey & Hall, 1990; López & Martínez, 2003; Loy, Álvarez, Rey & López, 1993), lo que puede dar cuenta de los resultados obtenidos. Sin embargo, estos resultados son inconsistentes con dicha propuesta, ya que se observó la renovación ABC cuando los sujetos tenían una experiencia equivalente con todos los contextos empleados. Asimismo, es importante considerar que el permitir que las ratas sedientas bebieran agua destilada en los contextos alternos, pudo favorecer que el contexto fuese asociado directamente con el consumo de agua; sin embargo, si el contexto hubiera actuado como otro EC y hubiera formado su propia asociación con el EI, se debería haber observado un mayor consumo del líquido en el grupo ABC durante la prueba en el Contexto C, ya que ese contexto estaría directamente asociado con el EI apetitivo (i. e. agua destilada), lo cual no se observó en los resultados encontrados.
Por lo tanto, se puede concluir que para observar el efecto de renovación ABC, las asociaciones contexto-EI no son necesarias o suficientes; por el contrario, los presentes resultados indican que el papel del contexto en este efecto es modular o señalar una asociación EC-EI (Loy & López, 1999; Nakajima, Kobayashi & Imada, 1995; Rosas et al., 2007), tal y como lo propone el modelo de recuperación de la información (Bouton, 1997; Bouton & Swartzentruber, 1986; Pearce & Bouton, 2001).
A pesar de que el hallazgo de renovación ABC en el CAS es consistente con las predicciones del modelo de Bouton, también existe evidencia que puede ser problemática para dicho modelo. Por ejemplo, Harris et al. (2000) demostraron un menor nivel de renovación ABC comparada con la renovación ABA utilizando una preparación de supresión condicionada. Recientemente, Rescorla (2008) ha reportado resultados similares en una preparación apetitiva con pichones. Incluso Havermans, Keuker, Lataster y Jansen (2005) reportan ausencia de renovación ABC cuando se empleó una tarea de supresión con humanos. Por lo anterior, los autores de este estudio consideran que para que se evalúe de forma más precisa el modelo de Bouton, hace falta realizar experimentos en los cuales se comparen directamente todos los diseños de renovación contextual, con lo cual se podría esclarecer si el mecanismo que subyace a los diseños ABA, AAB y ABC es el mismo o, como algunos autores lo han planteado, es distinto (ver Rosas, Callejas-Aguilera, Ramos-Álvarez & Abad, 2006).
Finalmente, es importante mencionar las implicaciones clínicas del estudio de la renovación, ya que aunque el condicionamiento clásico o pavloviano es un mecanismo de aprendizaje básico que puede encontrarse en una gran variedad de animales (e. g. Rescorla, 1988), también está relacionado con el desarrollo y el mantenimiento de un gran número de patologías humanas importantes (e. g. adicciones y fobias). Recientemente, varios autores han sugerido el CAS como un buen modelo para estudiar la ansiedad (Guitton & Dudai, 2004; Reilly & Schachtman, 2009), por ello no es de sorprender que los hallazgos relacionados con la renovación contextual sean considerados por varios terapeutas como un modelo para explicar las recaídas, sobre todo si se considera que el despacho del psicoterapeuta funciona como el Contexto B o el contexto de extinción (Glautier & Elgueta, 2009; Rauhut, Thomas & Ayres, 2001).
En relación con lo antes mencionado, los resultados reportados en la presente investigación sugieren dos puntos especialmente importantes: (a) es altamente probable que se observe una recaída aunque la intervención terapéutica haya "eliminado" en gran medida la conducta poco saludable, y (b) no es necesario que los sujetos regresen al lugar en donde sucedió el evento traumático para que haya una recaída o se observe de nuevo la conducta poco saludable (i. e. renovación ABC). Sin embargo, las manipulaciones experimentales en el presente estudio han mostrado que inclusive cuando la adquisición y la extinción se realizaron en contextos distintos, se puede evitar o atenuar la renovación de la respuesta si el contexto de prueba se iguala al contexto de extinción, justo como sucede en el diseño ABB (Bouton & Bolles, 1979; Harris et al., 2000; Tamai & Nakajima, 2000).
* Este proyecto se realizó con financiamiento otorgado por la DGAPA-PAPIIT 307509.
Referencias
Archer, T., Sjödén, P. O., Nilsson, L. G. & Carter, N. (1979). Role of exteroceptive background context in taste-aversion conditioning and extinction. Animal Learning & Behavior, 7, 17-22. [ Links ]
Archer, T., Sjödén, P. O., Nilsson, L. G. & Carter, N. (1980). Exteroceptive context in taste aversion conditioning and extinction: Odor, cage, and bottle stimuli. Quarterly Journal of Experimental Psychology, 32, 197-214. [ Links ]
Bonardi, C., Honey, R. C. & Hall, G. (1990). Context specificity of conditioning in flavor aversion learning: Extinction and blocking tests. Animal Learning & Behavior, 18, 229-237. [ Links ]
Bouton, M. E. (1993). Context, time, and memory retrieval in the interference paradigms of pavlovian learning. Psychological Bulletin, 114, 80-99. [ Links ]
Bouton, M. E. (1994). Context, ambiguity, and classical conditioning. Current directions in psychological science, 3, 49-53. [ Links ]
Bouton, M. E. (1997). Signals for whether versus when an event will occur. En M. E. Bouton & M. S. Fanselow (Eds.), Learning, motivation, and cognition: The functional behaviorism of Robert C. Bolles (pp. 385-409). Washington, DC: American Psychological Association. [ Links ]
Bouton, M. E. (2004). Context and behavioral processes in extinction. Learning & Memory, 11, 485-494. [ Links ]
Bouton, M. E. & Bolles, R. C. (1979). Contextual control of the extinction of conditioned fear. Learning & Motivation, 10, 445-466. [ Links ]
Bouton, M. E. & King, D. A. (1983). Contextual control of the extinction of conditioned fear: Tests for the associative value of the context. Journal of Experimental Psychology: Animal Behavior Processes, 9, 248-265. [ Links ]
Bouton, M. E. & Peck, C. A. (1989). Context effects on conditioning, extinction, and reinstatement in an appetitive conditioning preparation. Animal Learning and Behavior, 17, 188-198. [ Links ]
Bouton, M. E. & Ricker, S. T. (1994). Renewal of extinguished responding in a second context. Animal Learning and Behavior, 22, 317-324. [ Links ]
Bouton, M. E. & Swartzentruber, D. (1986). Analysis of the associative and occasion-setting properties of context participating in a pavlovian discrimination. Journal of Experimental Psychology: Animal Behavior Processes, 12, 333-350. [ Links ]
Bouton, M. E., Westbrook, R. F., Corcoran, K. A. & Maren, S. (2006). Contextual and temporal modulation of extinction: Behavioral and brain mechanisms. Biological Psychiatry, 60, 352-360. [ Links ]
Delamater, A. R. (2004). Experimental extinction in pavlovian conditioning: Behavioural and neu217 rosciences perspective. The Quarterly Journal of Experimental Psychology, 57 (2), 97-132. [ Links ]
Glautier, S. & Elgueta, T. (2009). Multiple cue extinction effects on recovery of responding in causal judgments. International Journal of Comparative Psychology, 22, 254-270. [ Links ]
Guitton, M. J. & Dudai, Y. (2004). Anxiety-like state associates with taste to produce conditioned taste aversion. Biological Psychiatry, 56, 901-904. [ Links ]
Harris, J. A., Jones, L., Bailey, G. K. & Westbrook, R. F. (2000). Contextual control over conditioned responding in an extinction paradigm. Journal of Experimental Psychology: Animal Behavior Processes, 26, 174-185. [ Links ]
Havermans, R. C., Keuker, J., Lataster, T., & Jansen, A. (2005). Contextual control of extinguished conditioned performance in humans. Learning & Motivation, 36, 1-19. [ Links ]
Konorski, J. (1948). Conditioned reflexes and neuron organization. London: Cambridge University Press. [ Links ]
López, M. & Martínez, A. (2003). Condicionamiento clásico de aversiones al contexto. En J. Vila, J. Nieto & J. M. Rosas (Eds.), Investigación contemporánea en aprendizaje asociativo (pp. 83-95). Jaén, España: Del Lunar-UNAM. [ Links ]
Loy, I., Álvarez, R., Rey, V. & López, M. (1993). Context- US associations rather than occasion setting in taste aversion learning. Learning & Motivation, 24, 55-72. [ Links ]
Loy, I. & López, M. (1999). Conditional control of toxicosis-based conditioning by context. Behavioural Processes, 46, 173-179. [ Links ]
Nakajima, S., Kobayashi, Y. & Imada, H. (1995). Contextual control of taste aversion in rats: The effects of context extinction. Psychological Record, 45, 309-318. [ Links ]
Nakajima, S., Tanaka, S., Urushihara, K. & Imada, H. (2000). Renewal of extinguished lever-press responses upon return to the training context. Learning & Motivation, 31, 416-431. [ Links ]
Nakajima, S., Urushihara, K. & Masaki, T. (2002). Renewal of operant performance formerly eliminated by omission or noncontingency training upon return to the acquisition context. Learning & Motivation, 33, 510-525. [ Links ]
Neumann, D. L. (2006). The effects of physical context changes and multiple extinction contexts on two forms of renewal in a conditioned suppression task with humans. Learning & Motivation, 37, 149-175. [ Links ]
Pavlov, I. P. (1927). Conditioned reflex. London: Oxford University Press [ Links ]
Pearce, J. M. & Bouton, M. E. (2001). Theories of associative learning in animals. Annual Review of Psychology, 52, 111-139. [ Links ]
Pineño, O. & Miller, R. R. (2004). Signaling a change in cue-outcome relations in human associative learning. Learning & Behavior, 32, 360-375. [ Links ]
Rauhut, A. S., Thomas, B. L. & Ayres, J. J. B. (2001). Treatments that weaken Pavlovian conditioned fear and thwart its renewal in rats: Implications for treating human phobias. Journal of Experimental Psychology: Animal Behavior Processes, 27, 99-114. [ Links ]
Reilly, S. & Schachtman, T. R. (2009). Conditioned taste aversion: Behavioral and neural processes. New York: Oxford University Press. [ Links ]
Rescorla, R. A. (1988). Pavlovian conditioning: It's not what you think it is. American Psychologist, 43, 151-160. [ Links ]
Rescorla, R. A. (2001). Experimental extinction. En R. R. Mowrer & S. Klein (Eds.), Handbook of contemporary learning theories (pp. 119-154), Hillsdale, N.J.: Erlbaum. [ Links ]
Rescorla, R. A. (2008). Within-subject renewal in sign tracking. The Quarterly Journal of Experimental Psychology, 61, 1793-1802. [ Links ]
Rosas, J. M. & Bouton, M. E. (1997). Renewal of a conditioned taste aversion upon return to the conditioning context after extinction in another one. Learning & Motivation, 28, 216-229. [ Links ]
Rosas, J. M. & Callejas-Aguilera, J. E. (2006). Context switch effects on acquisition and extinction in human predictive learning. Journal of Experimental Psychology: Learning, Memory, & Cognition, 32, 461-474. [ Links ]
Rosas, J. M., Callejas-Aguilera, J. E., Ramos-Álvarez, M. M. & Abad, M. J. F. (2006). Revision of retrieval theory of forgetting: What does make information context-specific? International Journal of Psychology and Psychological Therapy, 6, 147-166. [ Links ]
Rosas, J. M., García-Gutiérrez, A. & Callejas-Aguilera, J. E. (2007). AAB and ABA renewal as a function of the number of extinction trials in conditioned taste aversion. Psicológica, 28, 129-150. [ Links ]
Sánchez-Carrasco, L., Bernal-Gamboa, R. & Nieto, J. (2011). ABA, ABC, and AAB renewal in an Instrumental procedure. Manuscrito en preparación. [ Links ]
Sánchez-Carrasco, L. & Nieto, J. (2009). Recuperación de respuestas: una revisión de la evidencia y del modelo de recuperación de información. Revista Mexicana de Análisis de la Conducta, 35, 45-59. [ Links ]
Tamai, N. & Nakajima, S. (2000). Renewal of formerly conditioned fear in rats after extensive extinction training. International Journal of Comparative Psychology, 13, 137-147. [ Links ]
Thomas, B. L., Larsen, N. & Ayres, J. B. (2003). Role of context similarity in ABA, ABC and AAB renewal paradigms: Implications for theories of renewal and for treating human phobias. Learning & Motivation, 34, 410-436. [ Links ]
Vila, N. J. & Rosas, J. M. (2001). Renewal and spontaneous recovery after extinction in a causal learning task. Mexican Journal of Behavior Analysis, 27, 79-96. [ Links ]

