










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista MVZ Córdoba
versão impressa ISSN 0122-0268versão On-line ISSN 1909-0544
Rev.MVZ Cordoba v.13 n.2 Córdoba maio/agos. 2008
ZOOPLANCTON EN LA LARVICULTURA DE PECES NEOTROPICALES
ZOOPLANKTON IN LARVICULTURE OF NEOTROPICAL FISHES
Martha Prieto G1*, M.Sc, Victor Atencio G1, M.Sc.
Universidad de Córdoba, Facultad de Medicina Veterinaria y Zootecnia, Departamento de Ciencias Acuícolas, CINPIC. Montería, Colombia. *Correspondencia: mjprieto@sinu.unicordoba.edu.co
Recibido: Febrero 1 de 2008; Aceptado: Julio 30 de 2008.
RESUMEN
La larvicultura de peces neotropicales altriciales presenta limitaciones para el manejo de la primera alimentación dado el pobre desarrollo del tracto digestivo y capacidad natatoria. La alimentación de la larva debe considerar el tamaño, densidad y calidad de la presa ofrecida. Este documento revisa a la luz de recientes trabajos la incidencia del zooplancton en la larvicultura de peces, aborda la importancia de la larvicultura en el proceso piscícola, la importancia del zooplancton como alimento y las alternativas en el manejo de cladóceros, copépodos y rotíferos para la alimentación de larvas de especies neotropicales.
Palabras clave: Zooplancton, larva, peces, acuicultura, alimentación, tropico.
ABSTRACT
Because of the poor development of digestive system and swimming capability of young fish, larviculture of altricial neotropical fish presents for the management of the initial feeding. The feeding of the larvae must consider the size, density and quality of the prey offered. This document reviews the incidence of zooplankton in fish larvicultura, approaches the importance of larviculture in fish rearing, the importance of zooplankton as prey, and alternatives in the handling of cladocerans, copepods and rotifers for the feeding of larvae of neotropical fishes.
Key words: Zooplankton, larvae, fishes, aquaculture, feeding, tropic.
INTRODUCCIÓN
Uno de los puntos críticos en el ciclo de producción de peces, es sin duda, la fase de larvicultura, la cual requiere de alimentos externos apropiados tanto cuantitativa como cualitativamente. La disponibilidad de alevinos en cantidades y de buena calidad, se considerada un factor crítico para el éxito de la producción intensiva, en la cual la alimentación y la nutrición han sido señaladas como los principales factores responsables de los frecuentes desaciertos en la larvicultura, constituyendose en el cuello de botella que impide la expansión de la actividad.
Las postlarvas de la mayoría de especies de peces son planctófagas, principalmente zooplanctófagas, aun cuando sus adultos sean herbívoros, omnívoros o carnívoros (1, 2). Las larvas de la mayoría de especies comerciales tropicales poseen reservas vitelinas escasas, siendo denominadas como altriciales; las cuales, cuando inician la alimentación exógena presentan el tracto digestivo aún no completamente formado, el intestino anterior todavía esta indiferenciado y sin glándulas gástricas (3, 4).
La larva debe aprender a capturar, engullir y asimilar el alimento; por esto requiere en esta etapa de desarrollo una dieta especial de partículas pequeñas, de textura suave, fácilmente digerible, en forma constante, abundante y con alto valor nutritivo. El incompleto desarrollo de su tracto digestivo limita su capacidad de aprovechar satisfactoriamente los nutrientes de la dieta (1, 2, 5, 6), utilizando las enzimas de las presas, constituidas principalmente por zooplancton, para facilitar el proceso de digestión y estimular la producción de las enzimas endógenas. Así, este periodo es una fase crítica para su sobrevivencia, al contrario de las larvas precociales (con abundante vitelo); las cuales asimilan eficientemente el alimento artificial desde el inicio de la alimentación exógena, gracias a que presentan el tracto digestivo diferenciado y, por lo tanto, son menos dependientes del zooplancton para el proceso de digestión. Este trabajo revisa a la luz de recientes trabajos la incidencia del zooplancton en la larvicultura de peces.
Importancia de la larvicultura en el proceso piscícola. El manejo inadecuado de la primera alimentación es una de las barreras para el éxito de la larvicultura de peces (4, 6 - 8, 9). En la larvicultura de los peces las mayores limitaciones están dadas por el tamaño de la boca, pobre capacidad natatoria, densidades inadecuadas de presas, composición bioquímica del alimento y el precario estado de desarrollo del aparato digestivo con la consecuente ausencia de enzimas digestivas al inicio de la alimentación exógena (4). En las especies neotropicales altriciales, antes que se agote el vitelo y la boca esté bien desarrollada, se debe suministrar el primer alimento, principalmente zooplancton seleccionado de acuerdo con el tamaño de la boca de la larva.
Hoy existen tres procedimientos principales para la alimentación inicial de las larvas. El primero es el uso de zooplancton proveniente de colectas en ambiente natural o la concentración de las postlarvas en estanques en tierra fertilizados, luego de la abertura de la boca (6). El segundo es la larvicultura intensiva con el uso de organismos zooplanctónicos (rotíferos, cladóceros, copépodos y artemia) cultivados en laboratorio (6,8-10). El tercer procedimiento es la introducción precoz de alimento inerte, principalmente raciones microencapsuladas ( 2, 4, 7).
La técnica de larvicultura adoptada por la mayoría de los piscicultores neotropicales consiste en sembrar directamente las postlarvas en estanques fertilizados inmediatamente después del inicio de la alimentación exógena (9). Esa técnica generalmente resulta en bajas tasas de sobrevivencia dificultando la producción de alevinos a gran escala, la producción se torna muy variable, altamente dependiente de las condiciones ambientales, tales como temperatura, abundancia de alimento apropiado, presencia de predadores, enfermedades, entre otros, lo que no permite la proyección de la producción en una etapa posterior y se podría considerar como una larvicultura semi-intensiva.
Alternativamente existe la posibilidad de mantener las larvas en laboratorio donde permanecen protegidas de predadores y reciben alimentos de calidad y en cantidad adecuada a su desarrollo inicial; lo cual se podría considerar como una larvicultura intensiva. Posteriormente cuando están más crecidas, son transferidas a los estanques externos. Sin embargo es una técnica que eleva los costos de producción, siendo utilizada, apenas por algunos productores (2,10). El cultivo de larvas en laboratorio permite investigaciones mas detalladas sobre los hábitos y preferencias alimentares y sobre el comportamiento de las larvas, información imprescindible para el desarrollo de la piscicultura neotropical.
Se ha observado que las tasas de sobrevivencia en el alevinaje se incrementan cuando se realiza el manejo del inicio de la alimentación exógena en condiciones controladas, utilizando zooplancton, principalmente nauplios de Artemia, zooplancton silvestre seleccionado por tallas y libre de predadores y larvas forrajeras (8, 9, 11). Uno de los inconvenientes en el uso de zooplancton silvestre es la posibilidad de introducir patógenos y predadores al sistema de cultivo, tales como copépodos ciclopoides (8, 11).
Los efectos del manejo de la primera alimentación son fundamentales para garantizar un buen crecimiento y sobrevivencia de los alevinos. Las ventajas de la larvicultura intensiva se basan en evitar las influencias ambientales desfavorables, generar condiciones ambientales óptimas, disminuir el factor de conversión alimentaría, aumentar la tasa de sobrevivencia, mantener la producción de peces independiente de factores estacionales, mejorar el periodo de producción y producir peces de manera más continua para el mercado. En el mismo sentido y tendiendo en cuenta los costos de producción; con base en los resultados obtenidos por diferentes autores; se recomienda el mantenimiento de las larvas altriciales de peces tropicales, en sistemas de lavicultura intensiva, por no más de seis días (2, 8,10).
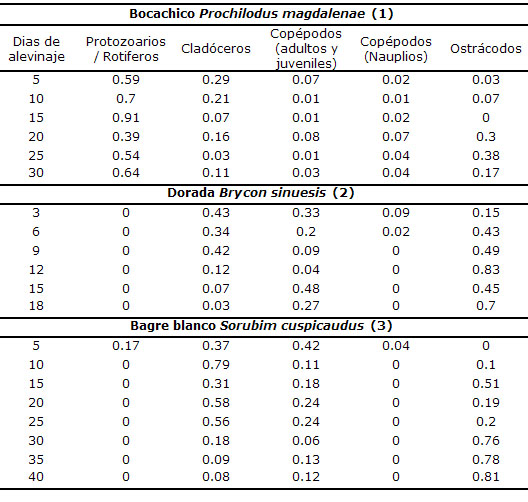
Entre las especies altriciales neotropicales de interés acuícola se incluyen el Bagre blanco (Sorubim cuspicaudus), Bagre rayado, (Pseudoplatystoma magdaleneatum), Cachama negra (Colossoma macropomum), Cachama blanca (Piaractus brachypomus), Bocachico (Prochilodus magdalenae), Dorada (Brycon sinuensis), yamú (Brycon amazonicus) Pacu (Piaractus mesopotamicus), Surubin (Pseudoplatystoma corruscans), Dourada (Salminus brasiliensis), Piracanjuba (Brycon orbignyanus) y Curimbatá (Prochilodus scrofa), entre otras. El primer alimento externo de estas especies es el zooplancton, compuesto principalmente por: protozoarios, rotíferos, cladóceros, copépodos, entre otros organismos. Durante su transformación de larva a alevino el bocachico muestra preferencia por grupos del zooplancton como protozoarios/rotíferos (12); mientras que dorada y bagre blanco por cladóceros, juveniles y adultos de copépodos y ostrácodos (13, 14) (Tabla 1). Las enzimas proteolíticas del propio zooplancton son liberadas por acción física de las larvas durante la captura e ingestión (15); estas enzimas exógenas desencadenan la hidrólisis de las proteínas del propio zooplancton ingerido, estimulando la secreción de enzimas endógenas por el tracto digestivo de las postlarvas.
Tabla 1. Grupos del zooplancton preferidos por tres especies de peces neotropicales durante su transformación de larva a alevino en estanques en tierra (alevinaje). El número corresponde al valor promedio del índice de Chesson, el cual indica preferencia cuando es mayor de 0.2.
Larvas de peces siempre son alimentadas con altas densidades de presas durante la larvicultura. Densidades de presas elevadas proporcionan mayor tasa de encuentro entre predador y presa y, consecuentemente, mayor consumo del alimento; una mayor alimentación generalmente resulta en más rápido crecimiento y desarrollo, mejores condiciones generales de las larvas y altas tasas de sobrevivencia (16). Igualmente importante es el tamaño de la presa, estudios que comparan la disponibilidad de presas en un ambiente con las presas ingeridas por las larvas de peces confirman que la característica del tamaño de la presa afecta fuertemente los patrones de selectividad por el alimento (8, 11). Otro aspecto relevante es la frecuencia de alimentación. Normalmente, el número de veces que las larvas deben ser alimentadas es mayor en las primeras fases de vida. Durante la larvicultura, es común ofrecer zooplancton varias veces al día. La frecuencia de dos a cuatro dosis de alimento en el día es más que adecuada para incrementar significativamente la cantidad de alimento ingerido por las larvas, maximizar la tasa de crecimiento, homogenizar el tamaño y controlar el canibalismo (6, 7, 16).
Importancia del zooplancton como alimento. Varios experimentos con larvas de especies neotropicales de agua dulce, destacándose pacú, Cachama negra, Piracanjuba Brycon hilarii, Curimbatá, Matrinxã Brycon cephalus, Bagre blanco, Bocachico, Yamú han demostrado la necesidad del uso de alimentos vivos, existiendo preferencia por pequeños cladóceros, nauplios y copepoditos de copépodos y rotíferos (7, 8,11, 17, 18). Ya en especies tropicales que presentan conducta caníbal al inicio de la alimentación exógena, es viable y mas recomendable el uso de larvas forrajeras, como en el caso de yamú, dorada Brycon sinuensis y otras especies del género Brycon (11) que presentan mejor desempeño en el alevinaje al usar larvas de otras especies de peces como alimento vivo en la primera alimentación (Tabla 2).
Tabla 2. Sobrevivencias obtenidas por varios autores durante el alevinaje de especies del genero Brycon cuando manejaron la primera alimentación con larvas forrajeras (HPE, horas post-eclosión; L/m2 Larva por metro cuadrado).
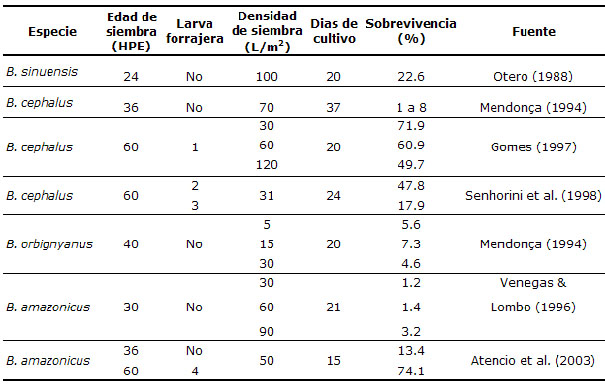
1. Con larvas de Curimbatá (Prochilodus scrofa), 2. Con larvas de Pacú (Piaractus mesopotamicus) 3.Con zooplancton; 4. Con larva de Cachama blanca (Piaractus brachypomus).
La composición bioquímica del zooplancton para los peces es importante, siendo considerado el alimento que contiene la mayoría de las sustancias nutritivas y que sirve como base para las dietas experimentales. Principalmente, el valor nutritivo se basa en el contenido de aminoácidos y ácidos grasos esenciales, entre otros elementos que favorecen el crecimiento y la sobrevivencia de las postlarvas (5,19). El zooplancton debido a su contenido de ácidos grasos esenciales, es una buena opción para la nutrición de las larvas, en general los alimentos naturales presentan altos niveles de proteína de excelente calidad (1, 2, 5, 20, 21), siendo fuente importante de vitaminas y minerales (15, 20). El plancton posee enzimas necesarias para el crecimiento y sobrevivencia de las larvas (1, 3, 5,19). Además, el movimiento natural de esos organismos zooplanctónicos estímula el comportamiento predador de las larvas (19; 7) y en cantidad adecuada no compromete la calidad del agua (5,19, 20).
A pesar de los esfuerzos para sustituir totalmente el alimento vivo por dietas artificiales, continúa la dependencia de la producción y empleo de zooplancton para la larvicultura de especies neotropicales. En general, el alimento artificial no suple las necesidades nutricionales o no presenta las características adecuadas para las larvas, constituyendo el zooplancton la mejor opción en la larvicultura (2, 22). Su uso en la larvicultura presenta como principales ventajas: menor grado de polución, mejor distribución, mantienen sus características por muchas horas (1,19), lo que no ocurre con alimentos artificiales. Además, el zooplancton presenta corto ciclo de vida, alta tasa de fertilidad y capacidad de vivir en altas densidades, características que facilitan su cultivo; su lento movimiento y coloración facilitan la captura por parte de las postlarvas, asi mismo, presentan la posibilidad de ser biocápsulas al ser enriquecidos.
Entre los grupos de zooplancton más utilizados están Artemia, rotíferos, cladóceros y copépodos. El valor nutricional de los rotíferos esta sujeto al alimento ofrecido; son considerados excelente alimento para larvas de peces marinos y algunos de agua dulce, gracias a su pequeño tamaño, constante movimiento en el agua, corto ciclo de vida para su cultivo (21). Son considerados de alto valor nutritivo por su digestibilidad y capacidad de transferencia de nutrientes cuando son enriquecidos. Para las especies de peces tropicales de agua dulce, en la década de los 80 consideraban que en compañía de algunos protozoarios de gran tamaño, los rotíferos eran la mejor opción en la primera alimentación, gracias a su reducido tamaño, lenta natación que permite fácil captura. Esta concepción cambió a inicios de los años 90 cuando se realizaron estudios para determinar las preferencias alimentares de las larvas de especies con importancia piscícola determinando que la selección y el consumo están orientados hacia los cladóceros y los copépodos.
La abertura de la boca al inicio de la alimentación exógena es determinante para la selección que hace el individuo del tamaño del alimento (23, 24). Las larvas seleccionan el alimento dependiendo de la abertura bucal para lograr una alimentación más eficiente con relación al tamaño de la presa, para especies neotropicales con restringido tamaño de boca al inicio de la alimentación exógena como liseta (Leporinus muyscorum, 350 mm), puede ser viable el uso de los rotíferos. Según Werner & Hall (25), la selección del tamaño es un mecanismo que el predador utiliza para optimizar la energía invertida en la captura de la presa.
La calidad nutricional de los copépodos se caracteriza por altos niveles de proteína (44-52%) y buen perfil de aminoácidos, la composición de ácidos grasos varia considerablemente acorde al alimento usado en su cultivo (19, 26, 27). A pesar de presentar movimientos rápidos, por saltos y consecuentemente buen escape del predador, su nauplio es considerado excelente alimento para larvas de peces gracias sus movimientos más lentos siendo fácilmente capturados por las postlarvas de peces marinos y de agua dulce (5, 27). Los copépodos son versátiles para la alimentación gracias a que presentan en su desarrollo diferentes tamaños que permiten su selección acorde a las necesidades de las larvas.
En los cladóceros, la fuente de alimentación determina su calidad nutricional. Además de poder elevar su contenido de ácidos grasos con una adecuada dieta (28), presentan un espectro de enzimas importantes (proteinasas, peptidasas, amilasas, lipasas y celulasas) que sirven como exoenzimas en el intestino de las larvas (1, 5). Se tiene claramente establecida la selectividad por cladóceros y copépodos en las especies neotropicales de agua dulce; tal es el caso de Dorada, Yamú, Bagre blanco, Bocachico, Pacu, Tambaqui, Curimbatá (8, 11, 29, 30). En general ha sido reportada la preferencia por el consumo de zooplancton de mayor tamaño (cladóceros y copépodos) y consumo insignificante de rotíferos e protozoarios en las mayoría de las especies neotropicales donde fue evaluado el régimen alimentar en la fase de alevinaje.
La preferencia por zooplancton mayor, puede ser explicada por las ventajas de la mayor eficiencia en el balance energético de las presas de mayor tamaño. En especies como el bagre blanco la abertura bucal máxima al inicio de la alimentación exógena es de 603.3±32.3 µm (17), similar a la reportada para el bocachico (642-671 µm) (8), pero mucho más pequeña que la reportada para el yamú (1470 µm) (11). Según Alcala & Ortega (17), el bagre blanco cuando comienza a comer, su abertura bucal le permite la captura de zooplancton como cladóceros y copépodos, sugiriendo que el tamaño apropiado para el manejo de primera alimentación de esta especie debe oscilar entre 250 y 450 µm. Los resultados de diversos estudios han mostrado que el consumo de zooplancton de mayor tamaño puede dar mejores tasas de crecimiento y sobrevivencia (11, 31). Otras características de las presas que pueden influir en la selección son: morfología, palatabilidad, facilidad de manejo, digestión y abundancia, así como el futuro riesgo de predación (32).
Artemia es el zooplancton más usado en la acuicultura. En los últimos años ha tenido papel central en el desarrollo de la piscicultura debido a su contenido de ácidos grasos (n-3 y n-6), que permite su suministro a especies de agua dulce y marina, y más de 47% en contenido de proteínas (33). Además su importancia se basa en la practicidad del almacemaniento y el manejo de sus cistos. Debido a su tamaño en el estadio de nauplio es un alimento práctico para muchas larvas de peces y crustáceos en la larvicultura comercial (33, 34, 35). Presenta varias enzimas proteolíticas con importante función en el tracto digestivo de las larvas (19). Sin embargo, la disponibilidad de quistes no es predecible, porque se cosechan de grandes ecosistemas naturales y los factores que interfieren en su producción están fuera del control humano; por lo que son frecuente crisis de producción de Artemia que ocasiona fluctuaciones marcadas de los precios (36).
Aquí la problemática actual de la Artemia genera la necesidad de contar con alternativas de producción masiva de otros organismos zooplanctónicos que sean aptos para la alimentación de las larvas, que presenten además adecuado valor nutritivo, facilidad de manejo y características que hagan eficiente su cultivo. En los diferentes ecosistemas acuáticos tropicales existe una gran variedad de organismos zooplanctónicos con potencial acuícola, por esto deben intensificarse los estudios sobre las especies que puedan ser fuente de alimento para larvas de peces.
Manejo de zooplancton para la alimentación de larvas. La producción de zooplancton es una práctica restringida a pocos organismos. Su cultivo se basa en la alimentación con diferentes especies de microalgas y levadura. Entre los rotíferos, el género más cultivado Brachionus; destacándose Brachionus plicatilis como la especie más cultivada en el mundo, seguida por B. callyciflorus, B. rubens, B. urceolaris y B. falcatus (21). Entre los cladóceros, principalmente los géneros Daphnia y Moina, son de gran importancia en la piscicultura. La mayoría de las especies de copépodos producidas y usadas como presa viva, son calanoides y harpacticoides, los copépodos planctónicos comúnmente encontrados en este sistema incluyen géneros como Acartia, Centropages y Temora, los copépodos harpacticoides también proliferan y son frecuentemente encontrados en los estómagos de las postlarvas siendo desarrolladas técnicas de cultivo para el género Tisbe (37).
En la última década, para atender los requerimientos nutricionales de postlarvas altriciales, ha sido el enriquecimiento un importante método para transferir toda clase de elementos esenciales a través de los organismos zooplactónicos (20, 38). Entre los diferentes enriquecimientos realizados se registran con vitamina C (39, 40), vitamina E (40), probióticos (41), antibióticos, fosfolipídios (42) y ácidos grasos (20, 38, 40, 43, 44).
El enriquecimiento en ácidos grasos puede ser realizado con emulsiones con altos niveles de fosfolípidos conteniendo ácidos grasos poliinsaturados especialmente con ácido eicosapentanóico (EPA, 20:5 n-3) y docosahexanóico (DHA, 22:6 n-3). La necesidad de ácidos grasos esenciales para la construcción y renovación de membranas es especialmente elevada durante el rápido crecimiento en los estadios de larvas y postlarvas de los peces que pueden exceder la capacidad de síntesis endógena. Por eso para atender esos requerimientos en postlarvas altriciales, son ofrecidos alimentos enriquecidos con ácidos esenciales, aumentando la tasa de crecimiento, la sobrevivencia y la resistencia al estrés.
Entre las vitaminas estudiadas para los peces, las vitaminas A, D, E y C se destacan por estar íntimamente asociadas al desempeño del sistema inmunológico (45). La vitamina C recibe mayor atención por no ser sintetizada por la mayoría de las especies de peces. Por su modo de acción esta involucrada en varias funciones fisiológicas inclusive el crecimiento, desarrollo, reproducción, cicatrización, respuesta al estrés entre otros procesos, gracias a ser un buen agente redactor. Los derivados de ácido ascórbico formados por esteres de fosfatos, el ascorbil monofosfato (AMP) y ascorbil polifosfato (AP) son los más usados como fuente de vitamina C debido a su mayor estabilidad (46).
El desarrollo tecnológico y las fronteras del conocimiento son impulsadas por la investigación y teniendo en cuenta que en nuestro medio se estudian y utilizan especies de peces tropicales con selectividad por ciertos grupos de zooplancton, es necesario realizar estudios que permitan conocer los aspectos más relevantes de diferentes especies de zooplancton, con énfasis en cladóceros y copépodos. Para ello se debe determinar bajo condiciones controladas para una población de organismos, los principales aspectos reproductivos (tiempo de generación, fecundidad, frecuencia reproductiva, periodo fértil, infertilidad juvenil, infertilidad senil y tiempo de incubación). Esto con el fin de establecer si la especie cumple con las características necesarias que permitan generar tecnología de manejo con fines de cultivo masivo, que permitan asegurar el suministro en cantidad y el manejo en la calidad de la primera alimentación de especies de peces neotropicales.
Actualmente se desarrollan en el país diversas investigaciones orientadas a generar alternativas con organismos de zooplancton para el manejo de la larvicultura de especies neotropicales. Estos se enfocan principalmente al estudio del potencial acuícola de especies de zooplancton de agua dulce y marina. Entre los rotíferos se reportan estudios sobre los aspectos reproductivos de Brachionus patulus (47), técnicas para su cultivo (48) y uso como presa viva en larvicultura de peces (18). Actualmente se desarrollan trabajos para la obtención de cuatro cepas de diferente tamaño (100 250 µm) de agua dulce y marinas. En cladóceros se reportan trabajos sobre aspectos reproductivos de Moina sp cepa Ciénaga de Lorica, manejo para su cultivo en sistemas controlados (49) y uso como presa viva en larvicultura de bocachico (18) y bagre rayado (50). En igual sentido, cepas regionales de copépodos de agua dulce y marina, calanoides y harpaticoides, así como, ciclopoides (Cyclopina sp) de agua de mar, estan siendo investigadas para determinar su potencial como alimento vivo en larvicultura y sus condiciones de manejo para generar tecnología de cultivo.
REFERENCIAS
1. Zimmermann S, Jost HC. Recentes Avanços na Nutrição de Peixes: a Nutrição por Fases em Piscicultura Intensiva. In: Memorias de Simpósio sobre manejo e Nutrição de Peixes; 1998, Piracicaba. Anais; Piracicaba- SP, 1998. p. 123-62. [ Links ]
2. Portella MC, Tasser MB, Jomori RK, Carneiro DJ. Substituição do Alimento Vivo na Larvicultura. In: Memorias de Simpósio Brasileiro de Aqüicultura, 2002, Goiânia-Go. Anais; Goiânia: ABRAQ, 2002. [ Links ]
3. Kolkovski S. Digestive enzimes in fish larvae and juveniles-implications and applications to formulated diets. Aquaculture 2001; 200(1/2): 181-201. [ Links ]
4. Tesser MB. Desenvolvimento do trato digestório e crescimento de larvas de pacu, Piaractus mesopotamicus (Holmberg, 1887) em sistemas de co-alimentação com náuplios de artemia e dieta microencapsulada. Dissertação [Tesis de Maestria] Universidade Estadual Paulista; Centro de Aqüicultura, Jaboticabal: 2002. [ Links ]
5. Sipaúba-Tavares LH, Rocha O. Produção de plâncton (Fitoplâncton e Zooplâncton) para alimentação de organismos aquáticos. São Carlos (Bra): RIMA; 2003. p.106. 20-24. [ Links ]
6. Cestarolli MA, Portella MC, Rojas ET. Efeito do nível de alimentação e do tipo de alimento na sobrevivência e no desempenho inicial de larvas de Curimbatá Prochilodus scrofa (Steindachner, 1881). Boletim Instituto de Pesca, São Paulo. 1997; 24: 119-29. [ Links ]
7. Portella MC, Verani JR, Cestarolli MA. Use of live and artificial diets enriched with several fatty acid sources to feed Prochilodus Scrofa larvae and fingerlings. Effects on survival and growth rates, New Delhi. Journal of Aquaculture in the Tropics 2000; 15: 45-58. [ Links ]
8. Atencio-García VJ, Kerguelen E. Wadnipar L, Narvaez A. Manejo de la primera alimentación del bocachico (Prochilodus magdalenae). Rev MVZ Córdoba 2003a; 8(1): 254-60. [ Links ]
9. Prieto Guevara MJ, Rosa Logato PV, Ferreira de Moraes G, Del Okamura, Guedes de Araújo F. Tipo de alimento, sobrevivência e desempenho inicial de Pós-larvas de pacu (Piaractus mesopotamicus), Larvas-Brasil. Ciência e Agrotecnologia; 2006a; 30(5): 1002- 1007. [ Links ]
10. Jomori RK, Carneiro DJ, Malheiros EB, Portella MC. Growth and survival of pacu Piaractus mesopotamicus (Holmberg, 1887) juveniles reared in ponds or at different initial larviculture periods indoors. Aquaculture 2003; 221(1/4): 277-87. [ Links ]
11. Atencio-Garcia VJ, Zaniboni-Filho E, Pardo-Carrasco SC, Arias-Castellanos A. Influência da primerira alimentação na larvicultura e alevinagem do yamú Brycon siebenthalae (Characidae). Maringá, Brasil. Acta Scientiarum. Animal Sciences. 2003b; 25(1): 61-72. [ Links ]
12. Lamadrid J, Arroyo A. Evaluación del régimen alimentario del bocachico Prochilodus magdalenae (Steindachner, 1878) en el alevinaje. [Trabajo de Pregrado] Montería, Colombia: Universidad de Córdoba; 2005. [ Links ]
13. Ramos J, Trujillo C. Evaluación del régimen alimentario de la dorada (Brycon sinuensis Dahl, 1955) en su fase de alevinaje en estanques en tierra. [Trabajo de Pregrado]. Montería, Colombia: Universidad de Córdoba; 2006. [ Links ]
14. Padilla J, Torres J. Régimen alimentario del blanquillo (Sorubim cuspicaudus) en fase de alevinaje en estanques en tierra. [Trabajo de Pregrado] Montería, Colombia: Universidad de Córdoba; 2005. [ Links ]
15. Kubitza F. Nutrição e alimentação dos peixes cultivados. Campo Grande, Mato Grosso do Sul: Projeto Pacu/Agropeixe, 1998. [ Links ]
16. Rabe J, Brown JA. A pulse feeding strategy for rearing larval fish: an experiment with yellowtail flounder. Aquaculture 2000; 191(4): 289-302. [ Links ]
17. Alcalá A, Ortega A. Influencia de la densidad de siembra y la alimentación en la larvicultura del bagre blanco Sorubim cuspicaudus (Littmann, Burr & Nass, 2000). [Trabajo de Pregrado]. Monteria, Colombia: Universidad de Córdoba; 2002. [ Links ]
18. Escobar MA, Prieto MJ, Morales MA. Efecto de alternativas nutricionales en el desempeño de postlarvas de bocachico Prochilodus magdalenae. In: Memorias de II Congreso Colombiano de Acuicultura. Villavicencio, Colombia: 2004. p. 86-87. [ Links ]
19. Lavens P, Sorgeloos P. Introdution. In : Manual on the production and use of life food for aquaculture.Fisheries Technical. Ghent, Belgium: FAO 1996. [ Links ]
20. Coutteau P, Sorgeloos P. Manipulation of dietary lipids, fatty acids and vitamins in zooplankton cultures. Freshw Biol 1997; 38(3): 501-512. [ Links ]
21. Hagiwara A, Gallardo WG, Assavaaree M, Kotani T, Araujo AB. Live food production in Japan: recent progress and future aspects. Aquaculture 2001; 200: 111-127. [ Links ]
22. Blair T, Castell J, Neil S, D'abramo L, Cahu C, Harmon P, et al. Evaluation of microdiets versus live feeds on growth survival and fatty acid composition of larval haddock (Melanogrammus aeglefinus). Aquaculture 2003; 225: 451-461. [ Links ]
23. Shirota A. Studies on the mouth size of fish larvae. Bull Jpn Soc Sci Fish 1970; 36: 353-68. [ Links ]
24. Yúfera M, Fernádez C, Pascual E. Prey-size selectivity of gilthead sea bream larvae (Spaurus aurata L.) fed on microcapsules and live food. In: Pittman K, Batty R, Verreth J. (eds.). Mass rearing of juvenile fish. ICES Mar. 1995; 201-206. [ Links ]
25. Werner E, Hall D. Optimal foranging and the selection of prey by the bluegill sunfiish (Lepomis macrochirus). Ecology 1974; 55: 1042-052. [ Links ]
26. Støttrup JG. The elusive copepods: their production and suitability in marine aquaculture. Aquac Res 2000; 31(8/9): 703-11. [ Links ]
27. Mckinnon AD, Duggan S, Nichols PD, Rimmer MA, Semmens G, Robino B. The potential of tropical paracalanid copepods as live feeds in aquaculture. Aquaculture 2003; 223(1/4): 89-106. [ Links ]
28. Ferrão-Filho AS, Fileto C, Lopes NP, Arcifa MS. Effects of essential fatty acids and N and P-limited algae on the growth rate of tropical cladocerans. Freshw Biol 2003; 48: 759-67. [ Links ]
29. Fregadolli CH. Estudo comparativo do comportamento alimentar de larvas de pacu Piaractus mesopotamicus (Holmberg, 1887) e Tambaqui Colossoma macropomum (Cuvier,1818) em laboratório. 1990. Dissertação. (Tesis de Maestria) Salvador BA. Universidade Federal da Bahia. [ Links ]
30. Pelli A, Dumont-Neto R, Silva J, Gonçalves S, Souza D, Barbosa N. Início de ingestão de ração por pacú (Piaractus mesopotamicus Holmberg, 1887), curimba (Prochilodus scrofa Steinchdacner, 1881) e piau (Leporinus friderici Bloch, 1794). Em condições de criação semi-intensiva. In: Memorias de Simpósio Brasileiro de -Aqüicultura, 9. 1996, Sete Lagoas. Sete Lagoas: ABRAQ, 1996. p. 88 (Resumos). [ Links ]
31. Portella MC, Cestarolli MA, Verani JR, Rojas NE. Produção de organismos planctônicos para alimentação inicial de larvas de peixes de água doce. Boletim Instituto de Pesca, São Paulo (Bra) 1997; 24: 79-89. Único. [ Links ]
32. Reiriz L, Nicieza A, Braña F. Prey selection by experienced and naive juvenile Atlantic salmon. J Fish Biol 1998; 53: 100-14. [ Links ]
33. Sorgeloos P, Dhert A, Candreva P. Use of the brine shrimp, Artemia spp., in marine fish larviculture. Aquaculture 2001; 200(1/2): 147-59. [ Links ]
34. Tamaru CS, Ako H, Pang L. Enrichment of artemia for use in freshwater ornamental fish production. CTSA Publication 1999;(133). [ Links ]
35. Han K, Geurden I, Sorgeloos P. Enrichment strategies for Artemia using emulsions providing different levels of n y 3 highly unsaturated fatty acids. Aquaculture 2000; 183(3/4): 335-47. [ Links ]
36. Lim LC, Cho YL, Dhert P, Wong CC, Nelis H, Sorgeloos P.Use of decapsulated Artemia cysts in ornamental fish culture. Aquac Res 2002; 33(8): 575-89. [ Links ]
37. Nanton DA, Castell JD. Mass culture of the Harpacticoid copepod, Tisbe sp. ICES CM Marine fish culture committee 1997: 10. [ Links ]
38. Manaffar R, Abtahi B. Enrichment of artemia urmiana naupllii with fatty acid emulsion and unicellular algae (Dunaliella tertiolecta) and effects on survival rate and biometry of naupllii under cold incubation. Book of Abstracts. J World Aquac Soc 2003. [ Links ]
39. Merchie G. Use of nauplii and meta-nauplii of Artemia. In: Lavens P, Sorgeloos P. (Ed.). Manual on the production of live food. Aquaculture. 1996. p.137-63. [ Links ]
40. Kolkovski S, Czesny S, Yackey C, Moreau R, Cihla F, Mahan D. The efect of vitamins C and E in (n-3) highly unsaturated fatty acids-enriched Artemia nauplii on growth, survival, and stress resistance of fresh water walleye Stizostedion vitreum larvae. Aquacult Nutr 2000; 6(3): 199-206. [ Links ]
41. Gomez-Gil B, Roque A, Turnbull JF. The use and selection of probiotc bacteria for use in the culture of larval aquatic organisms. Aquaculture 2000; 191: 259-70. [ Links ]
42. Coutteau P, Geurden I, Camara MR, Bergot P, Sorgeloos P. Review on the dietary effects of phospholipids in fish and crustacean larviculture. Aquaculture 1997; 155(1/4): 149-64. [ Links ]
43. Narciso I, Pousão-Ferreira P, Passos A, Luis O. HUFA content and DHA/EPA improvements of Artemia sp. with commercial oils during different enrichment periods. Aquac Res 1999; 30(1): 21-4. [ Links ]
44. Koven W, Barr Y, Lutzky S, Ben-Atia I, Weiss R, Harel M, et al. A The effects of dietary arachidonic acid (20:4 n-6) on growth, survival and resistance to handling stress in gilthead seabream (Sparus aurata) larvae. Aquaculture 2001; 193(1/2): 107-22. [ Links ]
45. Lazo J. Conocimiento actual y nuevas perspectivas en el desarrollo de dietas para larvas de peces marinos. In: Cruz -Suárez LE, Ricque-Marie D, Tapia-Salazar M, Olvera-Novoa MA, Civera-Cerecedo R. (Eds.). Avances en Nutrición Acuícola V. Memorias del V Simposium Internacional de Nutrición Acuícola. 19-22 Noviembre, 2000. Mérida, Yucatán, Mexico 2000. [ Links ]
46. Andersen Friede, Lygren Bjarte, Maage Amund, Waagbø Rune. Interaction between two dietary levels of iron and two forms of the ascorbic acid and the effect on growth, antioxidant status and some non-specific immune parameters in Atlantic salmon (Salmo salar) smolts. Aquaculture 1998; 161: 437-51. [ Links ]
47. Prieto Guevara MJ, Hernandez Padilla AC, Martinez Mena AL. Rotifero Brachionus patulus e seu potencial na aquicultura In: Memorias Anais do XII Simpósio Brasileiro de Aquicultura 2002; Goiânia: CAUNESP/USP/ESALQ, 2002. [ Links ]
48. Barguil D, Lora LG, Prieto Guevara MJ, Morales MA, Marinez A, Hernandez A. Efecto de la densidad inicial de siembra en el cultivo del rotífero Brachionus patulus. In: memorias de II Congreso Colombiano de Acuicultura- X Jornada de Acuicultura-IALL. Villavicencio (Col); 2004 [ Links ]
49. Prieto Guevara MJ, De la Cruz L, Morales MA. Cultivo experimental del cladocero Moina sp alimentado con Ankistrodesmus sp y Saccharomyces cereviseae. Rev MVZ Córdoba 2006; 11(1): 1-16. [ Links ]
50. Barros Angel, Martinez Arlinton. Alternativas nutricionales en la larvicultura del bagre rayado (peudoplatystoma fasciatum; linnaeus 1766). [Trabajo de pregrado] Barranquilla (Col); Universidad del Atlántico; 2006. [ Links ]

