










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCiencia en Desarrollo
Print version ISSN 0121-7488
Ciencia en Desarrollo vol.5 no.1 Tunja Jan./June 2014
Riqueza y Distribución Altitudinal de Gramíneas C3 y C4 en la Guayana Venezolana
Richness and Altitudinal Distribution of C3 and C4 Grasses in the Venezuelan Guayana
D. Giraldo-Cañasa,*
a,* Herbario Nacional Colombiano COL, Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá D. C., Colombia. Correo electrónico: dagiraldoc@unal.edu.co.
Recepción: 20-ago-13 Aceptación: 25-nov-13
Resumen
Se estudiaron la riqueza de especies y el patrón de distribución altitudinal de las gramíneas C3 y C4 en la Guayana venezolana. Las gramíneas están representadas en el gradiente altitudinal por 411 especies, de las cuales, 170 presentan la vía fotosintética C3, y 241, la C4. Las gramíneas C3 y C4 están diversificadas en las tierras bajas, no obstante, las C3 exhiben un patrón particular; así, en las tierras bajas se distribuyen ecológicamente en hábitats sombreados (sotobosques). En las tierras altas, las especies C3 desplazan a las C4, que están mínimamente representadas. El patrón de distribución altitudinal de las gramíneas C3 y C4 sigue la regla de Rapoport.
Palabras clave: Diversidad en gradientes altitudinales, Gramíneas C3 y C4, Flora de la Guayana, Gramíneas neotropicales, Regla de Rapoport.
Abstract
The species richness and altitudinal distribution pattern of C3 and C4 grasses from the Venezuelan Guayana were studied. The grasses are represented along the altitudinal gradient by 411 species, of which 170 species are C3 and 241 species are C4. The highest species richness is reached in lowlands. The C3 and C4 grasses are more diversified in the lowlands, however, the C3 species show and interesting pattern, which are mainly distributed in understories of lowlands. In the highlands, the C3 species replace the C4 species, which diminish their importance. The altitudinal distribution pattern of C3 and C4 grasses follows the Rapopor’s rule.
Key words: Diversity along altitudinal gradients, C3 and C4 grasses, Flora of the Guayana, Neotropical grasses, Rapoport’s rule.
1. Introducción
La región biogeográfica de la Guayana pertenece al Escudo Precámbrico Guayanés, que forma un arco discontinuo de cerros, lajas, afloramientos rocosos, serranías, tepuyes y macizos en el norte de la Amazonia [véanse las referencias en [1]]; esta región constituye un área cercana a los dos millones de km2, se localiza en el norte de Sudamérica y comprende el sur de Venezuela y partes de Guyana, Surinam, Guayana Francesa, Colombia y Brasil [véanse las referencias en [1]]. La región Guayana posee una rica flora vascular estimada en cerca de 1.800 géneros (140 endémicos) y unas 15.000 especies, de las cuales un 40% tiene un área de distribución endémica de la región [2].
En esta gran región se encuentra la Guayana venezolana, que abarca cerca de 454.000 km2 [3]; esta comprende un amplio gradiente altitudinal, que va desde las tierras bajas hasta los 3.014 m de altitud en la sierra de La Neblina [4]. En la Guayana venezolana se han documentado 411 especies de gramíneas [5, 6], lo cual la convierte en la cuarta familia más diversificada en esta área. Las gramíneas se distribuyen de manera diferencial en la Guayana venezolana, debido, principalmente, al tipo de vía fotosintética de cada especie, a la altitud, a los hábitats -ya bien sea que éstos estén expuestos a la luz solar directa o a condiciones de sombrío-, al tipo de sustrato en el que crecen las especies [7] y a las condiciones de madurez o sucesión secundaria de los hábitats. Con base en estas características de la distribución de gramíneas en ambientes tropicales, este estudio tuvo como finalidad relacionar y analizar los posibles patrones en la distribución diferencial de las gramíneas C3 y C4 en todo el gradiente altitudinal de la Guayana venezolana; y para tal fin se trabajó con base en tres hipótesis: 1) Las diferencias fisiológicas de las gramíneas C3 y C4 se reflejan en sus patrones de distribución a lo largo de gradientes ambientales y climáticos [7, 8]; 2) Los gradientes altitudinales proveen un importante escenario para estudiar las relaciones de la distribución de especies y el clima, debido a la gran variación climática en pequeñas distancias geográficas [9], y 3) Dadas las características fisiológicas de las gramíneas, se espera una mayor representatividad de las C3 en las tierras altas.
1.1. Las gramíneas y sus vías fotosintéticas
Las gramíneas presentan las vías fotosintéticas C3 y C4, con algunas modificaciones fisiológicas y ecológicas adicionales; así, se tiene que las gramíneas C4 difieren significativamente de las C3 en su fisiología, bioquímica, anatomía, ultraestructura y requerimientos ambientales; a su vez, los factores climáticos limitantes y las adaptaciones ecológicas también difieren entre estas dos vías fotosintéticas [7, 10, 11]. Ahora bien, las gramíneas C4 representan cerca del 45% de la familia [12], y se caracterizan, generalmente, por un bajo punto de compensación de CO2 atmosférico, un rápido crecimiento, bajas tasas de fotorrespiración y de pérdida de agua por unidad de materia producida, un alto intervalo óptimo de temperatura, un alto punto de saturación lumínica y una anatomía foliar en corona o "Kranz" [véanse las referencias en [7]] (tabla 1, figura 1). Entre tanto, las gramíneas C3 presentan un alto punto de compensación de CO2 atmosférico, altas tasas de fotorrespiración, un intervalo óptimo de temperatura de 10-25 °C, un punto de saturación lumínica mucho menor que las plantas C4 y una anatomía foliar "No-Kranz" [véanse las referencias en [7]] (tabla 1, figura 2). Según [13], la fotosíntesis en las plantas C4 no está afectada por altas o bajas concentraciones de O2 atmosférico, mientras que la fotosíntesis en las C3 es inhibida por altas concentraciones de O2 y aumenta a bajas concentraciones de este.
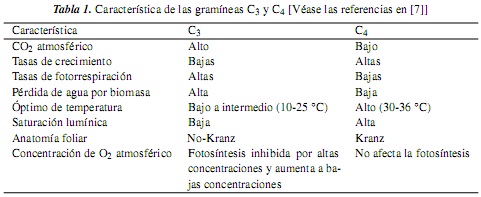
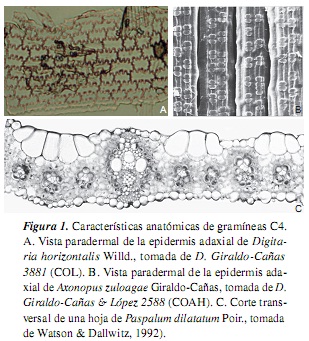
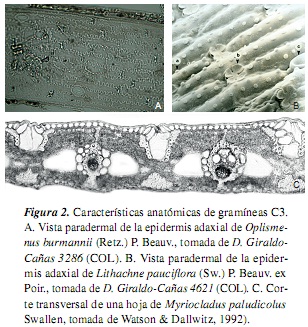
2. Experimental
La Guayana venezolana aquí reconocida sigue los límites geográficos, geológicos, florísticos y biogeográficos definidos por [2, 3, 4]. Los intervalos altitudinales se discriminaron con una amplitud de 500 m, siguiendo el uso frecuente de intervalos en diferentes investigaciones en gradientes altitudinales [véanse las referencias en [7]], lo que facilita las comparaciones y los análisis con otros estudios sobre distribución y riqueza de especies. Así, se definieron seis intervalos altitudinales, entre los 0 y los 3000 m de altitud (tabla 2). El inventario de especies y los datos de altitud se tomaron de [5, 6], por lo que no se repiten aquí. La clasificación de las gramíneas como plantas C3 o C4 se realizó con base en la información de la clasificación de las vías fotosintéticas consignada en numerosas obras [véanse las referencias en [7]].
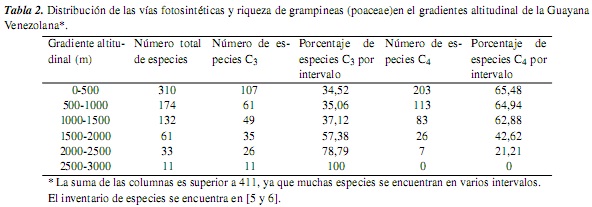
3. Resultados y discusión
De las 411 especies de gramíneas reconocidas para la Guayana venezolana [5, 6], 170 son C3 y 241 son C4. Ellas muestran una distribución diferencial dependiendo del intervalo altitudinal (tabla 2); así, las C4 son más ricas y más dominantes en las tierras bajas y medias (0-1500 m de altitud), y su riqueza y su representatividad van disminuyendo con la altitud; las C3, aunque presentan también un patrón de disminución de la riqueza de especies con la altitud, empiezan a ser más diversas y dominantes sobre las C4 a partir de los 1500 m de altitud (tabla 2). Estos datos se traducen en que hay un alto recambio de especies a medida que se incrementa la altitud, o sea, la diversidad beta es alta; esto también se ve reflejado en el aumento de los porcentajes de riqueza de especies por cada intervalo altitudinal, toda vez que las especies C3 empiezan a presentar -a partir de los 1500 m- mayores porcentaje de riqueza específica que las C4 (tabla 2), al igual que una mayor representatividad ecológica.
Este patrón de distribución se ajusta a las características y a los requerimientos ambientales de las gramíneas, tanto C3 como C4 (tabla 1), ya que en hábitats tropicales de montaña las temperaturas medias anuales van disminuyendo con la altitud, lo que favorece el alto recambio de especies C4 por C3, toda vez que estas últimas tienen un óptimo de temperatura bajo a intermedio (10-25 °C) (tabla 1). Asimismo, la concentración de O2 atmosférico va disminuyendo con la altitud, lo cual, a su vez, aumenta la fotosíntesis de las especies C3 (tabla 1); estas condiciones en ambientes tropicales de montaña favorecen el alto recambio de gramíneas C4 por gramíneas C3.
Si bien las especies C3 tienen una riqueza apreciable en las tierras bajas y medias (0-1500 m), nunca superan el 37,12% de la riqueza de cada intervalo altitudinal de las tierras bajas (tabla 2); no obstante, varias de sus especies, principalmente Bambusoideae y algunas Panicoideae, pueden ser más dominantes en las tierras bajas y medias que las C4, dado que presentan una distribución ecológica diferencial, ya que muchas de estas crecen en hábitats de sombra, los cuales favorecen el establecimiento de gramíneas C3, que presentan una saturación lumínica baja (tabla 1), característica presente en áreas de sombrío, y otras especies C3 se establecen en ambientes acuáticos de tierras bajas (principalmente especies de los géneros Hymenachne, Leersia, Luziola, Oryza, Phragmites, Sacciolepis, Steyermarkochloa), en donde las temperaturas son un poco más bajas y nunca hay estrés hídrico por déficit [7]; mientras que la mayoría de las especies C4 requieren de hábitats abiertos (saturación lumínica alta) y altas temperaturas (tabla 1). Adicionalmente, las gramíneas C4 tienen una alta razón de CO2 fijado/uso de agua, y, por lo tanto, están mejor adaptadas a las áreas muy cálidas, lo que podría respaldar la hipótesis de que esta vía fotosintética es una adaptación a este tipo de ambientes [14]. Así, las plantas C4 tienen una ventaja competitiva sobre las C3 en áreas de alta intensidad lumínica, elevadas temperaturas, altas concentraciones de O2 atmosférico y bajas concentraciones de CO2. Por otra parte, el mayor costo energético del metabolismo C4, en comparación con el del C3, lo excluye de hábitats sombreados y reduce su ventaja competitiva en hábitats fríos. Por lo tanto, la distribución de gramíneas C4 está climáticamente controlada [15], y su distribución corresponde, principalmente, con hábitats abiertos.
Es de destacar que ninguna de las gramíneas C4 se encuentra en sotobosques o en áreas sombreadas, distribución que muestra que las gramíneas de esta vía fotosintética no soportan las características ambientales propias de estos hábitats, como se mencionó anteriormente. Según [16], la ausencia de gramíneas C4 en hábitats de sombra puede ser debida a que no ha habido un suficiente tiempo para una adaptación evolutiva de esta vía fotosintética a este tipo de ambientes, dado que sólo se ha desarrollado recientemente en ambientes cálidos y áridos. Al respecto, [13] comentó que la vía C4 se desarrolló en gramíneas de origen tropical, durante el Mioceno, época en la cual las gramíneas C4 también se expandieron [17], a partir de múltiples orígenes [12, 17, 18]. Por otra parte, [17] mostraron que los orígenes de las C4 se remontan al Oligoceno (hace ca. 32 millones de años).
Todos los géneros son exclusivamente C3 o C4, salvo Aristida, el cual presenta ambas vías fotosintéticas, aunque casi todas sus especies son C4 [19] y solo una es C3, Aristida longifolia Trin. [20]. Al respecto, algunos autores [21, 22, 23] consideran que los géneros que presentan ambas vías fotosintéticas requieren de una revisión taxonómica-sistemática, ya que pueden considerarse no monofiléticos, puesto que las diferentes especies C3 o C4 de un mismo género no estarían relacionadas filogenéticamente; este tipo de revisión se ha dado recientemente en el género Panicum L. s.l., ya que se han empezado a reconocer varios géneros segregados de este con base en la combinación de varios caracteres, p. e. morfología, anatomía, datos moleculares y el tipo de vía fotosintética [24], lo que respalda las ideas iniciales de [21, 22, 23], entre otros.
En cuanto a la riqueza total de especies por gradiente altitudinal, tenemos que el intervalo más rico en especies es el que está comprendido entre el nivel del mar y los 500 m (310 especies), seguido por el segundo intervalo (500-1000 m), con 174 especies; mientras que los intervalos más altos presentan la menor riqueza de especies (tabla 2, figura 3). Estos datos se ajustan a los encontrados en algunas regiones colombianas estudiadas en un contexto de gradientes altitudinales [7, 25, 26], así como en Ecuador [27, 28], en las que las tierras bajas y medias (0-1500 m) tienen la mayor riqueza de gramíneas, no solo en número de especies, sino también en géneros y subfamilias. Este patrón de riqueza se ajusta a la regla de Rapoport [29, 30, 31], cuya ecuación para el total de especies de la Guayana venezolana es y = −0, 113x + 290, 6, con un R2 igual a 0,910 (figura 3), y = −0, 034x + 99, 51 (R2 = 0,903) para las especies C3 (figura 4), mientras que para las C4 la ecuación fue y = −0,079x + 191, 1 (R2 = 0,905) (figura 5). Por lo tanto, se deduce que muchas de las especies presentan tolerancias estrechas (alto recambio de especies), lo cual se traduce en una gran heterogeneidad espacial en los arreglos de las comunidades de gramíneas en la Guayana [1, 5].
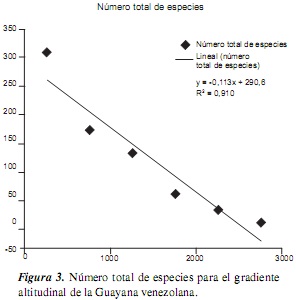
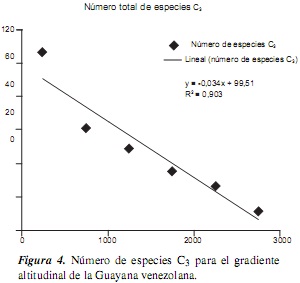
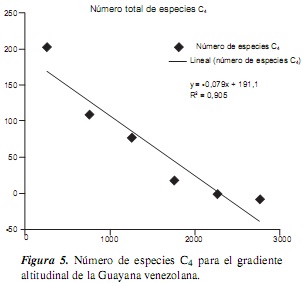
Por otra parte, los datos analizados para las gramíneas de la Guayana venezolana no se ajustan al "efecto del dominio medio" [30, 31, 32, 33, 34, 35], el cual contempla que la mayor riqueza de especies, en un contexto de gradientes altitudinales, se encuentra en las alturas intermedias, como el caso de la riqueza de mariposas en la serranía de Perijá (Colombia) [36, 37], el caso de varios grupos de vertebrados y plantas vasculares (Acanthaceae, Bromeliaceae, Pteridophyta, Solanaceae) [35, 38] o el caso de las especies de la familia Frullaniaceae de las hepáticas [39] en áreas montanas neotropicales.
4. Conclusiones
Las tierras bajas y medias (0-1500 m) están caracterizadas principalmente por gramíneas C4, mientras que las altas (1500-3000 m) están dominadas básicamente por gramíneas C3. El patrón de riqueza de especies en el contexto altitudinal -tanto para el total de especies como para la separación en especies C3 o C4- se ajusta a la regla de Rapoport, lo cual evidencia además, un alto recambio de especies a medida que se incrementa la altitud. Dicho patrón conduce a aceptar las dos primeras hipótesis de trabajo; en tanto que la tercera hipótesis no se respalda aquí en su totalidad, toda vez que las gramíneas C3 presentaron también la regla de Rapoport, pero a medida que se incrementa la altitud, desplazan a las C4 y, por lo tanto, su porcentaje de representatividad aumenta en los intervalos altitudinales mayores (1500-3000 m), al pasar de 34,52-37,12% en las tierras bajas y medias, a 57,38-100% en las tierras altas (tabla 2). Así, los datos aquí analizados en cuanto a la representatividad altitudinal de las gramíneas C3 se ajustan a lo hallado por [40], autor que realizó un análisis sobre la distribución de la familia Cyperaceae y sus vías fotosintéticas en los Andes colombianos.
En consecuencia, se podría concluir que las gramíneas responden de manera similar a las características ambientales asociadas con las vías fotosintéticas, independientemente de la región natural en la que se establecen, ya bien sea paleotropical, neotropical o templada. No obstante, por obvias razones, en las regiones templadas, los patrones responden según las estaciones, en donde los mejores estimadores del número de especies C3 y C4 son la media mínima y la media máxima de la temperatura del mes más cálido [41]. Por último, es necesario destacar que no se ha documentado en la Guayana venezolana ninguna gramínea C4 por encima de los 2500 m de altitud; no obstante, esta ausencia podría deberse a problemas de muestreo, ya que en áreas tropicales de alta montaña se han documentado varias especies C4 hasta cerca de los 4500 m de altitud [7, 26].
Agradecimientos
Quiero manifestar mi profundo reconocimiento al Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (Bogotá D. C.) por todas las facilidades que me brindó para la preparación de este trabajo. A la Prof. Dra. María Eugenia Morales (Universidad Pedagógica y Tecnológica de Colombia, Tunja) por su valiosa y permanente colaboración, así como por sus enriquecedores comentarios. A todos los miembros del Comité Editorial por su valiosa colaboración. A los evaluadores anónimos por sus diversos y valiosos comentarios. Esta contribución se deriva del proyecto "Estudios morfológicos, anatómicos y taxonómicos en gramíneas neotropicales (Fase II)" de la Universidad Nacional de Colombia, sede Bogotá D. C.
Referencias
[1] D. Giraldo-Cañas. "Las gramíneas (Poaceae) de la Guayana colombiana: análisis sobre su composición, riqueza, endemismo e invasión". Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, vol. 34, pp. 15-26, 2010. [ Links ]
[2] P. Berry, O. Huber & B. Holst. "Floristic analysis and phytogeography". Flora of the Venezuelan Guayana, vol. 1, pp. 161-190, 1995. [ Links ]
[3] P. Berry, B. Holst & K. Yatskievych. "Introduction". En: Flora of the Venezuelan Guayana, vol. 1, pp. xv-xx, 1995. [ Links ]
[4] O. Huber. "Geographical and physical features". Flora of the Venezuelan Guayana, vol. 1, pp. 1-61, 1995. [ Links ]
[5] G. Davidse, E. J. Judziewicz & F. O. Zuloaga. "Poaceae". Flora of the Venezuelan Guayana, vol. 8, pp. 1-297, 2004. [ Links ]
[6] D. Chitty & S. Nozawa. "Poa annua L. y Polypogon elongatus Kunth (Poaceae), dos nuevas especies exóticas para la flora de la Guayana venezolana". Acta Botanica Venezuelica, vol. 33, pp. 357-361, 2010. [ Links ]
[7] D. Giraldo-Cañas. "Distribución e invasión de gramíneas C3 y C4 (Poaceae) en un gradiente altitudinal de los Andes de Colombia". Caldasia, vol. 32, pp. 65-86, 2010. [ Links ]
[8] M. Cabido, N. Ateca, M. E. Astegiano & A. M. Anton. "Distribution of C3 and C4 grasses along an altitudinal gradient in Central Argentina". Journal of Biogeography, vol. 24, pp. 197-204, 1997. [ Links ]
[9] F. Lloret & J. M. González-Mancebo. "Altitudinal distribution patterns of bryophytes in the Canary Islands and vulnerability to climate change". Flora, vol. 206, pp. 769-781, 2011. [ Links ]
[10] W. D. Clayton & S. A. Renvoize. "Genera Graminum: Grasses of the world". Kew Bulletin, Additional series, vol. 13, pp. 1-389, 1999. [ Links ]
[11] B. F. Jacobs, J. D. Kingston & L. L. Jacobs. "The origin of grass-dominated ecosystems". Annals of the Missouri Botanical Garden, vol. 86, pp. 590-643, 1999. [ Links ]
[12] R. F. Sage. "The evolution of C4 photosynthesis". New Phytologits, vol. 161, pp. 341-370, 2004. [ Links ]
[13] R. L. Chazdon. "Ecological aspects of the distribution of C4 grasses in selected habitats of Costa Rica". Biotropica, vol. 10, pp. 265-269, 1978. [ Links ]
[14] C. A. Klink & C. A. Joly. "Identification and distribution of C3 and C4 grasses in open and shade habitats in São Paulo State, Brazil". Biotropica, vol. 21, pp. 30-34, 1989. [ Links ]
[15] B. F. Jacobs, J. D. Kingston & L. L. Jacobs. "The origin of grass-dominated ecosystems". Annals of the Missouri Botanical Garden, vol. 86, pp. 590-643, 1999. [ Links ]
[16] W. Pearcy & J. Ehleringer. "Comparative ecophysiology of C3 and C4 plants". Plant, Cell and Environment, vol. 7, pp. 1-13, 1984. [ Links ]
[17] A. Vicentini, J. C. Barber, S. S. Aliscioni, L. M. Giussani & E. A. Kellogg. "The age of the grasses and clusters of orogins of C4 phothosynthesis". Global Change Biology, vol. 14, pp. 2963-2977, 2008. [ Links ]
[18] P. A. Christin, N. Salamin, E. A. Kellogg, A. Vicentini & G. Besnard. "Integrating phylogeny into studies of C4 variation in the grasses". Plant Physiology, vol. 149, pp. 82-87, 2009. [ Links ]
[19] L. Watson & M. J. Dallwitz. "The grass genera of the world: descriptions, illustrations, identification, and information retrieval; including synonyms, morphology, anatomy, physiology, phytochemistry, cytology, classification, pathogens, world and local distribution, and references". Versión: 28 de julio de 2014. http://deltaintkey.com, 1992 (onwards). [ Links ]
[20] R. Cerros-Tlatilpa & J. T. Columbus. "C3 Photosynthesis in Aristida longifolia: Implication for photosynthetic diversification in Aristidoideae (Poaceae)". American Journal of Botany, vol. 96, pp. 1379-1387, 2009. [ Links ]
[21] D. Moss, G. Krenzer & A. Brun. "Carbon dioxide compensation points in related plant species". Science, vol. 164, pp. 187-188, 1969. [ Links ]
[22] W. Brown. "Variations in anatomy, associations and origins of Kranztissue". American Journal of Botany, vol. 62, pp. 395, 1975. [ Links ]
[23] S. Waller & K. Lewis. "Occurrence of C3 and C4 photosynthesis pathway in North American grasses". Journal Range and Management, vol. 32, pp. 12-28, 1979. [ Links ]
[24] F. O. Zuloaga & D. Giraldo-Cañas. "Panicum (Poaceae)". Flora de Colombia, vol. 30, pp. 1-102. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá D. C., 2013. [ Links ]
[25] D. Giraldo-Cañas. "Diversidad y distribución de gramíneas C3 y C4 (Poaceae) en diferentes gradientes altitudinales de la serranía de Perijá (región Caribe, Colombia)". En: J. O. Rangel Ch. (ed.), Serie Colombia Diversidad Biótica VIII: Media y baja montaña de la serranía de Perijá, pp. 229-244. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá D. C., 2009. [ Links ]
[26] D. Giraldo-Cañas. "Las gramíneas en Colombia: Riqueza, distribución, endemismo, invasión, migración, usos y taxonomías populares". Biblioteca José Jerónimo Triana, vol. 26, pp. 1-380. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá D. C., 2013. [ Links ]
[27] S. Lægaard. "Biogeography of Ecuadorian grasses". Arnaldoa, vol. 6, pp. 185-196, 1999. [ Links ]
[28] M. Kessler. "The elevational gradient of Andean plant endemism: varying influences of taxon-specific traits and topography at different taxonomic levels". Journal of Biogeography, vol. 29, pp. 1159-1165, 2002. [ Links ]
[29] G. C. Stevens. "The elevational gradient in altitudinal range: an extension of Rapoports latitudinal rule to altitude". American Naturalist, vol. 140, pp. 893-911, 1992. [ Links ]
[30] N. Sanders. "Elevational gradients in ant species richness: area, geometry, and Rapoports rule". Ecography, vol. 25, pp. 25-32, 2002. [ Links ]
[31] R. J. Guerrero & C. E. Sarmiento. "Distribución altitudinal de hormigas (Hymenoptera, Formicidae) en la vertiente noroccidental de la Sierra Nevada de Santa Marta (Colombia)". Acta Zoológica Mexicana (nueva serie), vol. 26, pp. 279-302, 2010. [ Links ]
[32] K. R. Colwell & D. C. Lees. "The mid-domain effect: geometric constraints on the geography of species richness". Trends in Ecology and Evolution, vol. 15, pp. 70-76, 2000. [ Links ]
[33] J. A. Grytnes & O. R. Vetaas. "Species richness and altitude: a comparison between Null Models and interpolated plant species richness along the Himalayan altitudinal gradient, Nepal". American Naturalist, vol. 159, pp. 294-304, 2002. [ Links ]
[34] M. R. Silman. "Plant species diversity in Amazonian forest". En: M. B. Bush & J. R. Flenley (eds.), Tropical rainforest responses to climatic change, pp. 269-294. Springer-Praxis Publishing, Berlín, 2007. [ Links ]
[35] J. Adams. "Species richness. Patterns in the diversity of life". Springer-Praxis Publishing, Berlín, 2009. [ Links ]
[36] H. W. Pulido-B. & M. G. Andrade-C. "Mariposas de las partes altas de la serranía de Perijá". En: J. O. Rangel-Ch. (ed.), Colombia Diversidad Biótica V. La alta montaña de la serranía de Perijá, pp. 235-248. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá D. C., 2007. [ Links ]
[37] H. W. Pulido-B. & M. G. Andrade-C. "Las mariposas de la serranía de Perijá". En: J. O. Rangel Ch. (ed.), Colombia Diversidad Biótica VIII: Media y baja montaña de la serranía de Perijá, pp. 509-559. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá D. C., 2009. [ Links ]
[38] M. Kessler & J. Kluge. "Diversity and endemism in tropical montane forests ? from patterns to processes". Biodiversity and Ecology Series, vol. 2, pp. 35-50, 2008. [ Links ]
[39] R. Gradstein & J. Uribe-M. "A synopsis of the Frullaniaceae (Marchantiphyta) from Colombia". Caldasia, vol. 33, pp. 347-376, 2011. [ Links ]
[40] C. A. Suárez Rojas. "Análisis de la distribución altitudinal de Cyperaceae en los Andes de Colombia, con base en sus vías fotosintéticas". Trabajo de grado, Departamento de Biología, Universidad Nacional de Colombia, Bogotá D. C., 2014. [ Links ]
[41] T. R. Wentworth. "Distribution of C4 plants along environmental and compositional gradients in southeastern Arizona". Vegetatio, vol. 52, pp. 21-34, 1983. [ Links ]
