










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista MVZ Córdoba
Print version ISSN 0122-0268On-line version ISSN 1909-0544
Rev.MVZ Cordoba vol.13 no.2 Córdoba May/Aug. 2008
CRECIMIENTO DE Brachiaria decumbens Stapf Y Cynodon nlemfuensis Vanderyst EN SUELOS SULFATADOS ÁCIDOS DE CÓRDOBA
GROWTH OF Brachiaria decumbens Stapf AND Cynodon nlemfuensis Vanderyst IN ACID SULPHATE SOILS OF CORDOBA
Enrique Combatt C,1* M.Sc, Alfredo Jarma O,1 M.Sc, Libardo Maza A.2. M.Sc.
1Universidad de Córdoba, Facultad de Ciencias Agrícolas. 2 Universidad de Córdoba, Facultad de Medicina Veterinaria y Zootecnia. Departamento de Ciencias Pecuarias *Correspondencia: ecombatt@sinu.unicordoba.edu.co
Recibido: Marzo 10 de 2008; Aceptado: Julio 18 de 2008.
RESUMEN
Objetivo. Evaluar el efecto químico de dosis de cal en suelos sulfatados ácidos (SSA) y las respuestas fisiológicas de Cynodon nlemfuensis Vanderyst y Brachiaria decumbens Stapf. Materiales y métodos. El trabajo se desarrolló durante 2006 en suelos sulfatados ácidos de la finca “El Deseo” (Ciénaga de Oro - Córdoba), donde se aplicaron 3, 6 y 9 toneladas de cal agrícola por hectárea y se establecieron dos especies de pasto, Cynodon nlemfuensis Vanderyst (pasto estrella) y Brachiaria decumbens Stapf (pasto brachiaria). Las respuestas consideradas fueron las propiedades químicas del suelo, peso fresco y seco de genotipos y relación hoja/tallo. Se diseñó un experimento de bloques completamente randomizados con estructura de parcelas divididas, donde las parcelas principales correspondieron a los pastos y las subparcelas a las dosis de encalamiento. Resultados. Los valores del pH del suelo aumentaron y las concentraciones de aluminio decrecieron significativamente con la dosis de cal reduciéndose hasta en un 80%. Las bases intercambiables aumentaron con la aplicación de cal y los microelementos disminuyeron en función del tiempo, aunque no con el encalamiento. Por su parte, independientemente del genotipo, la producción de biomasa fresca mostró una relación lineal con el nivel de encalamiento (y=0.656x + 11.99; r2=0.92); la materia seca y la relación hoja/tallo fueron mayores siempre para B. decumbens respecto a C. nlemfuensis, de manera independiente al nivel de encalamiento. Conclusiones. La aplicación de dosis crecientes de cal en SSA incrementó los valores de pH y la producción de biomasa en la pastura independientemente de la especie forrajera.
Palabras clave: Acidez, suelo, pastos, forrajes, brachiaria, trópico.
ABSTRACT
Objective. To assess the effect of different doses of lime in acid sulphate soils and physiological responses of two species of grasses, Cynodon nlemfuensis Vanderyst and Brachiaria decumbens Stapf. Materials and methods. The work was carried out during 2006 in acid sulfate soils at El Deseo farm (Ciénaga de Oro - Córdoba, Colombia), where 3, 6 and 9 tons/ha of agricultural lime was applied and two grass species, Cynodon nlemfuensis Vanderyst (pasto estrella) and Brachiaria decumbens Stapf (pasto brachiaria) were planted. The responses considered during 5 mowings were the chemical features of the soil, fresh and dry weight of each genotype and the leaf/stem ratio. An experiment of complete random blocks was designed using a structure of divided plots, where the main plots corresponded to the grasses and the subplots to the lime doses. Results. The most outstanding results suggested that the pH values and aluminum in the soil concentrations fall significantly with the quantity of applied lime, decreasing up to 80%. In general, the interchangeable bases increased with the lime dosage and the contents of the microelements decreased considerably over time, although this was not evident for the lime. Independently of genotype, the production of fresh biomass showed a linear relationship within lime dosage (y=0,656x + 11,99; R2=0.92); the dry mass and the leaf/stem ratio were always greater for B. decumbens compared to C. nlemfuensis, independent of lime dosage.
Key words: Acidity, soil, grasses, forages, Brachiaria, tropic.
INTRODUCCIÓN
Para el sector agropecuario, los suelos sulfatados ácidos (SSA) son los más limitados del mundo debido a que generan ácido sulfúrico, el cual contribuye a que su pH sea más bajo que 3. En este ambiente ácido, el aluminio y otros elementos tóxicos afectan la vegetación, la vida acuática y hacen que muchas especies se atrofien y enfermen. La fuerte acidificación de los suelos sulfatados ácidos es debida a la oxidación de la pirita (Fe2S), con la consiguiente producción de ácido sulfúrico (H2SO4) (1). Cuando la oxidación progresa, la acidificación influye en el pH y aumenta la solubilidad del aluminio (AI3+), manganeso (Mn2+) y el hierro (Fe2+). Los ácidos movilizan hierro, aluminio, manganeso y otros metales en forma soluble en el agua, en concentraciones que generalmente son fitotóxicas para las plantas y la vida acuática (2). Además, pueden resultar en impactos ecológicos importantes, si posteriormente son transportados o liberados al agua (3). A pH<4.5, el aluminio predomina como ion monomérico (Al3+) y puede ser liberado por arcillas minerales y la materia orgánica en descomposición, representando una fuente de acidez además del hidrógeno exportado.
La toxicidad por aluminio limita en un 30% la producción de los cultivos en los suelos arables del mundo que incluyen muchas áreas del trópico y subtrópico, donde predomina el aluminio en forma de Al3+, lo cual afecta el crecimiento de las raíces y aumenta la deficiencia de nutrientes (4). Kochian (5), informó que las estrategias para aumentar la producción en suelos ácidos, incluyen la aplicación de cal y el uso de plantas tolerantes a la acidez, ya que la aplicación de calcio o carbonato de calcio reduce el estrés causado por el aluminio, al ser neutralizado mediante la precipitación o la insolubilización con los hidroxilos producidos en la disociación del carbonato de calcio y el agua. En este sentido Ward et al (6), afirmaron que una de las técnicas de manejo comúnmente empleadas para la neutralización de la acidez de los suelos sulfatados ácidos potenciales y/o actuales, son los materiales alcalinos tales como la cal.
Por otra parte, el desarrollo de muchos genotipos de pastos tolerantes al aluminio ha contribuido grandemente al incremento de la productividad de su biomasa en suelos ácidos. Asímismo, la genética de las especies tolerantes al aluminio ha sido examinada últimamente en muchas instancias y sugiere que esta tolerancia es diversa entre variedades y que puede estar bajo el control de diferentes genes (7). Los mecanismos de esta adaptación, aunque parcialmente desconocidos, pueden ser por una parte la exclusión del aluminio y por otra la inactivación interna. Algunos autores indican que los mecanismos de exclusión del aluminio en algunas especies, puede no estar completamente identificada aunque es conocido que muchas de ellas tienen mecanismos eficientes para la exudación de este ión a través de ácidos orgánicos por las raíces (8,9).
Otros mecanismos de resistencia al aluminio también han sido reportados, como el incremento del pH en la rizosfera y la formación de complejos de aluminio con ácidos orgánicos o quelatos, que secuestran esos complejos en las vacuolas y juegan un papel importante en la detoxificación por parte de las plantas. En este sentido es importante considerar la alta efectividad de resistencia a la toxicidad por aluminio de Brachiaria decumbens Stapf, que secreta ácidos orgánicos y fosfato en los ápices de las raíces mediante diferentes mecanismos fisiológicos y acumula altas concentraciones de aluminio en las raíces acompañada por la estimulación de síntesis de malato (10). Marschner (11), indicó que la capacidad de detoxificación por parte de los ácidos orgánicos como el malato, depende de la relación de sus grupos hidroxilos/carboxilos (OH/COOH), y a estos grupos corresponde la posibilidad de formar una estructura estable de 5 ó 6 enlaces con el aluminio. A estas características de B. decumbens se atribuye su excelente adaptación a suelos infértiles ácidos, siendo la especie más cultivada del género Brachiaria (12,13).
Considerando lo anterior, y en razón a las áreas crecientes de SSA en el Caribe colombiano, se planteó la investigación con el objetivo de evaluar el efecto de diferentes dosis de cal a suelos sulfatados ácidos y las respuestas fisiológicas de dos especies de pastos Cynodon nlemfuensis Vanderyst y Brachiaria decumbens Stapf.
MATERIALES Y MÉTODOS
La investigación se realizó en la finca “El Deseo”, ubicada en el bajo Sinú, corregimiento Caño Bugre, municipio de Ciénaga de Oro, departamento de Córdoba.
Se tomaron muestras de suelo, con el propósito de verificar en el laboratorio las características que identifican un SSA (pH<5, M.O.>4%, Al2+>2 cmolc.kg-1 y azufre intercambiable >500 mg/Kg ppm en horizontes diagnóstico). Una vez determinadas las áreas afectadas por suelos sulfatados ácidos, se seleccionó la finca “El Deseo”, en donde se excavó una calicata de 1x1x1 m para su descripción taxonómica y colecta de las muestras de cada horizonte. El suelo se caracterizó mediante análisis químico según las normas establecidas por el IGAC y aprobadas por el Control Analítico de Laboratorios de Suelos de la Sociedad Colombiana de la Ciencia del Suelo (CALS), implementadas por el Laboratorio de Suelos y Aguas de la Universidad de Córdoba. Los parámetros químicos que se determinaron fueron: pH, materia orgánica, azufre, fósforo, calcio, magnesio, sodio, potasio, aluminio, capacidad de intercambio catiónico y los elementos menores hierro, manganeso, zinc y cobre.
Diseño experimental. Los trabajos se realizaron bajo un diseño experimental de bloques al azar con estructura de parcelas divididas, las parcelas principales correspondieron a los genotipos de pasto Cynodon nlemfuensis Vanderyst (pasto estrella) y Brachiaria decumbens Stapf (pasto brachiaria), en tanto que las subparcelas estuvieron conformadas por las dosis de cal agrícola (3, 6 y 9 ton.ha-1).
La preparación del terreno fue la convencional de la zona (2 pases de rastra pesada más 1 pase de rastrillo pulidor) y el área del experimento tuvo las siguientes dimensiones: 30 x 60 m como área total, dividida en cuatro bloques de 5 x 40 m cada uno separados entre sí por 2 m; cada bloque constituyó dos parcelas principales de 5 x 20 m cada una (genotipos) y cada parcela principal estuvo conformada por 4 subparcelas de 5 x 5 m donde se aplicaron las dosis de cal.
Luego de realizar los tratamientos de encalado en cada unidad experimental, se esperaron 30 días para la siembra de los genotipos de pasto con el propósito de que el carbonato de calcio se hidrolizara y se liberaran iones OH y Ca2+. Los materiales genéticos se establecieron con una distancia de siembra de 50 por 50 cm. La fertilización con nitrógeno, fósforo y potasio se realizó de acuerdo con los resultados del análisis químico para cada uno de estos elementos, en cantidades por hectárea como se relacionan a continuación: el fósforo en presiembra incorporado a razón de 46 Kg como fosfato diamónico, el nitrógeno como urea en dosis de 92 Kg y el potasio como KCl en razón de 60 Kg. Las aplicaciones se dividieron en cuatro épocas, de manera sincronizada con los cortes de las pasturas.
A los 90 días de encalado y al final del cultivo (150 días después de encalado), se tomaron muestras de 1 kg de suelo por tratamiento para determinar nuevamente su composición química y analizar su salinidad.
Como variables dependientes se consideraron las características químicas del suelo en las subparcelas, así como algunas características del crecimiento y la morfofisiología de la planta tales como el peso húmedo y peso seco en gramos y la relación biomasa de hojas/biomasa de tallos; las muestras se tomaron cada 30 días después de la siembra en un tamaño correspondiente a cuatro submuestras por tratamiento. A esto se le denominó época de corte que correspondieron a los 30, 60, 90, 120 y 150 días después de siembra.
Como variables independientes se consideraron los genotipos y los tratamientos de encalado, como se relaciona a continuación:
To = testigo (sin tratamiento de encalado)
T1 = 3 toneladas de cal agrícola por hectárea
T2 = 6 toneladas de cal agrícola por hectárea
T3 = 9 toneladas de cal agrícola por hectárea.
Los resultados se analizaron mediante el paquete estadístico Statistical Analysis System (SAS). Cuando se presentaron interacciones se llevaron a cabo comparaciones horizontales y ajustes de curvas de respuesta mediante polinomios ortogonales para cada especie; cuando no se detectaron interacciones significativas se realizaron pruebas de comparación de medias para las especies y ajustes de curvas de tendencia para las dosis.
RESULTADOS
En la tabla 1 se muestran los resultados del análisis químico de los suelos del estudio antes (To) y después del encalado para cada tratamiento (T1, T2 y T3), tanto a los 90 como a los 150 días después de realizar el encalamiento (DDE), lo que correspondió respectivamente a los 60 y 120 días después de siembra. Las condiciones iniciales del suelo indicaron que el pH es fuertemente ácido, la materia orgánica se consideró alta, el azufre es excesivo y el fósforo medio. El calcio, magnesio y potasio tienen niveles altos y el sodio a los niveles observados, no causaría problemas. La concentración de aluminio es excesiva y todos los microelementos tienen concentraciones consideradas altas.
Tabla 1. Caracterización química de suelos sulfatados ácidos en Ciénaga de Oro - Córdoba a los 90 y 150 días después de tres niveles de encalamiento.
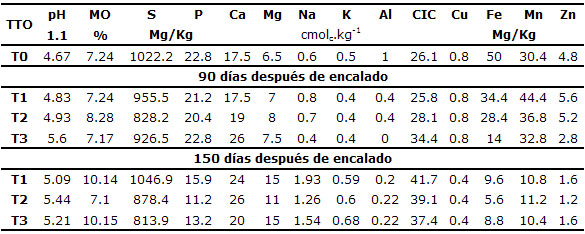
T0=Testigo absoluto sin encalar; T1, T2 y T3=Tratamientos con 3, 6 y 9 toneladas de cal por hectárea.
A los 90 días después de encalado, la reacción del suelo fue ácida en casi todos los tratamientos, la materia orgánica siguió siendo alta, el azufre disminuyó pero de igual forma hubo concentraciones excesivas y el fósforo no mostró cambios en sus niveles. Las concentraciones de calcio y magnesio aumentaron con las dosis de cal aplicada y para el potasio redujo su concentración. El sodio no mostró cambios y para el aluminio se observó una disminución importante. Los microelementos mostraron una leve tendencia a disminuir en sus niveles.
A los 150 días después de encalado, la reacción del suelo fue ácida a ligeramente ácida, siguiendo una tendencia creciente en sus valores absolutos en la medida que aumentó el nivel de encalamiento y el tiempo de aplicación de la cal; la materia orgánica siguió siendo alta con incrementos importantes en los niveles de 3 y 9 toneladas de cal/ha. El azufre, a pesar de registrar una tendencia decreciente con el aumento en los niveles de encalamiento, principalmente con la aplicación de 6 y 9 ton de cal/ha, continuó aún en niveles excesivos y el fósforo mostró un comportamiento similar al azufre aunque sus niveles podrían considerarse como relativamente bajos.
Respecto a las bases intercambiables, las concentraciones de calcio aumentaron, de los 90 a los 150 DDE, en un 36% en los tratamientos de 3 y 6 toneladas de cal por hectárea, en tanto que al aplicar 9 toneladas, la concentración de este elemento se redujo en un 24% en el mismo lapso de tiempo. El magnesio registró la misma tendencia, en el sentido de aumentar con el nivel de cal aplicada y el tiempo, aunque el incremento en los tratamientos de 3 y 9 ton de cal por hectárea fue mayor que el de 6 toneladas, pasando de 7 y 7,5 cmolc.kg-1 de suelo respectivamente, a 15 (un incremento porcentual de 114%), en tanto que en el tratamiento de 6 toneladas de cal/ha, los valores de magnesio pasaron de 8 a 11 cmolc.kg-1 de suelo, lo que indicó un incremento del 38%. Es importante resaltar que las concentraciones de este elemento pueden considerarse como altas. Por su parte, el potasio tendió a aumentar levemente entre los 90 y 150 días después del encalado, aunque no parece que hubo un efecto importante de la dosis de cal en ninguna de estas dos épocas. En este mismo período de tiempo, el sodio mostró una tendencia a aumentar en proporción de 1.4, 0.8 y 2.8 veces para los tratamientos de 3, 6 y 9 toneladas de cal/ha respectivamente.
Al considerar los niveles de aluminio en el suelo antes de aplicar las enmiendas (cmolc.kg-1 de suelo) respecto a cualquiera de los tratamientos a los 90 y 150 DDE, se observó claramente una reducción importante, siendo lo más significativo que a los 150 DDE, cualquiera de los tratamientos de encalado, redujo en 80% la concentración de éste ion respecto a sus valores iniciales.
Por otra parte, en términos generales se observa que los contenidos de los microelementos disminuyeron considerablemente en función del tiempo desde el encalamiento, aunque esta tendencia no fue tan clara cuando se analizó en función de la dosis de cal aplicada. El cobre, de manera independiente al grado de encalamiento se redujo a la mitad de su concentración, y este efecto fue evidente a los 150 DDE. El hierro mostró una reducción progresiva desde su concentración inicial (50 mg/kg) a un promedio de 25.6 y 8 mg/kg a los 90 y 150 DDE respectivamente. Las concentraciones de manganeso se vieron afectadas solamente a los 150 DDE, reduciéndose de 30.4 mg/kg en el suelo sin tratar, hasta niveles promedio de 10.8 mg/kg en los tres tratamientos de enmienda. Por último, los niveles de zinc se redujeron aproximadamente a una tercera parte al pasar de 4.8 mg/kg en el testigo absoluto a 1.5 mg/kg a los 150 DDE.
Análisis fisiológico de los pastos. En la tabla 2 se muestran los cuadrados medios de las variables fisiológicas consideradas en el estudio con su respectiva significancia estadística. Es importante resaltar que para las tres variables se observó una alta significancia para el efecto de la interacción época por genotipo, así como los efectos individuales de cada uno de estos factores. Las interacciones época por dosis y genotipo por dosis no mostraron, en general, una significancia importante.
Tabla 2. Cuadrados medios de las variables fisiólogicas consideradas.
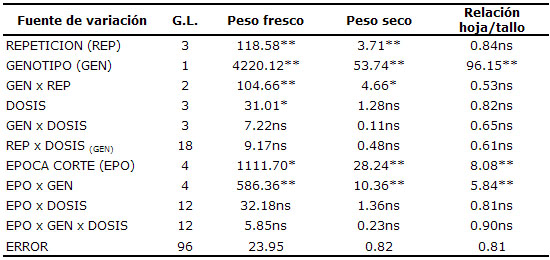
* y ** = Diferencias significativas (p<0.05) y altamente significativas (p<0.01) respectivamente; ns = no hay diferencias estadísticas (prueba de Tuckey: 5%).
Peso fresco. Los resultados estadísticos indicaron que solamente se presentaron diferencias importantes del peso fresco para las dosis y la interacción época por genotipo. La figura 1 muestra la descomposición de la interacción mencionada y evidencia que está explicada por el mejor comportamiento del pasto Brachiaria en condiciones de alta disponibilidad de agua (las épocas 30, 60 y 90 DDE fueron de mayor precipitación aunque los datos no son mostrados en el artículo), expresado en producción de masa fresca por hectárea. Estas diferencias no son tan evidentes, sino que más bien sugieren una tendencia a un comportamiento similar cuando la disponibilidad de agua no es tan alta como se observa a los 120 y 150 DDE.
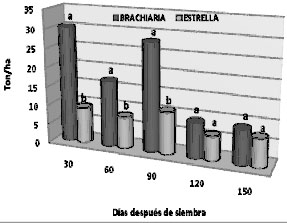
Figura 1. Análisis de la interacción época de corte x genotipo para el peso fresco de dos genotipos de pasto en cincoépocas de corte en suelos sulfatados ácidos de Ciénaga de Oro - Córdoba. (Promedios dentro de una misma época con la misma letra, no difieren estadísticamente de acuerdo a la prueba de Tuckey a un nivel del 5% de probabilidad).
La figura 1 evidencia además que, en términos generales, los rendimientos de biomasa de pasto Brachiaria tendieron a decrecer con el tiempo en tanto que los de pasto estrella se mantuvieron a ser más o menos estables. Sin embargo, los rendimientos de biomasa totales de Brachiaria, al considerar las cinco épocas de corte, superaron en más de 200 toneladas por hectárea a los de pasto estrella (375 y 170 Ton/ha respectivamente).
Por otra parte, al estudiar el efecto de las dosis de cal sobre el peso fresco de los genotipos, se pudo determinar que existió una fuerte correlación entre esta variable y las dosis de cal aplicadas, de manera independiente al tipo de pasto. La biomasa por hectárea de los genotipos fue significativamente mayor al aplicar la mayor cantidad de CaCO3/ha, en una relación explicada de manera lineal y directamente proporcional por la ecuación Y = 0.656X + 11.99 (R2 = 0,925), donde Y es el rendimiento de biomasa (Ton/ha) y X las toneladas de cal/ha. Los rendimientos de biomasa pueden pasar de 12.7 a 14.8 Ton/ha, en un suelo sulfatado ácido sin tratamiento respecto a uno tratado con 9 toneladas de cal por hectárea, lo que sugiere una ganancia hipotética de 230 kilos/ha de pasto fresco por cada tonelada de cal aplicada.
La figura 2 muestra la integración de las respuestas de cada genotipo en función de la dosis de cal aplicada y la época de corte. La primera época de corte (30 DDE), independientemente de la dosis de encalamiento, presentó el mayor peso fresco en ambos genotipos y dentro de esta, la época. Este registro fue mayor cuando se aplicaron 9 ton de CaCO3/ha, con una ganancia de 37.74 Ton/ha para el pasto brachiaria y de 13.73 Ton/ha para pasto estrella. Para esta misma época el menor peso fue de 24.39 y 7.21 Ton/ha para Brachiaria y estrella respectivamente, en el tratamiento sin cal. Los rendimientos más bajos en peso fresco para ambos genotipos, se observaron a los 120 y 150 DDE, con valores que oscilaron entre 5.7 y 9.7 Ton/ha.
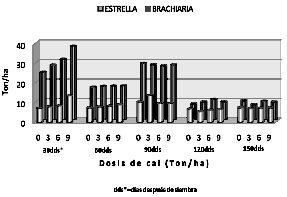
Figura 2. Peso fresco de dos genotipos de pasto bajo diferentes dosis de encalamiento y cinco épocas de corte en suelos sulfatados ácidos de Ciénaga de Oro - Córdoba
Peso seco. Tal como se mencionó anteriormente, el análisis de varianza para esta variable indicó que existió un efecto importante de la interacción época por genotipo. La figura 3, registra el análisis gráfico de la interacción mencionada cuya tendencia fue similar a lo observado para el peso fresco de esta localidad. En las primeras etapas de evaluación, los rendimientos de peso seco fueron más altos que en las épocas donde las deficiencias de agua fueron evidentes (120 y 150 DDE).
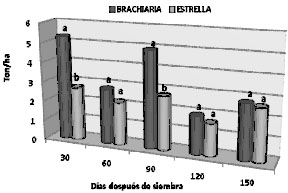
Figura 3. Análisis de la interacción época de corte x genotipo para el peso seco de dos genotipos de pasto en cincoépocas de corte en suelos sulfatados ácidos de Ciénaga de Oro - Córdoba (Promedios dentro de una misma época con la misma letra, no difieren estadísticamente de acuerdo a la prueba de Tuckey a
un nivel del 5% de probabilidad).
Durante las épocas de mayor disponibilidad de agua (30 y 90 DDE), los rendimientos de materia seca por hectárea de pasto brachiaria superaron en 2.6 y 2.2 toneladas respectivamente, en tanto que en la época de baja disponibilidad (120 y 150 DDE), estas ventajas sólo alcanzaron a 0.29 y 0.14 toneladas respectivamente. La producción acumulada de masa seca para cada genotipo independientemente del nivel de encalamiento indicó que, de manera consistente con lo observado para el peso fresco, el rendimiento total de biomasa seca del pasto brachiaria es superior al de estrella en 5.8 toneladas (17.7 y 11.9 Ton/ha respectivamente).
Relación hoja/tallo. La significancia estadística observada para el efecto de los factores evaluados sobre la relación hoja/tallo fue evidente para la interacción época por genotipo. De manera consistente con lo discutido para el peso húmedo y el peso seco, se encontró que la relación fue altamente significativa a favor de Brachiaria decumbens en la mayoría de las épocas de corte evaluadas, tal como se registra en la figura 4.
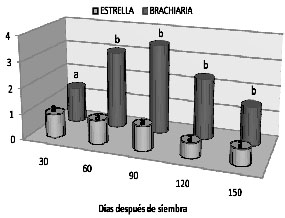
Figura 4. Análisis de la interacción para la relación hoja/tallo en cinco épocas de corte de dos genotipos de pastos en suelos sulfatados ácidos en Cienaga de Oro - Córdoba
A los 30 DDE, la distribución de asimilados que hacen los dos pastos hacia la hoja de manera preferencial respecto al tallo, fueron similares estadísticamente y esa proporción es del orden de 1.15. Sin embargo, las relaciones varíaron posteriormente con el genotipo y la época, llegando a registrarse proporciones de hasta 3.47 para pasto Brachiaria en el tercer corte (90 DDE), momento a partir del cual esta relación comenzó a decrecer. Por su parte, esta fracción siempre estuvo por debajo de 1 en pasto estrella, indicando que éste genotipo en cualquier edad, dentro del rango observado y considerando solamente la época de corte, envió más fotoasimilados al tallo que a la hoja, contrariamente a lo observado para Brachiaria.
Al considerar el comportamiento de la relación entre los asimilados enviados a la hoja respecto a los enviados al tallo, en función del genotipo, la época y la dosis de cal aplicada, se encontró que la mayor relación hoja/tallo para el pasto Brachiaria se dio a los 60 y 90 DDE cuando se aplicaron 9 ton de CaCO3/ha con 4.02 y 3.74 respectivamente. Por su parte, en esta misma época en pasto estrella esta relación pareció mas estable en función del encalamiento; las mayores relaciones hoja/tallo fueron de 1.20 (3 ton de CaCO3/ha) y 1.29 (sin aplicación de cal) a los 60 y 90 dds respectivamente.
DISCUSIÓN
Características químicas del suelo. A los 90 DDE, el suelo mostró una reacción muy ácida con una tendencia a decrecer con las dosis de cal, comparada con la del suelo sin encalar (4.67), tal como se mostró en la tabla 1. Lo anterior indica que los hidróxidos provenientes de la calcita neutralizaron gran cantidad de iones de hidrógeno y de aluminio presentes en la solución del suelo, lo cual produjo un aumento significativo del pH, resultado que coincide con estudios en donde se encontró que el pH aumenta significativamente como respuesta al encalado (14). Aunque la materia orgánica es muy alta, no presentó cambios considerables con respecto a los valores iniciales. Por su parte, el azufre disminuyó con las dosis de cal, lo cual indica que los iones de calcio reaccionaron con los iones sulfato, produciendo la insolubilización de éste elemento.
El fósforo, presentó valores medios con una tendencia a disminuir con las dosis de cal aplicada, lo que demuestra que no se presentó solubilización de este elemento; esto difiere con resultados de otros autores, quienes evaluando la frecuencia de encalado en suelos ácidos de los Llanos Orientales, encontraron que el pH y el contenido de calcio aumentaban con el encalado, existiendo una tendencia a aumentar la disponibilidad del fósforo conforme se incrementaba el calcio disponible (15). Los resultados observados en el presente estudio, probablemente pueden deberse a que el aumento de la actividad iónica del calcio en la solución, ocasionó la unión y precipitación de este elemento con el fósforo.
El calcio, como se esperaba, aumentó con las aplicaciones de cal, lo que demuestra que la calcita aplicada se disolvió eficientemente produciendo iones calcio y bicarbonato; esto concuerda con un estudio realizado en esta área, en donde se encontró que el encalado aumenta los contenidos de calcio en el suelo (16). El magnesio y potasio presentaron contenidos altos con valores menores al inicio, lo cual sugiere que no se presentó un desplazamiento de estos elementos de la fase soluble y que el sodio no se concentró de manera excesiva. La acidez intercambiable disminuyó con las aplicaciones de cal, indicando que se neutralizó gran cantidad de aluminio intercambiable, disminuyendo así los porcentajes de saturación del aluminio. Estos resultados concuerdan con estudios en los que se indica que el encalamiento presenta efectos positivos al corregir la acidez en el suelo y reducir la toxicidad por aluminio (17).
A los 150 DDE, sigue siendo consistente la reacción del suelo al aumentar con las dosis de cal, lo que demuestra que la cal neutralizó significativamente la acidez del suelo, coincidiendo con trabajos que encontraron que el pH del suelo se incrementó en forma lineal y proporcional con las dosis de cal aplicada (14). La materia orgánica presentó valores muy altos, similares a la época anterior, sugiriendo que la neutralización de la acidez del suelo, permitió aumentar la población de microorganismos, con el consecuente efecto sobre la mineralización de este; el azufre con su tendencia a disminuir con las dosis de cal agrícola aplicada, sugiere que ésta última inmovilizó parte del elemento, formando sulfato de calcio (yeso) y agua. El comportamiento del fósforo continuó con la misma tendencia de los 90 días después del encalado, confirmando que el calcio lo precipitó.
Los altos contenidos de calcio en este período pueden ser explicados por la alta solubilidad de la cal aplicada, suministrando iones calcio al suelo, lo que coincide con reportes que afirman que el efecto directo de los materiales encalantes tales como el carbonato de calcio, consiste en el aumento del calcio en el perfil, en la reducción del aluminio intercambiable y en la consecuente inducción de un aumento del pH del suelo (18). Por su parte, los valores altos de magnesio y potasio indican que se presentó solubilización y desplazamiento de estos elementos de la fase intercambiable, coincidiendo con trabajos que indican que el encalado disminuye el porcentaje de saturación del aluminio, aumentando la saturación de las bases (19). Los niveles de sodio en este período, pueden considerarse de medios a excesivos, con tendencia a aumentar con las aplicaciones de cal, lo que sugiere que podrían presentarse problemas de formación de sales fitotóxicas. La acidez intercambiable disminuyó, producto de la neutralización del aluminio intercambiable por acción de la cal.
Análisis fisiológico de los pastos
Peso fresco. Los resultados observados por el efecto de la época y del encalamiento sobre le peso fresco de los pastos, sugieren que este encalamiento, además de neutralizar el efecto negativo de algunos elementos como el aluminio en este caso. Esto permitió un mayor desarrollo de raíces, lo que conllevaría a una mayor área de exploración de volumen de suelo con la consecuente eficiencia en la captación de agua y nutrientes, situación que expresa mejor B. decumbens. Esto coincidiría con los resultados de un trabajo en donde se concluye que la adaptación y persistencia de Brachiaria decumbens es atribuida a la habilidad de este genotipo para asimilar nutrientes importantes como el N, P y Ca, en un ambiente de bajo pH y alta acidez intercambiable, así como de buena disponibilidad de agua en el suelo (12). Asímismo, en estudios realizados en un ultisol (suelo ácido), se encontró un aumento importante en el peso seco de Brachiaria decumbens como respuesta a los tratamientos con encalado (20).
Esta respuesta puede favorecerse además por la disminución de las concentraciones de aluminio, cuyo exceso lleva a modificaciones en las hojas, reducción en la apertura estomática, disminución de la actividad fotosintética e inhibición del crecimiento de las raíces. La reducción en el crecimiento de raíces se traduciría en una sistemática deficiencia en la asimilación de calcio y fósforo, trayendo como consecuencia una reducción significativa de la biomasa de la planta.
Al considerar la integración de las respuestas de cada genotipo en función de la época de corte y la dosis de cal aplicada (Figura 3), las ventajas comparativas del pasto Brachiaria sobre el pasto estrella, pueden estar explicadas por los mayores contenidos de humedad del primero de estos genotipos, como consecuencia de un mejor estado hídrico y uso eficiente del agua, durante los primeros meses del cultivo, cuando el agua no fue un factor limitante. Resultados similares se reportaron en un trabajo de nutrición en Brachiaria en los Llanos Orientales de Colombia, donde se encontró que esta gramínea pasó de producir 0,75 ton/ha de masa seca en la estación seca a 1,55 en la de lluvias (21). A esto podría sumarse también el efecto de la cal que, al neutralizar los elementos acidificantes presentes en este tipo de suelos, permitió un mejor desarrollo radicular, haciendo más eficiente la captación de agua y nutrientes como se discutió anteriormente. Otros trabajos de evaluación fisiológica también han demostrado el mejor comportamiento de Brachiaria respecto a estrella en aspectos como producción de forraje y contenido de proteína bruta (22).
Peso Seco. Los mayores valores observados para el pasto Brachiaria en condiciones de alta disponibilidad de agua (mayor precipitación), confirman la mayor eficiencia fisiológica de este material bajo condiciones adecuadas de humedad, lo que explicaría las diferencias significativas a favor de Brachiaria a los 120 y 150 DDE. El factor humedad es considerado como el que más afecta el crecimiento de las plantas, principalmente gramíneas, debido a que, además de intervenir en el crecimiento celular por las presiones de turgencia en los meristemos, juega un papel muy importante en la solubilidad y disponibilidad de nutrientes para pastos y forrajes, tales como el nitrógeno, fósforo y potasio que controlan la promoción y desarrollo de nuevos brotes y aumentan el numero de hojas por planta y con ello el área foliar y la producción de biomasa.
Algunos autores analizando diferentes especies de gramíneas en varias fechas de corte observaron una mayor producción de biomasa durante las épocas de mayor precipitación (23). Esto coincide con lo encontrado en las dos especies de pastos evaluadas en el presente trabajo, las cuales mostraron un comportamiento decreciente en su peso seco en la medida que la disponibilidad de agua fue menor, independientemente del nivel de encalamiento, sugiriendo que las precipitaciones contribuyeron de manera positiva en la producción de biomasa. Esta situación también es comparable a los resultados obtenidos en otros trabajos en donde encontraron que el rendimiento en materia seca de Brachiaria brizantha disminuía a medida que se incrementaba el numero de cortes, motivado por el comportamiento irregular de las precipitaciones (24). El agua tiene efectos directos e indirectos sobre las actividades metabólicas, ya que es una de las materias primas necesarias para la producción de carbohidratos, aminoácidos y otros. Por otra parte, es importante resaltar que el peso seco indica la eficiencia en la producción de asimilados que cada material realizó durante cada período de crecimiento, ya que, aunque la disponibilidad de agua fue el factor determinante en la expresión del crecimiento y acumulación de biomasa, la medición del peso seco cuantifica esta eficiencia al ser eliminada el agua de sus tejidos.
Es importante resaltar que Brachiaria decumbens se caracterizó por presentar los mayores pesos secos durante todo el experimento, lo cual puede deberse a sus características genéticas, ya que es un material adaptado a condiciones de alta acidez intercambiable y ácidos orgánicos que se generan con la oxidación de la pirita. Esta apreciación estaría acorde con investigadores (9, 10), quienes exponen que a diferencia de otros pastos, B. decumbens tolera elevados contenidos de aluminio ya que puede acumular altas cantidades de este en las raíces. Además puede sintetizar eficientemente ácidos orgánicos que juegan un papel importante en la detoxificación del aluminio tanto a nivel interno como externo (9,10). Asímismo, un trabajo desarrollado por Bernal (13) confirmó que esta especie se adapta bien a diversas condiciones agroecológicas como altitud (desde el nivel del mar hasta los 2.200 msnm), es tolerante a la sequía, quemas, alta precipitación, pastoreo intensivo, pisoteo, suelos ácidos y pobres en nutrimentos y alta saturación de aluminio.
Al parecer, la resistencia genética de pasto estrella estaría mas limitada por los mecanismos fisiológicos que especies de esta familia utilizan al compararlos con los del pasto brachiaria. En este sentido, se conoce que callos celulares de genotipos de Cynodon sp, sometidos a concentraciones diferenciales de aluminio, utiliza diversos mecanismos rizosféricos externos para contrarrestar la toxicidad, como el aumento del pH extracelular y la captura de iones Al3+ en la pared (25). Sin embargo, no ha sido evidente la producción de ácidos orgánicos como el ácido cítrico, como mecanismo de resistencia (25); otros autores indican que estos mecanismos aun no se conocen bien (26).
Relación hoja/tallo. La relación hoja/tallo indica la eficiencia del genotipo en el envío de asimilados a la lámina foliar respecto a la formación de tejidos de sostén (tallo), probablemente como una respuesta a condiciones favorables, lo que le permitiría a la planta aumentar las tasas de fotosíntesis. En el presente trabajo, esta condición se expresó de manera evidente en B. decumbens. Es probable que las condiciones genéticas del pasto estrella condicionen el balance de la relación hoja/tallo, manteniéndola relativamente estable a través de variaciones ambientales, tales como la disponibilidad de agua en el suelo. Los valores de la relación hoja/tallo encontrados en la investigación para el pasto Brachiaria, fueron mayores a los encontrados por Vallejo et al (27), quienes reportaron valores de 1.3 ± 0,3. Por otra parte, en trabajos realizados en Brachiaria mutica se encontró una baja relación hoja/tallo con valores que estaban por el orden de 0,666 (28). En estudios sobre el comportamiento de cuatro especies de gramíneas en el Medio Sinú colombiano, se encontró una relación hoja/tallo para B. decumbens de 1,12 (29). Lo anterior confirmaría la plasticidad de esta relación, en respuesta a la variabilidad de las condiciones ambientales, tal como se mencionó anteriormente.
El comportamiento decreciente de la relación hoja/tallo del pasto Brachiaria a partir de lo 90 DDE, probablemente se deba a la capacidad de las plantas de producir nuevos rebrotes, superando la producción de hojas, además de la formación de tallos gruesos y resistentes que se incrementan con el tiempo, como consecuencia lógica de la senectud. Estos resultados estarían de acuerdo con lo reportado por Alvarado et al (30) donde se evaluó el comportamiento de B. decumbens bajo diferentes dosis de nitrógeno, encontrando relaciones hoja/tallo que decrecieron con el tiempo desde 1.32 hasta 0.63
En conclusión, la aplicación de dosis crecientes de cal a suelos sulfatados ácidos incrementó los valores de pH, así como la producción de biomasa en la pastura independientemente de la especie forrajera.
REFERENCIAS
1. Golez N. Formation of acid sulfate soil and its implications for brackishwater ponds. Aquac Eng 1995; 14: 297-316. [ Links ]
2. Jusop S, Muhrizal S. Pyrite in acid sulfate soils: transformation and inhibition of its oxidation by application of natural materials. 2007. [acceso 5 de febrero de 2007]. URL disponible en: http://www.ldd.go.th/Wcss2002/papers/0097.pdf. [ Links ]
3. Cook F, Hicks W, Gardner E, Carlin G, Froggatt D. Export of acidity in drainage water from acid sulfate soils. Mar Pollut Bull 2000; 41: 319-26. [ Links ]
4. Nowak J, Friend A. Aluminum fractions in root tips of slash pine and loblolly pine families differing in Al resistance. Victoria, Canada. Tree Physiol 2005; 25: 245-250. [ Links ]
5. Kochian L. Cellular mechanisms of aluminum toxicity and resistence in plant. Annu rev plant physiol plant mol biol 1995; 46: 237-260. [ Links ]
6. Ward N, Sullivan L, Bush R. Sulfide oxidation and acidification of acid sulfate soil materials treated with CaCO3 and seawater-neutralised refinery residue. Australian journal of soil research 2002; 40: 1057-1067. [ Links ]
7. Tang D, Garvin L, Kochian M, Sorrells E, Carver B. Physiological Genetics of Aluminum Tolerance in the Wheat Cultivar Atlas 66. Crop Sci 2002; 42: 1541-1546. [ Links ]
8. Watanabe T, Osaki M. Mechanisms of adaptation to high aluminum condition in native plant species growing in acid soils: a review. Comm Soil Sci Plant Anal 2002; 33: 7-8. [ Links ]
9. Taylor G, McDonald-Stephens J, Hunter D, Bertsch P, Elmore D, Rengel Z. et al. Direct measurement of aluminum uptake and distribution in single cells of Chara corallina. Plant Physiol 2000; 123: 987-996. [ Links ]
10. Feng J. Physiological Mechanisms of Al Resistance in Higher Plants. Soil Sci Plant Nutr 2005; 51(5): 609-612. [ Links ]
11. Marschner H. Mineral nutrition of higher plants. London: Academic Press; 2002. [ Links ]
12. Rao I, Kerridge P, Macedo M. Nutritional requirements of Brachiaria and adaptation to acid soils. In: Biology, Agronomy and Improvement: Cali, Colombia: CIAT and EMBRAPA-CNPGC; 1996. [ Links ]
13. Bernal J. Pastos y forrajes tropicales, producción y manejo. 3a ed. Santa fé de Bogotá D.C. Colombia: Banco Ganadero. 1994. [ Links ]
14. Barroti G, Vahas E. El fósforo y el encalado sobre las fosfatasas y la producción de Braquiaria ruziziensis y Cajanus cajan. Agronomía Trop 2003; 53(2): 209-225. [ Links ]
15. Rodríguez T, Tenias J. Frecuencia del encalado y su efecto sobre el rendimiento del maní en un suelo ultisol de los Llanos Orientales. Agronomía Trop 1983; 33: 243-272. [ Links ]
16. Tenias J. Efecto del encalado en la producción de cuatro variedades de caña de azúcar sembradas en un ultisol del estado Monagas. Caña de azúcar 1989; 7(1): 5-16. [ Links ]
17. Guimaraes E, Sanz I, Rao I, Amezquita M, Amezquita E. Sistemas agropastoriles en sabanas tropicales de América Latina. Cali: Publicación CIAT; 2000. [ Links ]
18. Mengel K, Kirkby E. Principios de nutrición vegetal 4a ed. Suiza: Basel; 2000. 474-475. [ Links ]
19. Pérez R. Efectos del encalado en la neutralización del aluminio intercambiable y sobre el crecimiento del tomate (Lycopersicon esculentum). Agronomía Trop 1986; 36(1-3): 89-110. [ Links ]
20. Pérez M, Smyth T. Comparación del efecto de dos especies forrajeras sobre el ph de la rizósfera y la disolución de rocas fosfóricas de diferente reactividad. Revista de la Facultad de Agronomía LUZ 2005; 22: 142-154. [ Links ]
21. Rincon A. Rehabilitación de pasturas y producción animal en Brachiaria decumbens en la Altillanura plana de los Llanos Orientales de Colombia. Pasturas Tropicales 2004; 26(3): 2-12. [ Links ]
22. Botrel M, Alvim J, Ferreira D. Avaliação de gramíneas forrageiras na região sul de minas gerais. Pesqui Agropecu Bras 1999; 34 (4): 683-689. [ Links ]
23. Romero S, Alfonso R, Medina R, Flores R. Evaluación inicial de la fertilización con roca fosfórica en tres especies del género Brachiaria. Zootecnia Trop 2003; 21(2): 183-196. [ Links ]
24. Pietrosemoli S, Faria L, Villalobos N. Respuesta del pasto B. brizantha a la fertilización nitrogenada. Revista de la Facultad de Agronomía-LUZ 1996; 13(5): 551-560. [ Links ]
25. Ramgareeb S, Cooke J, Watt M. Responses of meristematic callus cells of two Cynodon dactylon genotypes to aluminium. J Plant Physiol 2004; 161: 1245-1258. [ Links ]
26. Hinsinger P, Plassard C, Tang C, Jaillard B. Origins of root mediated pH changes in the rhizosphere and their responses to environmental constraints: a review. Plant Soil 2003; 248: 43-59. [ Links ]
27. Vallejo A, Pizarro E, Chaves C, Pezo D, Ferreira P. Evaluación agronómica de gramíneas en Guapiles, Costa Rica. 2 Ecotipos de Panicum maximum. Pasturas Tropicales 1989; 11(2): 10-15. [ Links ]
28. Entrena I, Chacón E, González V. Influencia de la carga animal y la fertilización con azufre sobre las tasas de crecimiento, biomasa y producción aérea neta de una asociación de Brachiaria mutica y Teramnus uncinatus. Zootecnia Trop 1998; 16(2): 183-206. [ Links ]
29. Cuadrado H, Torregroza L, Jiménez N. Comparación bajo pastoreo con bovinos machos de ceba de cuatro especies de gramíneas del género brachiaria. Rev MVZ Córdoba 2004; 9(2): 438-443. [ Links ]
30. Alvarado A, Arriojas L, Chacón E, Rodríguez S, Chacin F. Estudios sobre henificación del pasto barrera (Brachiaria decumbens), en condiciones de sabanas del Piedemonte Barinés. Zootecnia Trop 1990; 8(1 y 2): 17-36. [ Links ]

