










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista MVZ Córdoba
versión impresa ISSN 0122-0268versión On-line ISSN 1909-0544
Rev.MVZ Cordoba v.13 n.2 Córdoba mayo/agos. 2008
ISOLATION OF Rickettsia bellii FROM Amblyomma ovale AND Amblyomma incisum TICKS FROM SOUTHERN BRAZIL
AISLAMIENTO DE Rickettsia bellii A PARTIR DE GARRAPATAS Amblyomma ovale Y Amblyomma incisum PROCEDENTES DEL SUR DE BRASIL
Richard Pacheco,*1 Ph.D, Simone Rosa,1 MVZ, Leonardo Richtzenhain,1 Ph.D, Matias P. J. Szabó,2 Ph.D, Marcelo B. Labruna,1 Ph.D.
1University of São Paulo, Faculty of Veterinary Medicine, São Paulo, SP, Brazil 2Federal University of Uberlândia, Faculty of Veterinary Medicine, Uberlândia, MG, Brazil *Correspondencia: pachecorc@hotmail.com
Recibido: Julio 4 de 2008; Aceptado: Agosto 1 de 2008
ABSTRACT
Objective. To isolate and characterize rickettsiae from the ticks Amblyomma ovale and Amblyomma incisum collected in the state of São Paulo. Materials and methods. Adult, free-living A. ovale and A. incisum were collected in an Atlantic rainforest area in the state of São Paulo, Brazil. Each tick was tested using the hemolymph assay; samples from positive ticks were placed in shell vials in order to isolate rickettsiae and subsequently grown in Vero cells. Amplification of three rickettsial genes (gltA, htrA and ompA) was attempted using polymerase chain reaction (PCR) for each isolate obtained. Amplicons were subsequently sequenced. Results. A total of 388 A. incisum and 50 A. ovale were collected. Only one A. incisum and one A. ovale were hemolymph-test positive. Rickettsiae were successfully isolated from these ticks; however establishment in Vero cell culture was successful only for the isolate from A. ovale. Bacterial contamination in the first cell passage of the A. incisum isolate precluded successful isolation of the organism. PCR products were obtained with the gltA and htrA primers for the two isolates, but no product was obtained with the ompA primers. By BLAST analysis, partial gltA and htrA sequences of isolates from A. ovale and A. incisum were similar to the corresponding sequences of R. bellii. Conclusions. This is the first report of R. bellii infecting A. incisum and the first successful isolation from A. ovale.
Key words: Rickettsia bellii, Amblyomma ovale, Amblyomma incisum, Brasil.
RESUMEN
Objetivo. Aislar Rickettsias mediante cultivo celular a partir de muestras de garrapatas Amblyomma ovale y Amblyomma incisum del estado de São Paulo Materiales y métodos. A. ovale y A. incisum adultas de vida libre fueron colectadas en una área de selva tropical Atlántica en el estado de São Paulo, Brazil. Cada garrapata fue sometida a la prueba de hemolinfa, las garrapatas positivas en esta prueba fueron evaluadas con la técnica de shell vial con el propósito de aislar rickettsias en cultivo de células Vero. Pasajes celulares de los aislados fueron identificados genotípicamente por la reacción en cadena por la polimerasa (PCR) dirigidos a fragmentos de tres genes de rickettsias (gltA, htrA y ompA), seguido por secuenciación de ADN. Resultados. Un total de 388 A. incisum y 50 A. ovale fueron colectadas. Por la prueba de hemolinfa, únicamente una A. incisum y una A. ovale fueron positivas. Las Rickettsias fueron exitosamente aisladas de estas garrapatas. Sin embargo, el cultivo continuo en células Vero fue posible sólo para la garrapata A. ovale, debido a contaminación bacteriana en el primer pasaje celular de la muestra de A. incisum. Los productos de PCR fueron obtenidos con los primers gltA y htrA para los dos aislados, no obstante, ningún producto fue obtenido con los primers ompA. Por análisis BLAST, secuencias parciales de gltA y htrA procedentes de los aislados de A. ovale y A. incisum fueron similares a las secuencias correspondientes a R. bellii. Conclusiones. Este es el primer reporte de R. bellii infectando A. incisum y el primer establecimiento exitoso de un aislado de A. ovale.
Palabras clave: Rickettsia bellii, Amblyomma ovale, Amblyomma incisum, Brasil.
INTRODUCTION
Rickettsiae are pathogenic bacteria that require eukaryotic cells in order to grow. They are gram-negative, small, rod-shaped, a-proteobacteria causing mild to severe diseases in humans and animals. The genus subdivides into the conventionally well defined typhus group (TG) and spotted fever group (SFG) based on phylogenetic analyses. The TG includes the louse-borne Rickettsia prowazekii (the agent of epidemic typhus) and the flea-borne Ricketsia typhi (the agent of endemic typhus). The SFG includes tick-borne Rickettsia rickettsii (the agent of Rocky Mountain spotted fever), Rickettsia conorii (the agent of Mediterranean spotted fever), Rickettsia africae (the agent of African tick bite fever), Rickettsia sibirica (the agent of North Asian tick-borne fever), mite-borne Rickettsia akari (the agent of rickettsialpox), and flea-borne Rickettsia felis (the agent of flea-borne spotted fever) (1,2). Rickettsia bellii is the most divergent species, being representative of a distinct group within the genus Rickettsia, together with Rickettsia canadensis (3). R. bellii, the most common rickettsia found in ticks in America, was isolated for the first time in embryonated chicken eggs from a triturated pool of unfed adult Dermacentor variabilis in the United State, one of the most important vectors of R. rickettsii in that country, but there is no evidence that R. bellii can infect humans (4). It has been found in various other hard ticks, including species of Amblyomma, Ixodes, Haemaphysalis, and in soft ticks, including species of Argas and Ornithodoros (4-12).
In Brazil, despite R. bellii was also the most common rickettsial species identified in ticks from the state of Rondônia, Western Amazon, it was only successfully isolated and established in continuous cell passages from Amblyomma scalpturatum (7) and in Amblyomma aureolatum, Amblyomma dubitatum, and Haemaphysalis juxtakochi in the state of São Paulo (10-12). In the present study, we tested samples of Amblyomma ovale and Amblyomma incisum ticks collected in the state of São Paulo (southeastern Brazil), attempting to isolate rickettsiae in cell culture and establishment in continuous cell passages.
MATERIALS AND METHODS
During 2004-2006, adult free-living A. ovale and A. incisum ticks were collected on the vegetation in an Atlantic rainforest area in the Intervales State Park, Ribeirão Grande Municipality, state of São Paulo (24o18’S, 48o24’W) by visual inspection of vegetation during another study on ecology of free-living ticks in the Intervales State Park (13). Ticks were brought alive to the laboratory and before testing live ticks by the hemolymph test, ticks were held in an incubator at 32oC and 95% RH for 3 to 5 days, in order to stimulate rickettsial growth (14).
Thereafter, each tick was subjected to the hemolymph test for detection of Rickettsia-like structures, as previously described (14). Briefly, the distal portion of one leg of each tick was cut with scissors and a drop of hemolymph was fixed by air in a glass slide and stained by Gimenez staining (15). Right after the collection of hemolymph, ticks were frozen in a -80oC freezer.
Attempts to isolate Rickettsia in Vero cell culture were performed with ticks shown to contain Rickettsia-like structures by the hemolymph test. For this purpose, frozen ticks were thawed and subjected to the shell vial technique as previously described (16), with some modifications (8). Briefly, individual ticks were thawed in water bath at 37oC and disinfected for 10 minutes in iodine alcohol followed by several washes in sterile water. Then each tick was triturated in 500 ml of brain heart infusion broth (BHI) and the resultant tick homogenate inoculated into shell vials containing a monolayer of confluent Vero cells. After inoculation, the shell vials were centrifuged for one hour at 700 g at 22oC. Thereafter, the monolayer was washed once with minimal essential medium containing 5% bovine calf serum, and then incubated at 28oC with medium containing antibiotics (1% penicillin, streptomycin). After three days, the medium was switched to antibiotic-free medium, and the aspirated medium was checked by Gimenez staining for the presence of Rickettsia like-organisms. If the result was positive, the monolayer of the shell vial was harvested and inoculated into a 25 cm2 flask containing a monolayer of confluent uninfected Vero cells. Cells of the 25 cm2 flask were checked by Gimenez staining until more than 90% of the cells were infected, when they were harvested and inoculated into 150 cm2 flasks of Vero cells. At all instances, inoculated Vero cells were incubated at 28oC. The level of infection of cells was monitored by Gimenez staining (15), of scrapped cells from inoculated monolayer. A rickettsia isolate was considered established in the laboratory after at least three passages through 150 cm2 Vero cell flasks, each reaching more than 90% of infected cells (8).
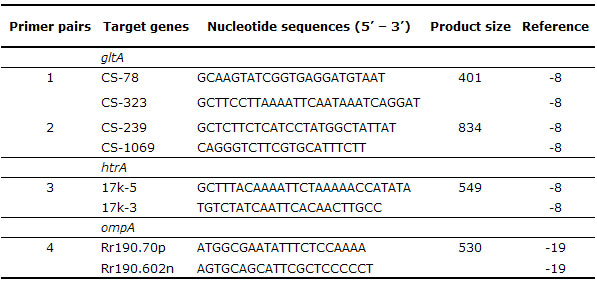
Cell passages of isolates were genotypicaly identified by polymerase chain reaction (PCR) amplification and sequencing the product of the resultant infected cells. For this purpose, DNA was extracted using the Dneasy Tissue Kit (Quiagen®), following manufacturer’s protocol for isolation of DNA from culture cells samples and DNA from infected cell passages were tested by a battery of PCR using all primer pairs listed in table 1, targeting fragments of tree rickettsial genes: citrate synthase (gltA), 17-kDa outer membrane protein (htrA), and 190-kDa outer membrane protein (ompA). For each set of reactions, a negative control (5 µl of water) and positive control (5 µl of DNA extracted from A. cajennense ticks experimentally infected with R. parkeri) was included (17).
Table 1. Primer pairs used for amplification of rickettsial genes.
All PCR products of the expected size obtained in the present study were purified using ExoSap (USB) and sequenced in an automatic sequencer (Applied Biosystens/Perkin Elmer, model ABI Prism 310 Genetic, California, USA) according to the manufacturer’s protocol. Partial sequences obtained were submitted to BLAST analysis, (18), to determine similarities to other Rickettsia species.
RESULTS
A total of 388 ticks A. incisum were collected (180 males, 208 females) and 50 ticks A. ovale (17 males, 33 females). By the hemolymph test, one A. incisum male and one A. ovale female that showed to contain rickettsia-like organisms by the hemolymph test were processed by the shell vial technique (8,16). Rickettsia was successfully isolated and established in Vero cell culture from tick A. ovale. Although the establishment of Rickettsia in Vero cell culture from the A. incisum tick was unsuccessful, the initial cultivation of rickettsia was possible due to the presence of numerous Giménez-stained intracellular rickettsia-like organisms from the inoculated shell vials, which could not be cultivated further than the first passage owing to contamination of the cell cultured by extracellular bacteria.
Even though, DNA of contaminated cell culture at first passage was tested by PCR, as described below. The isolate from A. ovale generated in the present study has been deposited in the Rickettsial Collection of the Laboratory of Parasitic Diseases of the Faculty of Veterinary Medicine, in the University of São Paulo, where it is available upon request.
DNA of infected cells at fourth passage of A. ovale and first passage of A. incisum were subjected to PCR targeting the rickettsial genes gltA, ompA and htrA. PCR products of the expected size were obtained with the gltA and htrA primers for two isolates, but no product was obtained with the ompA primers (these primers have been specific for SFG rickettsiae). We sequenced 1060 and 500 nucleotides of the gltA and htrA genes, respectively, from the A. ovale isolate, and 1096 and 499 nucleotides of the gltA and htrA genes, respectively, from the A. incisum isolate. By BLAST analysis, partial gltA sequence of the A. ovale rickettsia in the present study was 100% (1060/1060) similar to the corresponding sequence of R. bellii strain HJ7 (DQ865204), and partial gltA sequence of the A. incisum rickettsia was 99.9% (1095/1096) similar to the corresponding sequence of Rickettsia bellii RML369-C (CP000087). Partial htrA sequence of the A. ovale rickettsia was 99.6% (497/499) similar to the corresponding sequence of R. bellii (AY362702), whereas partial htrA sequence of the A. inicisum rickettsia was 100% (499/499) similar to R. bellii (AY362702). Thus, both rickettsial agents from A. ovale and A. inicisum were genotypically identified as R. bellii.
DISCUSSION
R. bellii has been reported in 18 tick species belonging to the genera Amblyomma (8 species), Dermacentor (5 species), Haemaphysalis (2 species), Ixodes (1 species), Argas (1 species), and Ornithodoros (1 species) from different parts of Brazil, Argentina and United States, (4-12). Previously in Brazil, this bacterium was reported infecting A. ovale from Rondônia (northern Brazil), besides the ticks A. dubitatum, A. aureolatum, Amblyomma oblongoguttatum, A. scalpturatum, Amblyomma humerale, Amblyomma rotundatum, Amblyomma nodosum, Ixodes loricatus, and H. juxtakochi, (6-12). Among the genus Rickettsia, R. bellii is indeed the species with the greatest number of tick records. The present study reports for the first time R. bellii infecting the tick A. incisum, adding one more species for the list of ticks that are naturally found infected with this rickettsia in the New World. We also provide the first successful in vitro culture of R. bellii from A. ovale.
Although infection rates by R. bellii in tick populations are high variable (3.6 to 100%), but usually high (5,7,8), there has been no direct or indirect evidence of human infection by R. bellii. Among animals, there is only one study showing serological evidence of R. bellii infection in capybaras from several parts of the state of São Paulo, possibly transmitted by A. dubitatum ticks, (20).
A. incisum and A. ovale are ticks of Neotropical origin, (21) and while conducting projects on ticks from deer and on tick ecology in animal trails in this same Atlantic rainforest reserve of the present study, researchers were bitten by ticks numerous times, with most of the reports caused by A. incisum, (22), which was the must abundant tick species along the trails (23).
The occurrence of human infestation by A. ovale was also reported in the study area, (22), beyond being the most frequent human-biting tick in the state of Rondônia, western Brazilian Amazon, (24). Thus, it is very important to know the Rickettsia species that are infecting these important human-biting ticks in South America. In addition, less pathogenic rickettsiae could have important ecological implications if one considers that the presence of less pathogenic rickettsiae within a tick population can minimize the transmission of a greater pathogenic rickettsia. (25)
Acknowledgments
We thank Pilar Vejarano for translating the abstract to Spanish. This work was supported by FAPESP (grant 03/13872-4 to M.B.L. and scholarship 03/13871-8 to R.C.P.) and CNPq (Academic Career Research Fellowship to M.B.L. and M.P.J.S.).
REFERENCES
1. Raoult D, Roux V. Rickettsioses as paradigms of new or emerging infectious diseases. Clin Microbiol Rev 1997; 10: 694-719. [ Links ]
2. Ogata H, La Scola B, Audic S, Renesto P, Blanc G, Robert C, et al. Genome Sequence of Rickettsia bellii Illuminates the Role of Amoebae in Gene Exchanges between Intracellular Pathogens. PLoS Genet 2006; 2: 733-744. [ Links ]
3. Stothard DR, Clark JB, Fuerst PA. Ancestral divergence of Rickettsia bellii from the spotted fever and typhus groups of Rickettsia and antiquity of the genus Rickettsia. Int J Syst Bacteriol 1994; 44: 798-804. [ Links ]
4. Philip RN, Casper EA, Anacker RL, Cory J, Hayes SF, Burgdorfer W, et al. Rickettsia bellii sp. nov.: a tick-borne Rickettsiae, widely distributed in the United States, that is distinct from the spotted fever and typhus biogroups. Int J Syst Bacteriol 1983; 33: 94-106. [ Links ]
5. Labruna MB, Pacheco RC, Nava S, Brandão PE, Richtzenhain LJ, Guglielmone AA. Rickettsia bellii and Rickettsia amblyommii infection in Amblyomma neumanni ticks from Argentina. Microb Ecol 2007; 54: 126-133. [ Links ]
6. Horta MC, Pinter A, Schumaker TTS, Labruna MB. Natural infection, transovarial transmission, and transstadial survival of Rickettsia bellii in the tick Ixodes loricatus (Acari: Ixodidae) from Brazil. Ann N Y Acad Sci 2006; 1078: 285-290. [ Links ]
7. Labruna MB, Whitworth T, Bouyer DH, McBride JW, Camargo LMA, Camargo EP, et al. Rickettsia bellii and Rickettsia amblyommii in Amblyomma ticks from the state of Rondônia, Western Amazon, Brazil. J Med Entomol 2004; 41: 1073-1081. [ Links ]
8. Labruna MB, Whitworth T, Horta MC, Bouyer DH, McBride JW, Pinter A, et al. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of São Paulo, Brazil, where Brazilian spotted fever is endemic. J Clin Microbiol 2004; 42: 90-98. [ Links ]
9. Ogrzewalska M, Pacheco RC, Uezu, A, Ferreira F, Labruna MB. Rickettsia infection in Amblyomma nodosum ticks from Brazil. En:Congreso; 5th International Meeting on Rickettsiae and Rickettsial Diseases. Brazil: Program Abstract Book 2008. [ Links ]
10. Pinter A, Labruna MB. Isolation of Rickettsia rickettsii and Rickettsia bellii in cell culture from the tick Amblyomma aureolatum in Brazil. Ann N Y Acad Sci 2006; 1078: 523-529. [ Links ]
11. Pacheco RC, Horta MC, Pinter A, Moraes-Filho J, Martins TF, Nardi MS, et al. Pesquisa de Rickettsia spp em carrapatos Amblyomma cajennense e Amblyomma dubitatum no Estado de São Paulo, Brasil. Rev Soc Bras Med Trop in press 2008. [ Links ]
12. Labruna MB, Pacheco RC, Richtzenhain LJ, Szabó MPJ. Isolation of Rickettsia rhipicephali and Rickettsia bellii from ticks Haemaphysalis juxtakochi in the state of São Paulo, Brazil. Appl Environ Microbiol 2007; 73: 869-873. [ Links ]
13. Szabó MPJ, Labruna MB, Garcia MV, Pinter A, Castagnolli KC, Pacheco RC el al. Ecological aspects of free-living ticks (Acari: Ixodidae) on animal trails in an Atlantic rainforest of Southeastern Brazil Ann Trop Med Parasitol in press 2008. [ Links ]
14. Burgdorfer W. The hemolymph test. Am J Trop Med Hyg 1970; 19: 1010-1014. [ Links ]
15. Giménez DF. Staining rickettsiae in yolk-sac cultures. Stain Technol 1964; 39: 35-140. [ Links ]
16. Kelly JP, Raoult D, Mason PR. Isolation of spotted fever group rickettsiae from triturated ticks using a modification of the centrifugation vial technique. Trans R Soc Trop Med Hyg 1991; 85: 397-398. [ Links ]
17. Sangioni LA, Horta MC, Vianna MCB, Gennari SM, Soares RS, Galvão MAM, Schumaker TTS, Ferreira F, Vidotto O, Labruna MB. Rickettsial infection in animals and Brazilian spotted fever endemicity. Emerg Infect Dis 2005; 11: 265-270. [ Links ]
18. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol 1990; 215: 403-410. [ Links ]
19. Regnery RL, Spruill CL, Plikaytis BD. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J Bacteriol 1991; 173: 1576-1589. [ Links ]
20. Pacheco RC, Horta MC, Moraes-Filho J, Ataliba AC, Pinter A, Labruna MB. Rickettsial infection in capybaras (Hydrochoerus hydrochaeris) from São Paulo, Brazil: serological evidence for infection by Rickettsia bellii and Rickettsia parkeri. Biomédica 2007; 27: 364-371. [ Links ]
21. Guglielmone AA, Estrada-Penã A, Keirans JE, Robbins RG. Ticks (Acari: Ixodida) of the zoogeographic region. International Consortium on Ticks and Tick-Borne Diseases (ICTTD-2), The Netherlands, Atalanta, Houten, 2003. [ Links ]
22. Szabó MPJ, Labruna MB, Castagnolli KC, Garcia MV, Pinter A, Veronez VA et al. Ticks (Acari: Ixodidae) parasitizing humans in an Atlantic rainforest reserve of Southeastern Brazil with notes on host suitability. Exp Appl Acarol 2006; 39: 339-346. [ Links ]
23. Szabó MPJ, Labruna, MB, Vogliotti A, Duarte JMB. Ticks (Acari: Ixodidae) on Small Red Brocket Deer (Mazama bororo Duarte) along Deer Trails in the Atlantic Rain Forest of Southeastern Brazil. Syst Appl Acarol 2006; 11: 41-45. [ Links ]
24. Labruna MB, Camargo LM, Terrassini FA, Ferreira F, Schumaker TS, Camargo EP. Ticks (Acari: Ixodidae) from the state of Rondônia western Amazon, Brazil. Syst Appl Acarol 2005; 10: 17-32. [ Links ]
25. Macaluso KR, Sonenshine DE, Ceraul SM, Azad AF. Rickettsial infection in Dermacentor variabilis (Acari: Ixodidae) inhibits transovarial transmission of a second rickettsia. J Med Entomol 2002; 39: 808-813. [ Links ]

