










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Psicología
versión impresa ISSN 0121-5469
Rev. colomb. psicol. vol.22 no.2 Bogotá jul./dic. 2013
Coordinación en Ratas: Ajuste a Restricciones Interdependientes*
Coordination in Rats: Adjustment to Interdependent Restrictions
Coordenação em Ratos: Ajuste a Restrições Interdependentes
ALEJANDRO SEGURA
ARTURO BOUZAS
Universidad Nacional Autónoma de México, México, D.F., México.
La correspondencia relacionada con este artículo debe dirigirse a Alejandro Segura, e-mail: alejandrosegurab@gmail.com.
Facultad de Psicología, Universidad Nacional Autónoma de México, Av. Universidad 3004, Col. Copilco-Universidad, CP 04510, Coyoacán, México, D.F.
Cómo citar este artículo: Segura, A. & Bouzas, A. (2013). Coordinación en ratas: ajuste de restricciones interdependientes.
Revista Colombiana de Psicología, 22(2), 321-331.
ARTÍCULO DE INVESTIGACIÓN CIENTÍFICA
RECIBIDO: 13 DE JUNIO DE 2013 - ACEPTADO: 23 DE SEPTIEMBRE DE 2013
Resumen
El estudio del ajuste conductual a entornos donde las interacciones entre los individuos y sus consecuencias son interdependientes ha derivado en procedimientos que limitan la interacción y restringen la operante a la simple simultaneidad de acciones sobre operandos individuales. En este estudio se evaluó una metodología alterna, en la que se expuso a pares de ratas a ir de un lugar a otro. El acceso al alimento fue contingente al trabajo individual o conjunto, en función de las posibilidades de interacción directa, determinadas por la ubicación de los sujetos. Se identificó coordinación únicamente en las diadas expuestas a interacción directa. El protocolo propuesto permite estudiar el ajuste a pagos interdependientes producto de patrones de acción conjunta.
Palabras clave: patrones de coordinación, restricciones interdependientes, propuesta metodológica, elección, ratas.
Abstract
The study of behavioral adjustment to environments in which interactions among individuals and their consequences are interdependent has resulted in procedures that limit interaction and restrict the operant to the simple simultaneity of actions on individuals' operands. This study evaluated an alternative methodology in which pairs of rats were made to go from one place to another. Access to food was contingent upon individual or joint work, depending on the possibilities of direct interaction determined by the location of the subjects. Coordination was identified only in the pairs that had direct interaction. The suggested protocol makes it possible to study the adjustment to interdependent payoffs resulting from joint action patterns.
Keywords: coordination patterns, interdependent constraints, methodological proposal, choice, rats.
Resumo
O estudo do ajuste de conduta a ambientes nos quais as interações entre os indivíduos e suas consequências são interdependentes deriva em procedimentos que limitam a interação e restringem a operante à simples simultaneidade de ações sobre operandos individuais. Neste estudo, avaliou-se uma metodologia alterna, na qual se expôs a pares de ratos a ir de um lugar a outro. O acesso ao alimento foi contingente ao trabalho individual ou conjunto, em função das possibilidades de interação direta, determinadas pela localização dos sujeitos. Identificou-se coordenação unicamente nas díades expostas à interação direta. O protocolo proposto permite estudar o ajuste a pagamentos interdependentes produto de padrões de ação conjunta.
Palavras-chave: padrões de coordenação, restrições interdependentes, proposta metodológica, eleição, ratos.
LA DIFICULTAD para replicar entornos donde las consecuencias de la conducta individual no dependen exclusivamente de la propia conducta, sino que son interdependientes de la conducta de otros, así como el patrón de actividades que emerge bajo estas condiciones, han hecho que la comprensión de fenómenos como la cooperación y el altruismo sea incompleta. Si bien diversas teorías y mecanismos han sido propuestos para su explicación (empatía, reciprocidad, inteligencia social, descuento temporal), la identificación y cuantificación de los patrones de actividad que emergen del trabajo conjunto, sincronía, coordinación, entre otros, y que han sido observados en condiciones naturales (e.g., caza cooperativa), aún no son claras. En muchos casos simplemente no se tienen en cuenta, y se aísla completamente a los sujetos, lo que hace cuestionable que los hallazgos obtenidos sean auténticas instancias de los fenómenos estudiados (Noë, 2006; Segura & Gutiérrez, 2006).
Uno de los debates más fuertes en el área tiene que ver con el hecho de que este tipo de comportamientos son ajustes conductuales a entornos con estructuras sociales complejas y presentes en unas pocas especies con alto grado de encefalización e inteligencia social (véase Drea & Carter, 2009). En gran medida esta hipótesis surge de la observación natural de patrones de coordinación y colaboración en primates, leones y mamíferos marinos (Busse, 1978; Rossbach, 1999; Scheel & Packer, 1991). Por ejemplo, al estudiar la caza cooperativa en chimpancés, C. Boesch y Boesch (1989) identificaron los patrones que emergen al actuar conjunto, indicando los grados de ajuste espacio-temporal; desde la simple similitud de actos (similaridad), a acciones ejecutadas al unísono (sincronía), el ajuste de posición y velocidad en espacio y tiempo (coordinación), hasta actos complementarios realizados en el mismo espacio y tiempo (colaboración). La clasificación propuesta por los autores sigue siendo útil y se ha ajustado al estudio de estos fenómenos en otras especies (véase Drea & Carter, 2009).
Desafortunadamente, las investigaciones de laboratorio desatienden la identificación de estos patrones, y centran su interés en el estudio del altruismo y la cooperación: fenómenos definidos a partir de la simetría o no en los pagos obtenidos (véase Brosnan & de Waal, 2002; Busse, 1978; Hamilton, 1964; Krebs & Davies, 1993; Locey, Jones, & Rachlin, 2013) o el tipo de relación entre los actores implicados (véase Rutte & Taborsky, 2008; Trivers, 1971). Los protocolos desarrollados para su estudio (e.g., dilema del prisionero, solución de problemas cooperativos) enfatizan en uno o varios de estos elementos, lo cual limita la interacción entre los sujetos y hace que la observación de los patrones de actividad no sea una tarea sencilla.
Los hallazgos experimentales han sido contradictorios. Algunas veces los sujetos son ineficientes para desarrollar la tarea, se observa preferencia por el trabajo individual; el número de sesiones de entrenamiento y moldeamiento es muy extenso y, generalmente, los resultados obtenidos son inconsistentes. Dichos resultados son atribuidos a habilidades y demandas cognitivas muy altas o, cuando se observa preferencia por el trabajo conjunto, esta suele ser explicada como producto de la coacción azarosa o coordinación deliberada entre los individuos (véase Drea & Carter, 2009; Plotnik, Lair, Suphachoksahakun, & de Waal, 2011). La inconsistencia en los resultados obtenidos puede ser producto de que los sujetos simplemente no se ajustan a las restricciones, probablemente porque los protocolos experimentales no replican la estructura de los entornos naturales que generan las formas de interacción a estudiar.
En contraste con los resultados obtenidos y las premisas que subyacen a su estudio (e.g., alto grado de encefalización), actualmente se han multiplicado los intentos por replicar la cooperación, el mutualismo y el altruismo en diferentes especies, entre ellas, hienas -Crocuta crocuta- (Drea & Carter, 2009), ratas -Rattus norvegicus- (Rutte & Taborsky, 2008; Schuster & Perelberg, 2004; Segura & Clavijo, 2010; Segura & Gutiérrez, 2006), arrendajos azules -Cyanocitta cristata- (Clements & Stephens, 1995; Stephens, McLinn, & Stevens, 2002), cuervos -Corvus frugilegus- (Seed, Clayton, & Emery, 2008) y humanos (Brown & Rachlin, 1999; Locey, Safin, & Rachlin, 2013).
El ajuste conductual en estas especies, similar a los hallazgos en primates (Chalmeau, 1994), implica una exposición prologada a las contingencias, el desarrollo de varias fases de entrenamiento e incluso el entrenamiento de la respuesta objetivo (Drea & Carter, 2009; Rutte & Taborsky, 2008; Scheid & Noë, 2010; Schuster, 2002). Moldear la respuesta objetivo le resta validez a los resultados obtenidos, cuando no hay evidencia de que la especie objeto de estudio presente ese comportamiento en entornos naturales o que se haya observado en ambientes controlados sin entrenamiento previo.
Investigaciones experimentales para el estudio de comportamientos altruistas y cooperativos en una especie tan simple como la rata (Rattus norvegicus) han aumentado (Bartal, Decety, & Mason, 2011; Lopuch & Popik, 2011; Rutte & Taborsky, 2008; Schneeberger, Dietz, & Taborsky, 2012). Sin embargo, no se cuenta aún con procedimientos que faciliten la emergencia de actividades sincrónicas y coordinadas en esta especie; y, excepto en las tareas desarrolladas por Schuster (2002), Schuster y Perelberg (2004), y Segura y Gutiérrez (2006), los autores normalmente moldean la conducta objetivo, por lo cual no es claro si el ajuste conductual es producto de las restricciones, el entrenamiento, el uso de manipulandos compartidos o la posibilidad de contacto directo entre los sujetos. ¿Cómo determinar que el comportamiento observado es el ajuste de la conducta interindividual a consecuencias interdependientes?
Para responder a esta pregunta se diseñó un protocolo experimental que permitiera: (a) identificar los patrones de actividades conjuntas que pueden emerger bajo restricciones y (b) detectar si las ratas realmente se ajustan a restricciones interdependientes cuando tienen contacto directo entre sí, aislando los efectos que el entrenamiento (moldeamiento) de una respuesta específica o el uso de operandos compartidos (e.g., palanca, balín) pudieran generar sobre el comportamiento.
Se seleccionó la tarea más sencilla posible en la búsqueda de alimento en esta especie: ir de un lugar a otro cruzando por uno de dos corredores posibles. La obtención de alimento se hizo contingente a la elección de ambos sujetos, lo que permitió la interacción directa entre ellos (condición de interdependencia directa, ITD). Para evaluar y comparar la efectividad de este procedimiento con los protocolos tradicionales, los sujetos fueron asignados a compartimientos individuales, contiguos y separados por una división transparente. Los pagos obtenidos podían depender de la elección conjunta (condición de interdependencia presencia, ITP) o individual (condición de independencia presencia, IDP).
Método
Sujetos
Se emplearon 8 ratas wistar macho (Rattus norvegicus), experimentalmente ingenuas y de 70 días de edad al comienzo del estudio. Fueron alojadas en una habitación del bioterio de la Facultad de Psicología de la Universidad Nacional Autónoma de México (UNAM). Se asignaron individualmente a cajas habitacionales de acrílico, cada una con las siguientes dimensiones: 22 cm de alto, 25 cm de ancho y 45 cm de largo, para una superficie disponible total de 1,125 cm2. Durante 10 días los sujetos fueron habituados a las condiciones individuales de alojamiento y se realizó la línea de base del peso corporal en condiciones de libre acceso al alimento. Posteriormente, se redujo de forma progresiva la cantidad de alimento suministrada (e.g., 2 gr diarios), hasta que los sujetos alcanzaron el 85% de su peso corporal; este porcentaje se mantuvo hasta el final del experimento, al restringir el acceso diario a ± 9 gr de alimento, 2 horas después de concluida la sesión experimental. El ciclo luz-oscuridad fue de 12*12 horas (iniciando luz a las 7:00 a.m.). Las sesiones experimentales se realizaban 6 horas después de iniciado el ciclo de luz. Se mantuvo la temperatura ambiente a 21° ± 2°centígrados. El experimento se realizó de acuerdo con la Norma Oficial Mexicana para la producción, cuidado y uso de los animales del laboratorio (NOM-062-Z00-1999).
Instrumento
Se diseñó un instrumento compuesto por dos cajas superpuestas divididas por un piso transparente (véase Figura 1-B). Cada caja estaba compuesta por dos cajas inicio-meta y una plataforma de 100 cm de largo y 30 cm de ancho. Esta plataforma tenía una división central opaca que dividía la plataforma en dos corredores de 50 cm de largo y dos zonas de elección (véase Figura 1-A).
Procedimiento
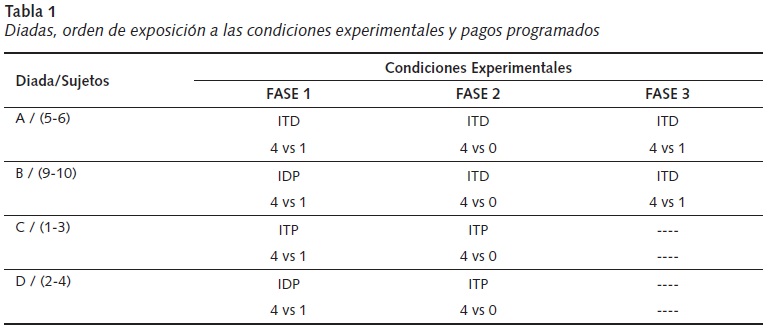
Se asignaron aleatoriamente los 8 sujetos a cuatro diadas. Durante dos sesiones de 20 min al día, se les permitió, en parejas, la libre exploración de la caja experimental. Posteriormente, los sujetos fueron expuestos a las condiciones y fases experimentales según fueron asignados (véase Tabla 1).
Cada sesión experimental estuvo compuesta por dos bloques de cuatro ensayos forzados (dos por opción) y dos bloques de 10 ensayos libres (20 por sesión). El primer y segundo bloque de ensayos forzados antecedía al primer y segundo bloque de ensayos libres, respectivamente. Un ensayo libre comenzaba con ambos sujetos en cajas de inicio del mismo lado; luego de 5 s, las compuertas de las cajas se abrían y los sujetos podían ir hasta la caja meta, pasando por uno de los dos corredores, donde tenían acceso o no al alimento (pellets), según la restricción que estaba operando (véase Tabla 1); posteriormente, las compuertas de la caja meta se cerraban y después de 3 s iniciaba un nuevo ensayo, ahora en la dirección opuesta.
En los ensayos forzados, la operante se reforzaba contingentemente a la condición experimental evaluada. Se bloqueaba el acceso a una de las dos opciones durante dos ensayos consecutivos, luego se hacía lo mismo para la otra opción. Se contaba como ensayo perdido cuando uno de los sujetos había recorrido completamente uno de los corredores al tiempo que el otro no hubiese iniciado su recorrido por ninguno de ellos. A continuación, se describen las restricciones experimentales:
1. Interdependencia-Directa (ITD). Ambos sujetos de la diada estaban en el mismo compartimiento (caja superior) y podían elegir uno de los dos corredores. Si tomaban idéntico corredor obtenían cuatro pellets, si elegían corredores diferentes obtenían uno o cero pellets, según la fase experimental (véase Tabla 1).
2. Interdependencia-Presencia (ITP). Al igual que en la condición anterior, los pagos obtenidos dependían de la elección conjunta (igual o diferente corredor). La diferencia consistió en que uno de los sujetos fue asignado siempre a la caja superior y el otro a la inferior.
3. Independencia-Presencia (IDP). Uno de los sujetos era siempre ubicado en la caja superior y el otro en la inferior. Al elegir uno de ellos el corredor del lado izquierdo generaba acceso a cuatro pellets y un pellet si elegía la otra opción. Para el otro sujeto de la diada, elegir la opción del lado derecho le permitía obtener cuatro pellets, si elegía el corredor izquierdo obtenía solamente uno. A diferencia de las condiciones de interdependencia, la elección de cada uno de los sujetos no interfería en lo que el otro obtenía, era independiente.
La asignación a las condiciones experimentales, el cambio entre fases y el número de sesiones al que fueron expuestas cada una de las cuatro diadas se realizaron con el fin de evaluar el ajuste a restricciones interdependientes producto del patrón de actividades conjuntas. Se comparó la metodología tradicional, en la cual los sujetos son aislados en compartimientos diferentes, con un protocolo que permite la interacción directa entre ellos. Por esta razón, una de las diadas fue expuesta en todas las condiciones a ITD (metodología propuesta), otra a la metodología tradicional (ITP) y las dos restantes iniciaron en independencia presencia (IDP) y luego fueron asignadas a uno de los dos protocolos evaluados.
Resultados
Los análisis presentados se basan en la frecuencia relativa de ensayos en los que los sujetos obtenían cuatro pellets (proporción alta). Como se piloteó una metodología, el único criterio a priori fue establecer en 120 (seis sesiones) el número mínimo de ensayos que componían cada condición experimental. El cambio de una fase experimental a otra obedeció a la observación de una tendencia de ajuste o no de la conducta a las restricciones evaluadas.
Las figuras están compuestas por un polígono de frecuencias y un histograma de barras. En el primero, se presenta la frecuencia de ensayos en que los sujetos de cada diada elegían la opción asociada con el mayor pago, durante todas las condiciones y organizadas de forma consecutiva. Como en las condiciones de interdependencia (ITD-ITP) los pagos dependen de la elección conjunta, las frecuencias relativas son idénticas entre los sujetos; por esta razón, se observa una sola línea. En el histograma se presentan los promedios obtenidos en las últimas seis sesiones de cada una de las fases.
Como se observa en la Figura 2, la frecuencia relativa de ensayos en la que ambos sujetos tomaban el mismo corredor varió en función de las condiciones experimentales. En la primera fase, la proporción fue muy baja (x, .11); aumentó de forma gradual en la segunda fase (x, .73), cuando elegir corredores diferentes no era reforzado y hacía así más salientes las consecuencias de la elección opuesta. En la última fase, la preferencia de los sujetos por elegir la misma opción se acentuó (x, .90), aunque las restricciones fueron idénticas a las de la primera fase.
En la Figura 3 se puede observar el ajuste de la conducta individual de los sujetos 9 y 10 a restricciones independientes (Fase 1). Al final de la fase ambos sujetos muestran preferencia por el corredor asociado a la mayor cantidad (x, .90: .92, respectivamente). El cambio a restricciones interconectadas (Fase 2) genera que la distribución de respuestas conjuntas sea inicialmente baja y aumente gradualmente. Similar a los sujetos de la diada A, la preferencia por elegir el mismo corredor se acentuó en la última fase, llegando al 82%.
Es importante resaltar que al final de la Fase 1 (IDP) los sujetos tomaban consistentemente corredores opuestos. Ese ajuste hace más interesantes los resultados observados en la Fase 2, donde la elección conjunta del mismo corredor aumentó de forma gradual, haciendo evidente la sensibilidad de los sujetos a restricciones interdependientes.
En los sujetos expuestos a contingencias de interdependencia presencia (ITP), se encontró insensibilidad a las restricciones. El promedio de respuestas en que ambos sujetos de la diada elegían la misma ruta, igual corredor uno arriba del otro, fue de 0.49 en la primera fase y de 0.48 cuando no obtenían nada si tomaban corredores opuestos (Fase 2). El cambio en la distribución de sus respuestas, entre las fases y a lo largo de las sesiones, fue mínimo y muy cercano al azar (véase Figura 4). Por esta razón, no se realizó una tercera fase de reversión a las condiciones de la Fase 1.
Es importante señalar que, aunque en la segunda fase no obtenían nada si elegían caminos diferentes, no se observó un ajuste similar al presentado por los sujetos de la diada A cuando fueron expuestos a idéntica condición. Estos resultados indican que la asignación a compartimientos diferentes (metodología tradicional) imposibilitó el ajuste a restricciones interdependientes.
El ajuste de la conducta individual a las restricciones impuestas a los sujetos de la diada D fue poco estable (véase Figura 5). En la condición donde los pagos obtenidos eran independientes de la elección del compañero (IDP), se observó variabilidad en las distribuciones. Los promedios obtenidos por los sujetos 2 y 4 en esta fase (x, .88: .53, respectivamente) indican que solo el sujeto 2 se ajustó a la restricción individual. Cuando los pagos obtenidos fueron interdependientes (ITP), el promedio de la distribución observada estuvo sobre el azar (x, .50). Esta insensibilidad a restricciones interdependientes determinó que no se realizara una tercera fase.
Discusión
La tarea evaluada en este pilotaje se constituye en un protocolo experimental útil para replicar patrones de coordinación en ratas, cuando se permite el contacto directo entre los sujetos y los pagos obtenidos dependen de la elección conjunta. En contraste, no permite replicar estos patrones al emplear una metodología tradicional, asignando los sujetos a compartimientos separados y restringiendo su interacción.
Los resultados sugieren que la conducta inter-individual en ratas se ajusta a restricciones donde los pagos obtenidos son mediados por la conducta del otro. Dos aspectos fundamentales observados en los entornos de interdependencia directa (ITD) soportan este hallazgo. Primero, la dinámica de ajuste conductual identificada en las diadas A y B (Fases 2 y 3) y, segundo, la emergencia de patrones de actividad (no entrenados) que antecedían la elección conjunta del mismo corredor y, en consecuencia, generaban el acceso a cuatro pellets.
El ajuste conductual a restricciones de ITD se desarrolló de forma gradual, a medida que los sujetos experimentaban las consecuencias del trabajo conjunto (Fase 2), y se estabilizó rápidamente en la última fase. La preferencia por el trabajo conjunto fue muy similar en ambas diadas, al margen de haber sido expuestas a historias experimentales diferentes en la primera fase.
Observaciones casuales permitieron identificar tres patrones de actividad que emergieron conjuntamente al ajuste gradual: primero, se observó una pausa en la zona de elección y aumento en la velocidad de carrera al cruzar el corredor; luego, se identificó un patrón de pausa-seguimiento, donde uno de los sujetos elegía un corredor y el otro "copiaba" esta elección (no se observaron actos complementarios, distribución de tareas líder-seguidor). Estos patrones fueron observados en la Fase 2, en ambas diadas. Por último, correlacionado con el incremento en la preferencia por elegir la misma opción (Fase 3), los sujetos establecían contacto directo en la zona de elección, acercando sus hocicos y vibrisas, y luego elegían idéntico corredor (para evidencia audio-visual, véase Segura, 2013).
El nivel de interacción social (contacto directo) entre los sujetos facilitó que coordinaran sus acciones, lo cual permitió el ajuste gradual de la conducta inter-individual a restricciones interdependientes (ITD). En contraste, restringir la interacción de los sujetos, ubicándolos en compartimientos diferentes, imposibilitó el ajuste a pagos interdependientes y la emergencia de actividades coordinadas (diadas C y D). Adicionalmente, en entornos donde los pagos dependen únicamente de la conducta individual, la presencia de un compañero no parece tener efecto alguno en el ajuste a estas contingencias (IDP).
Las relaciones encontradas en el presente estudio son consistentes con los hallazgos reportados por Daniel (1942), quien encontró sincronía y coordinación en ratas, al permitirles la interacción directa en una situación experimental donde la evitación de un choque y la alimentación eran interdependientes de la conducta de ambos sujetos. Efectos similares se han reportado al permitir el contacto visual, auditivo y olfativo en ratas. Al respecto Gardner, Corbin, Beltramo y Nickell (1984) señalan que la posibilidad de observación entre sí facilita la cooperación, además de la emergencia de actividades coordinadas, siempre y cuando se utilicen manipulandos compartidos (Segura & Clavijo, 2010).
Los resultados reportados son consistentes con la evidencia de patrones de coordinación en ratas cuando se permite el contacto directo entre ellas (Schuster, 2002; Schuster & Perelberg, 2004) o cuando la operante se realiza con manipulandos compartidos, permitiendo el contacto visual entre los sujetos (Segura & Clavijo, 2010; Segura & Gutiérrez, 2006).
Es muy probable que las actividades no entrenadas en esta especie surjan como ajustes de posición y velocidad en el mismo espacio y tiempo (coordinación), y no sean simples actividades ejecutadas simultáneamente (sincronía), como los protocolos tradicionales facilitan. No se observaron actos complementarios (colaboración). Estos últimos podrían ser un reto para esta especie y obedecer a presiones selectivas diferentes en especies con un mayor grado de organización social (e.g., delfines, leones). Para probar estas hipótesis, es necesario que en replicaciones posteriores se registre sistemáticamente la actividad conjunta; se hace imprescindible cuantificar estos patrones.
La relativa facilidad con la que emergen patrones coordinados puede reflejar, parcialmente, una fuerte preparación biológica para su realización, y ser un subproducto de la vida gregaria en esta especie. Es probable que en ambientes naturales la rata participe en la búsqueda conjunta de alimento, siguiendo una ruta común y mostrando patrones similares para su captura y consumo. Desafortunadamente, no existen investigaciones de campo que hayan explorado esta posibilidad, aunque como señalan Koolhaas, De Boer, y Buwalda (2005), el paradigma central en la biología de la rata es su naturaleza social.
En síntesis, para determinar que el comportamiento observado en ambientes de laboratorio es el producto del ajuste de la conducta interindividual a consecuencias interdependientes, es imprescindible que los protocolos experimentales permitan la emergencia de patrones de actividad conjunta (e.g., coordinación, colaboración). Desafortunadamente, gran parte de las tareas experimentales emplean el paradigma del dilema del prisionero iterado (Gardner et al., 1984; Stephens et al., 2002) o aíslan a los sujetos en cámaras independientes, y limitan la operante a la simple simultaneidad de las acciones (Lopuch & Popik, 2011). Bajo estas condiciones, la validez de las conductas observadas como instancias de cooperación o altruismo siempre será cuestionable.
Como señala Noë (2006), aún hay un abismo enorme entre los experimentos donde los sujetos interactúan con una máquina y los experimentos donde los sujetos ajustan sus acciones en función de la información recibida de un compañero. Tal vez la ruta para integrar los hallazgos en el área sea identificar y analizar cuantitativamente los patrones conductuales que emergen en este tipo de entornos, señalando las similaridades y diferencias en distintas especies y generando así una perspectiva comparativa más amplia.
Agradecimientos
El primer autor reconoce una deuda de gratitud con el Consejo Nacional de Ciencia y Tecnología (CONACYT), por la beca n.° 239774, para realizar sus estudios de doctorado. Agradecemos a José Luis Baroja por su valiosa colaboración en la representación gráfica de los resultados.
* La metodología evaluada en este estudio hace parte del pilotaje de la tesis doctoral del primer autor. El segundo autor dirigió y orientó la presente propuesta como tutor del primero. Esta investigación fue financiada por el proyecto 104396 de CONACYT. Resultados preliminares fueron presentados en el XXI Congreso Mexicano de Análisis de la Conducta, septiembre de 2011, Acapulco, México, y en el XIX Congreso Mexicano de Psicología, octubre de 2011, Cancún, México.
Referencias
Bartal, I., Decety, J., & Mason, P. (2011). Empathy and pro-social behavior in rats. Science, 334(6061), 1427-1430. [ Links ]
Boesch, C. & Boesch, H. (1989). Hunting behavior of wild chimpanzees in the Tai National Park. American Journal of Physical Anthropology, 78, 547-573. [ Links ]
Brosnan, S. & de Waal, F. (2002). A proximate perspective on reciprocal altruism. Human Nature, 13, 129-152. [ Links ]
Brown, J. & Rachlin, H. (1999). Self-control and social cooperation. Behavioural Processes, 47, 65-72. [ Links ]
Busse, C. (1978). Do chimpanzees hunt cooperatively? The American Naturalist, 112, 767-770. [ Links ]
Chalmeau, R. (1994). Do chimpanzees cooperate in a learning task? Primates, 35, 385-392. [ Links ]
Clements, K. & Stephens, D. (1995). Testing models of non-kin cooperation: Mutualism and the prisoner's dilemma. Animal Behaviour, 50, 527-535. [ Links ]
Daniel, W. J. (1942). Cooperative problem solving in rats. The Journal of Comparative Psychology, 34, 361-368. [ Links ]
Drea, C. & Carter, A. (2009). Cooperative problem solving in a social carnivore. Animal Behaviour, 78, 967-977. [ Links ]
Gardner, R., Corbin, T., Beltramo, J., & Nickell, G. (1984). The prisoner's dilemma game and cooperation in the rat. Psychological Reports, 55, 687-696. [ Links ]
Hamilton, W. (1964). The genetical evolution of social behaviour. Journal of Theoretical Biology, 7, 1-52. [ Links ]
Koolhaas, J., De Boer, S., & Buwalda, B. (2005). Stress. En I. Whishaw. & B. Kolb (Eds.), The behavior of the laboratory rat (pp. 236-244). New York: Oxford University Press. [ Links ]
Krebs, J. R. & Davies, N. B. (1993). An introduction to behavioural ecology. London: Blackwell. [ Links ]
Locey, M., Jones, B., & Rachlin, H. (2013). Self-control and altruism. En G. J. Madden., W. V. Duve., T. D., Hackenberg., G.P., Hanley, & K. A. Lattal (Eds.), APA handbook of behavior analysis: Vol. 1. Methods and principles (pp. 463-48). Washington DC: APA Books. [ Links ]
Locey, M., Safin, V., & Rachlin, H. (2013). Social discounting and the prisoner's dilemma game. Journal of the Experimental Analysis of Behavior, 99, 85-97. [ Links ]
Lopuch, S. & Popik, P. (2011). Cooperative behavior of laboratory rats (Rattus norvegicus) in an instrumental task. Journal of Comparative Psychology, 125, 250-253. [ Links ]
Noë, R. (2006). Cooperation experiments: Coordination through communication versus acting apart together. Animal Behaviour, 71, 1-18. [ Links ]
Plotnik, J., Lair, R., Suphachoksahakun, W., & de Waal, F. (2011). Elephants know when they need a helping trunk in a cooperative task. PNAS, 108, 5116-5121. [ Links ]
Rossbach, K. (1999). Cooperative feeding among bottlenose dolphins (Tursiops truncatus) near Grand Bahama Island, Bahamas. Aquatic Mammals, 25, 163-167. [ Links ]
Rutte, C. & Taborsky, M. (2008). The influence of social experience on cooperative behavior. Behavioral Ecology and Sociobiology, 62, 499-505. [ Links ]
Scheel, D. & Packer, C. (1991). Group hunting behaviour of lions: A search for cooperation. Animal Behaviour, 41, 697-709. [ Links ]
Scheid, C. & Noë, R. (2010). The performance of rooks in a cooperative task depends on their temperament. Animal Cognition, 13, 545-553. [ Links ]
Schneeberger, K., Dietz., M., & Taborsky, M. (2012). Reciprocal cooperation between unrelated rats depends on cost to donor and benefit to recipient. Evolutionary Biology, 12, 1-7. [ Links ]
Schuster, R. (2002). Cooperative coordination as a social behavior: Experiments with an animal model. Human Nature, 13, 47-83. [ Links ]
Schuster, R. & Perelberg, A. (2004). Why cooperate? An economic perspective is not enough. Behavioural Processes, 66, 261-267. [ Links ]
Seed, A., Clayton, N., & Emery, N. (2008). Cooperative problem solving in rooks (Corvus frugilegus). Proceedings of the Royal Society B, 275, 1421-1429. [ Links ]
Segura, A. (2013). Coordinación en ratas [Archivo de video]. Recuperado de https://www.youtube.com/watch?v=I_F9vqNq49A&_feature=youtu.be [ Links ]
Segura, A. & Clavijo, A. (2010). Conductas cooperativas, altruistas y de competencia: formas de interacción social que emergen bajo exposición a diferentes relaciones de contingencia. (Tesis de maestría inédita). Universidad Nacional de Colombia, Bogotá, Colombia. [ Links ]
Segura, A. & Gutiérrez, G. (2006). Cooperación en ratas: efectos de la experiencia temprana. Revista Interamericana de Psicología, 40, 241-252. [ Links ]
Stephens, D., McLinn, C., & Stevens, J. (2002). Discounting and reciprocity in an iterated prisoner's dilemma. Science, 298, 2216-2218. [ Links ]
Trivers, R. (1971). The evolution of reciprocal altruism. Quarterly Review of Biology, 46, 35-57. [ Links ]

{kind=link}