










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista MVZ Córdoba
Print version ISSN 0122-0268On-line version ISSN 1909-0544
Rev.MVZ Cordoba vol.14 no.3 Córdoba Sept./Dec. 2009
ORIGINAL
FEEDING HABITS OF MAYUPA (Sternopygus macrurus Bloch & Schneider, 1801) IN THE SINU RIVER, COLOMBIA
Charles Olaya-Nieto,* M.Sc, Pedro Soto-Fernández, Acuicult, Javier Barrera-Chica, Acuicult.
Universidad de Córdoba. Facultad de Medicina Veterinaria y Zootecnia. Departamento de Ciencias Acuícolas. Laboratorio de Investigación Biológico Pesquera-LIBP. Lorica, Colombia.
Correspondencia: charles_olaya@hotmail.com
Recibido: Diciembre 11 de 2008; Aceptado: Julio 18 de 2009
Resumen
Objetivo. Estudiar los hábitos alimentarios de la Mayupa (Sternopygus macrurus Bloch & Schneider, 1801) en el río Sinú, Colombia. Materiales y métodos. El contenido estomacal se evaluó con el coeficiente de vacuidad, grado de llenado, grado de digestión, frecuencia de ocurrencia, frecuencia numérica, gravimetría, indice de importancia relativa (IIR) y la relación longitud intestinal (LI)-longitud total (LT). Resultados. Solo pocos estómagos se encontraron vacíos (CV =6.9%) y el 60.0% de las presas se encontraron frescas. Se identificaron cuatro ítems alimentarios: peces, crustáceos, material vegetal y otros. Peces fue el ítem más frecuente (76.9%), abundante (48.3%), con mayor composición por peso (81.9%) y con mayor importancia relativa (63.2%). Conclusiones. Los resultados obtenidos permiten inferir que la Mayupa es un pez de hábitos alimentarios carnívoros, con preferencia por los peces.
Palabras clave: Sternopygus macrurus, hábitos alimentarios, río Sinú, Colombia.
Abstract
Objective. To study the feeding habits of Mayupa (Sternopygus macrurus Bloch & Schneider, 1801) in the Sinu river, Colombia. Materials and methods. The stomach contents were analyzed using the proportion of empty stomachs, grade of digestion, frequency of occurrence, numerical frequency, gravimetry, relative importance index (RII) and the gut length (GL)- total length (TL) relationship. Results. Only few stomachs were empty (VC =6.9%) and the 60.0% of preys were fresh. Four food items were identified: fishes, crustaceans, vegetable raw and others. Fishes was the group most frequent (46.5%), abundant (44.7%), with the greatest composition in weight (81.9%) and with greatest relative importance (63.2%). Conclusions. The results indicates that Mayupa is a carnivorous feeding habits fish, with preference by fishes.
Key words: Sternopygus macrurus, feeding habits, Sinu river, Colombia.
INTRODUCCIÓN
La Mayupa (Sternopygus macrurus Bloch & Schneider, 1801) es un Gymnotiforme perteneciente a la familia Sternopygidae, que se encuentra ampliamente distribuida en los ríos Sinú, Magdalena (1), Catatumbo (2) y en toda la Amazonia (3); mostrando relevancia ecológica debido a que hace parte de las relaciones tróficas de muchos ecosistemas, aunque tiene baja importancia comercial en la cuenca del río Sinú.
Se caracteriza por tener un cuerpo largo descamado (4), la aleta anal es muy alargada y utilizada en su propulsión mediante movimientos ondulatorios, las aletas caudal y adiposa no están presentes (2). La cabeza presenta una fontanela dorsal, la mandíbula inferior es más corta que la superior, borde del ojo libre, dientes villiformes (1), y el ano se localiza en el tercio anterior del cuerpo y a veces se desplaza hacia adelante con la edad, pudiendo alcanzar hasta el mentón (5-7).
En el territorio colombiano se encuentra en las grandes cuencas, desde sus partes bajas hasta los 1000 m.s.n.m. (cuencas transandinas) (2) y por lo general viven en el fondo de los ríos, específicamente en el canal principal, por lo que su alimentación es abundante durante todas las épocas del año (8).
Los peces de este género alcanzan la madurez sexual al año de edad, presentándose un tipo de dimorfismo sexual, en donde las hembras maduras tienen mayores descargas eléctricas que los machos en el mismo estado (7).
El objetivo de este trabajo fue establecer los hábitos alimentarios de la Mayupa en la cuenca del río Sinú, Colombia.
MATERIALES Y MÉTODOS
Obtención de especímenes. Se analizaron 335 estómagos de individuos capturados en el bajo río Sinú, entre enero y diciembre 2004. Las capturas se efectuaron a diferentes horas del día, en varios sitios, encontrándose individuos de diferentes tallas. A cada individuo se le midió longitud total (LT) al milímetro más cercano con un ictiómetro graduado y el peso total (WT) al gramo más cercano con una balanza eléctrica Ohaus de 5 kg (± 1 g) de capacidad.
Extracción de los estómagos. Aplicando las técnicas de Laevastu (8) y Marrero (9), una vez efectuada la disección de los peces se ubicaron las diferentes partes del tubo digestivo (esófago, estómago, intestino, y otros). Luego se procedió a retirar el estómago para conservarlo en frascos con formol al 10% bufferado. Cuando un estómago fue muy grande se le inyectó formol para detener la actividad enzimática. Todos los frascos se rotularon, indicando la especie, el número de la muestra, fecha. En un formato se colocó la información citada arriba más el sitio de captura, arte de pesca usado, talla, peso, sexo y estado de madurez sexual.
Análisis del contenido estomacal. El contenido estomacal se colocó en una caja Petri y se examinó al estereoscopio y microscopio, separando, identificando y enumerando el alimento o presas presentes. La identificación se efectuó hasta el nivel taxonómico permitido por el grado de digestión del alimento, agrupándose en presas, ítems o categorías, pesándose en una balanza eléctrica Ohaus de 1500 g (± 0.01 g) de capacidad. El material animal que estaba totalmente digerido se identificó por los fragmentos, en lo posible (10).
El coeficiente de vacuidad (CV) se obtuvo con la técnica de Windell (11):
CV= 100* No. estómagos vacíos / No. total de estómagos analizados.
El grado de digestión (GD) se evaluó con la escala de Laevastu (8), la cual clasifica el estado de las presas así: fresco, medio digerido y digerido.
Se utilizaron 3 métodos para cuantificar el contenido estomacal, expresado en valores promedios mensuales y anuales: frecuencia de ocurrencia (FO), frecuencia numérica (FN) y gravimetría (G) (11-13):
FO= 100* Ocurrencia de presas del ítem A/ No. total de estómagos con alimento
FN= 100* No. de presas del ítem A/No. total de presas
G= 100* Peso de las presas del ítem A/Peso de todas las presas
Para establecer la importancia de cada presa en la composición de la dieta se estimó el índice de importancia relativa (IIR) (14) modificado por Olaya-Nieto et al (15). En este índice, la relación matemática es la siguiente:
IIR = F*G/1OO
En donde, IIR representa el índice de importancia relativa, F es el porcentaje de la frecuencia de ocurrencia y G es el porcentaje gravimétrico. Esta expresión es porcentual presentando un rango de 0 a 100, donde el rango evaluativo del 0 a 10% representa grupos tróficos de importancia relativa baja, de 10 a 40% grupos de importancia relativa secundaria y 40 a 100% grupos de importancia relativa alta.
De igual forma, se observaron las preferencias alimentarias de acuerdo con las tallas que presenta la especie en estudio y con el ciclo hidrológico del río Sinú, y se estableció la relación longitud intestinal/ longitud total de acuerdo con la escala de Brusle (16), la cual plantea lo siguiente: 0.5 a 0.7, planctófagos; 0.5 a 2.4, carnívoros; y 3.7 a 6.0, herbívoros.
Análisis estadístico. Se utilizó estadística descriptiva, expresando las variables como media ± desviación estándar. También se estimó el coeficiente de correlación (r) y/o determinación (r2) entre las variables estudiadas.
RESULTADOS
Se analizaron 335 estómagos de peces colectados entre enero y diciembre 2004, cuyas tallas y pesos oscilaron entre 39.6 y 112.0 cm de longitud total y 132.0 y 1748.0 g, de los cuales 162 individuos fueron machos, 161 hembras y 12 indiferenciados. Se identificaron 4 ítems alimentarios: peces, crustáceos, material vegetal y otros.
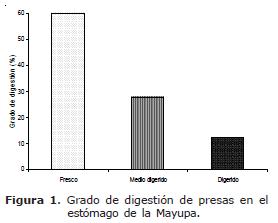
Solo el 6.9% de los estómagos analizados se encontró vacío, especialmente en abril (13.8%), agosto (10.7%) y octubre (10.7%) con los valores máximos. El 60% del alimento consumido se encontró fresco, el 27.8% medio digerido y el 12.2% digerido (Figura 1).
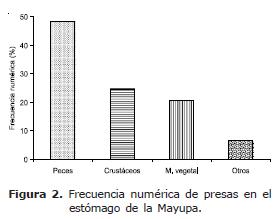
Los ítems alimentarios encontrados presentaron la siguiente frecuencia anual: peces, presente en todos los meses, mostró la mayor ocurrencia con (76.9%), especialmente en julio (91.7%) y diciembre (89.7%), representado por Cocobolo Aequidens pulcher (43.1%), Yalúa Cyphocharax magdalenae (25.5%), restos de peces (escamas, espinas, estructuras esqueléticas y musculares) (21.5%), Poté Dormitator sp. (4.2%), Sardina Astyanax sp. (3.5%), Chipe Hoplosternum magdalenae (1.5%), Mayupa (Sternopygus macrurus) (0.4%); crustáceos (42.0%), conformado por Camarón (Macrobrachium sp.), Jaiba (Callinectes sp.) y restos de crustáceos (músculo y apéndices), presente en todos los meses, principalmente en febrero (63.0%) y agosto (52.0%); material vegetal (40.1%), constituido por semillas y raíces, también presente en todos los meses, especialmente en enero (68.2%) y abril (64.0%) y otros (11.5%) integrado por bivalvo, carbón, nemátodo, nylon, papel aluminio, pelo, piedra, plástico, serpiente y vidrio, presente en todos los meses, excepto noviembre.
La participación de cada grupo alimentario mostró a peces (Figura 2) como el más abundante (48.3%), conformado por Cocobolo (45.0%), destacándose el alto consumo en junio, julio y noviembre; Yalúa (27.2%), restos de peces (19.0%), Poté (3.7%), Sardina (3.4%), Chipe (1.4%) y Mayupa (0.3%); seguido por Crustáceos (24.5%), material vegetal (20.6%) y otros (6.6%).
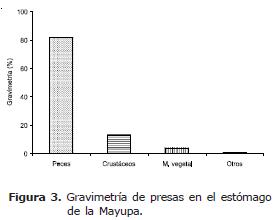
El análisis gravimétrico mostró que el 81.9% del total de presas consumidas durante el año son peces, seguido por crustáceos (13.2%), material vegetal (4.0%), mientras que otros apenas registró el 0.9% (Figura 3). Dentro del ítem peces, la especie que alcanzó el mayor valor fue Cocobolo (51.4%), seguida por Yalúa (37.4%), restos de peces (4.5%), Sardina (2.7%), Poté (2.2%), Chipe (1.7%) y Mayupa (0.1%).
El indice de importancia relativa (IIR) mostró a peces como un alimento de alta importancia (63.2%) dentro de la dieta de la Mayupa. Los demás ítems como crustáceos (5.5%) y material vegetal (1.6%) y otros (0.1%) se consideran de baja importancia e incidentales, respectivamente. La relación longitud intestinal (LI)-longitud total (LT) se estimó en 0.6, lo que ubica a la especie en estudio como carnívora, teniendo en cuenta la escala propuesta por Brusle (16).
Para el análisis de las preferencias alimentarias de acuerdo con la talla, los ejemplares de la especie en estudio se agruparon en ocho intervalos de tallas, en donde se puede observar que peces no aparece en el intervalo menor (35.0–45.0 cm LT) y participa con alrededor del 50% en la dieta de la Mayupa en los demás intervalos, excepto (35.0-45.0 y 75.0-85.0 cm LT); crustáceos es el ítem principal (50.0%) en el intervalo menor (35.0–45.0 cm LT), alcanzando valores cercanos al 25.0% en el resto; material vegetal mantiene una participación entre 19.1 y 25.0%; mientras otros decrece con el tamaño, pasando de 25.0% (35.0–45.0 cm LT) a 0.0% en los intervalos mayores (95.0–105.0 y 105.0-115.0 cm LT).
DISCUSIÓN
El coeficiente de vacuidad encontrado en este trabajo fue similar al reportado para un congénere como el Caloche (Sternopygus aequilabiatus) en la Represa de Prado (7.7%), Colombia (17) y menor que el estimado en el Embalse de Betania (0.0%), Colombia (18).
La literatura revisada plantea que los peces, en especial los carnívoros, presentan alto porcentaje de estómagos vacíos (19) y de mayor volumen que los herbívoros y omnívoros (20), en donde el fenómeno de la regurgitación está asociado con su estrategia de defensa y escape (21). Dicho fenómeno puede ser estimulado también por el uso de anzuelos y redes agalleras en la captura de los individuos estudiados (22). En contraste, la Mayupa presenta alto porcentaje de estómagos con alimento, lo que permite inferir que no adopta este tipo de estrategia.
El buen estado de las presas encontradas facilitó la identificación de las mismas, puesto que la mayoría estaban enteras, lo que permite suponer que fue capturada en horas muy asociadas con su alimentación, lo que puede ser confirmado con la alta presencia de presas en estado fresco. Aunque, Smith (23) reporta que la mayoría de los organismos hidrobiológicos con estómago verdadero presentan células productoras de ácido clorhídrico, de tal manera que el pH del fluido gástrico puede bajar de 2 y suele fluctuar entre 2 y 5, según el tiempo transcurrido después de una comida. Lo anterior deja de manifiesto que el estómago de la Mayupa posiblemente presente un pH no muy ácido para digerir con rapidez las presas ingeridas, de allí la gran cantidad de alimentos frescos y medio digeridos encontrados en los estómagos analizados durante el periodo de estudio.
En torno a la frecuencia de ocurrencia, Villa-Navarro & Losada-Prado (17) reportaron valores de 56.0 y 20.0% para material vegetal y peces, respectivamente. Galvis et al (2) observaron el consumo de pequeños crustáceos, Villa-Navarro & Losada-Prado (17) encontraron que peces alcanza un valor de 67.7% y material vegetal 7.5%, coincidiendo con los resultados obtenidos en esta investigación en el análisis gravimétrico; y Rotta (20) concluyó que la longitud del intestino parece estar más relacionada con la cantidad de material indigerible del alimento que con su origen animal y/o vegetal.
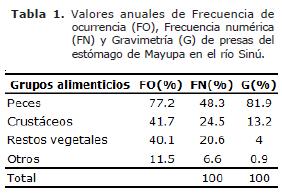
Analizando la composición de la frecuencia de ocurrencia, frecuencia numérica, gravimetría (Tabla 1), se observó la prevalencia del grupo peces sobre los demás grupos alimentarios, lo que sumado a la información arrojada por el IIR y la relación LI-LT permite inferir que la especie en estudio tiene una dieta basada principalmente en Peces, lo que indica que sus hábitos alimentarios son carnívoros con tendencia a la ictiofagia.
Al desagregar la información de este índice en cuanto a peces se refiere, debido a que fue el más importante en el régimen alimentario de la especie en estudio, se observó que la presa con mayor importancia relativa (IIR) fue Cocobolo (22.1%), seguida por Yalúa (9.6%), las demás presas se caracterizaron por presentar una baja importancia debido a que sus valores son menores del 1% (Tabla 2).
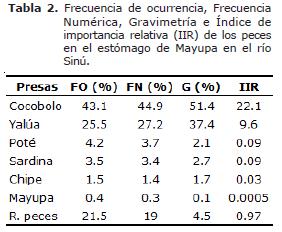
Se observó que la frecuencia y abundancia de Cocobolo son mayores en junio, julio y noviembre; mientras que para Yalúa fue en febrero, abril y mayo; lo cual puede estar asociado a que la Mayupa es un pez oportunista y poco selectivo alternando su dieta de acuerdo con la disponibilidad de las presas en el ambiente o en su entorno, ya sea en el cauce principal o en los planos inundables.
En la Tabla 3 se presenta la composición y tamaño de las presas consumidas por la Mayupa, enfatizando en el ítem peces por ser la base de su dieta. En el intervalo (45.0- 55.0 cm), Yalúa (50.0%) es la principal especie consumida, seguida por Cocobolo (25.0%); en los cuatro intervalos siguientes (55.0-65.0 cm, 65.0-75.0 cm, 75.0-85.0 cm y 85.0-95.0 cm) Cocobolo es la principal especie consumida (64.8, 58.0, 46.2 y 59.5%, respectivamente); en el próximo (95.0-105.0 cm) estas dos especies alcanzan el 40.0% cada una; y en el último (105.0-115.0 cm) la Yalúa vuelve a ser la especie preferida con el 100.0%, aunque sólo se colectó un ejemplar.
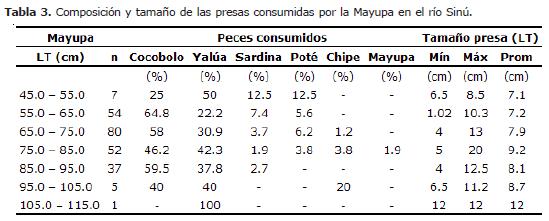
La talla de la Mayupa osciló entre 39.6 y 112.0 cm LT, mientras que la talla de los diferentes peces consumidos se encontró entre 1.02 y 20.0 cm LT, por lo que se puede inferir que la especie en estudio no consume peces de tallas mayores a 20.0 cm LT. La relación talla media de la presa/talla media del predador mostró valores alternantes a medida que el pez va creciendo (14.2, 12.0, 11.3, 11.5, 9.0, 8.7 y 10.9%), observándose que sus presas no alcanzan el 15% de su talla, aunque no haya correlación entre ellas. Analizando la talla máxima de peces consumidos con la talla del predador, se observó que la talla máxima que puede consumir es del 23.5%.
Mancera-Rodríguez & Cala (18) afirman que la especie basa su dieta en el consumo de peces; Galvis et al (1) también encontraron consumo de pequeños crustáceos y Villa- Navarro & Losada-Prado (17) reportan que peces y material vegetal son los ítems preferenciales de la Mayupa en la Represa de Prado; resultados similares a los obtenidos en esta investigación.
Resende et al (24) reportaron que los peces de ambientes tropicales y subtropicales son eurífagos que consumen -en lo posible- otros alimentos que se encuentran disponibles en el medio como insectos, microcrustáceos y camarones; y que los peces carnívoros, especialmente de los ríos con planicies inundables que funcionan ecológicamente como sistemas de pulsos son carnívoros generalistas, alimentándose con las presas más abundantes y disponibles en el medio ambiente, aplicándose este concepto a la Mayupa.
Los peces carnívoros ingieren sobre todo ítems de origen animal, alimentándose de invertebrados de buen tamaño y peces, especializándose en algún tipo en particular, preferencias que pueden cambiar con la disponibilidad estacional de alimento (20); aunque Hiss et al (25), afirman que la Mayupa es capaz de seleccionar los organismos que consume como una estrategia para evitar enfrentamientos con competidores más eficientes dentro del medio en donde vive.
Aunque solo se encontró un individuo (9.5 cm LT y 4.8 g) en el mes de abril, la presencia de peces de su misma especie en su estómago denota comportamiento caníbal, caso que no había sido reportado hasta el momento. Por canibalismo se entiende al acto de matar y consumir todo o la mayor parte de un individuo procedente de la misma especie, independientemente de su estado de desarrollo, siendo una forma especial de predación que introduce preguntas evolucionarias intrigantes y que tiene un potencial para la regulación de la población dependiente de la densidad, lo cual puede ser muy costoso en la acuicultura (26).
Las especies de peces que presentan esta conducta tienen diversos hábitos alimentarios, desde carnívoro hasta herbívoro y desde los que presentan o no algún tipo de cuidado parental (26), siendo más común el fenómeno en los peces carnívoros y en los estados iniciales del desarrollo (27, 28), al igual que entre los piscívoros, grupo que tiene adaptaciones para la predación, como la gran habilidad para detectar y capturar peces (26). Durante su edad temprana los piscívoros tienen una boca grande y dientes bien formados, que les permiten consumir presas relativamente grandes y continuar con su conducta caníbal (29).
En algunos casos el canibalismo hace una contribución importante a la dieta (estado nutricional) (26) atribuido a menudo a la cantidad o disponibilidad de alimento, aunque la baja calidad del mismo puede generarlo (30-32). La mayoría de los casos se originan cuando los peces grandes predan peces pequeños (disparidad de tallas), relación común en las interacciones predador-presa, la cual es determinada frecuentemente por el tamaño de la boca del predador (33).
En conclusión, los resultados obtenidos permiten inferir que la Mayupa es un pez de hábitos alimentarios carnívoros, con preferencia por los peces.
Agradecimientos
Los autores expresan sus agradecimientos a la Oficina de Investigación y Extensión de la Universidad de Córdoba, por la financiación del proyecto de investigación "Estimación de los parámetros biológicos básicos de peces comerciales del Río Sinú–Fase II", Código FMV-01-04, Numeral 112010, del cual hace parte este trabajo; así como a los pescadores de la cuenca del río Sinú.
Referencias
1. Dahl G. Los peces del norte de Colombia. Bogotá D.C: Inderena; 1971. [ Links ]
2. Galvis G, Mojica JI, Camargo M. Peces del Catatumbo. Bogotá D.C: Asociación Cravo Norte; 1997. [ Links ]
3. Albert JS. Familiy Sternopygidae (Glass Knifefishes, Rattail Knifefishes). In: Reis RE, Kullander SO, Ferraris Jr CJ. (eds.). Checklist of the freshwater fishes of South and Central America. Porto Alegre: Editora da Pontifícia Universidade Católica do Rio Grande do Sul-EDIPUCRS 2003; 487-491. [ Links ]
4. Albert JS. Species diversity and phylogenetic systematics of American knifefishes (Gymnotiformes, Teleostei). Miscellaneous Publications Museum of Zoology, University of Michigan; 2001. [ Links ]
5. Mago-Leccia F. Electric fishes of the continental waters of America. Biblioteca de la Academia de Ciencias Físicas, Matemáticas y Naturales 1994; Vol. XXIX: 1-222. [ Links ]
6. Mago-Leccia F. Los peces de la familia Sternopygidae de Venezuela, incluyendo una descripción de la osteología de Eigenmannia virescens y una nueva definición y clasificación del orden Gymnotiformes. Acta Cient Venez 1978; 29(Supl. 1):1-89. [ Links ]
7. Provenzano F. Aspectos de la reproducción de peces Gymnotifomes del Bajo Llano de Venezuela. Trabajo de ascenso, Facultad de Ciencias, Universidad Central de Venezuela, Caracas; 1984. [ Links ]
8. Laevastu T. Manual de métodos de biología pesquera. Zaragoza: Acribia; 1980. [ Links ]
9. Marrero C. Métodos para cuantificar contenidos estomacales en peces. Universidad Nacional Experimental de los Llanos Ezequiel Zamora-Unellez, Vicerrectorado de Producción Agrícola, Programa de Recursos Naturales Renovables, Museo de Zoología, Guanare; 1994. [ Links ]
10. Lugo RL. Determinación de hábitos, madurez sexual y desove en tres especies íctícas de la cuenca del Río Tomo (Vichada) y consideraciones para el mantenimiento de los padrotes. Villavicencio: Universidad Tecnológica de los Llanos Orientales - I. I. O .C. N° 2; 1989. [ Links ]
11. Windell JT. Food analysis and rate of digestion. In: Ricker WE. (ed.). Methods for assessment of fish production in fresh waters. 2nd edition. Oxford: Blackwell Scientific Publications 1971; 215-226. [ Links ]
12. Windell JT, Bowen SH. Methods for study of fish diets based on analysis of stomach contents. In: Bagenal T. (ed.). Methods for assessment of fish production in fresh waters. 3rd edition. Oxford: Blackwell Scientific Publications 1978; 219-226. [ Links ]
13. Silva M, Stuardo J. Alimentación y relaciones tróficas generales entre algunos peces demersales y el bentos de Bahía Coliumo (Provincia de Concepción, Chile). Gayana Zool 1985; 49(3-4): 77-102. [ Links ]
14. Yáñez-Arancibia A, Curiel-Gómez J, Leyton V. Prospección biológica y ecología del bagre marino Galeichthys caerulescens (Günther) en el sistema lagunar costero de Guerrero, México (Pisces:Ariidae). An Centro Cienc del Mar y Limnol Univ Nal Autón México 1976; 3(1): 125-180. [ Links ]
15. Olaya-Nieto CW, Tobías-Arias AJ, Segura-Guevara F, Brú-Cordero SB, Tordecilla-Petro G. Modificación del índice de importancia relativa (IIR) de Yáñez-Arancibia, Curiel-Gómez & Leyton (1976). Universidad de Córdoba, Facultad de Medicina Veterinaria y Zootecnia, Departamento de Acuicultura, Laboratorio de Investigación Biológico Pesquera-LIBP, Lorica; 2003. [ Links ]
16. Brusle J. Food and feeding in grey mullet. In: Oren OH. (ed.). Aquaculture of grey mullet. Cambridge: Cambridge University Press 1981; 185-217. [ Links ]
17. Villa-Navarro FA, Losada-Prado S. Aspectos bioecológicos del Caloche, Sternopygus macrurus. (Gymnotiformes: Sternopygidae), en la represa de Prado, Tolima, Colombia. Dahlia-Rev Asoc Colomb Ictiol 2004; 7: 49-56. [ Links ]
18. Mancera-Rodríguez NJ, Cala P. Aspectos biológicos de la comunidad íctica asociada a un cultivo de Tilapia roja en jaulas flotantes en el Embalse de Betania, Colombia. Dahlia-Rev Asoc Colomb Ictiol 1997; 2: 31-53. [ Links ]
19. Resende EK de, Pereira RAC, Almeida VLL de, Silva AG da. Peixes onívoros da planície inundável do Río Miranda, Pantanal, Mato Grosso do Sul, Brasil. Embrapa Pantanal Boletim de Pesquisa 2000; 16: 1-44. [ Links ]
20. Rotta MA. Aspectos gerais da fisiología e estructura do sistema digestivo dos peixes relacionados á piscicultura. Embrapa Pantanal Documentos 2003; 53: 1-48. [ Links ]
21. Pérez-Chaparro L, Ajiaco-Martínez R, Ramírez-Gil H. Aspectos biológicos de algunas especies de peces de interés ornamental en la baja Orinoquía colombiana. En: Ramírez-Gil H, Ajiaco- Martinez RE. (eds.). La pesca en la baja Orinoquía colombiana: una visión integral. Bogotá: Instituto Nacional de Pesca y Acuicultura-INPA 2001; 155- 210. [ Links ]
22. Prejs A, Colomine G. Métodos para el estudio de los alimentos y las relaciones tróficas de los peces. Caracas: Universidad Central de Venezuela; 1981. [ Links ]
23. Smith LS. Digestion in teleost fishes. Chapter 1. In: Lectures presented at the FAO/UNDP Training Course in Fish Feed Technology at University of Washington. Rome: FAO; 1980. [ Links ]
24. Resende EK de, Pereira RAC, Almeida VLL de, Silva AG da. Alimentação de peixes carnívoros da planicie inundável do Rio Miranda, Pantanal, Mato Grosso do Sul, Brasil. Embrapa Pantanal Boletim de Pesquisa 1996; 03: 1-37. [ Links ]
25. Hiss JS, Shirley KE, Aristizábal W. La pesca en la Represa de Prado, Tolima, 1974-1978. Bogotá: Cuerpos de Paz; 1978. [ Links ]
26. Smith C, Reay P. Cannibalism in teleost fish. Rev Fish Biol Fish 1991; 1: 41-64. [ Links ]
27. Ricker WE. Stock and recruitment. J Fish Res Bd Can 1954; 11: 559-623. [ Links ]
28. Hecht T, Pienaar A. A review of cannibalism and its implications in fish larviculture. J World Aquac Soc 1993; 24(2): 246-261. [ Links ]
29. Loadman NL, Moodie GEE, Mathias JA. Significance of cannibalism in larval walleye (Stizostedion vitreum). Can J Fish Aquat Sci 1986; 43: 613-618. [ Links ]
30. Polis GA. The evolution and dynamics of intraspecific predation. Annu Rev Ecol Syst 1981; 12: 225-251. [ Links ]
31. Teng SK, Akatsu S, Al-Abdul-Elah KM, El-Zahr CR, Downing N, Al-Marzouk A, Chazal N. Spawning, fingerling production and market-size culture of sobaity (Acanthopagrus cuvieri) in Kuwait. A Res Rep Kuwait Inst Sci Res 1981; 66-70. [ Links ]
32. Smith C. An investigation into the problem of conspecific predation among the fry of the Nile tilapia, Oreochromis niloticus (Linnaeus, 1757) in an intensive culture system. MSc Thesis. Plymouth Polytechnic. Plymouth; 1989. [ Links ]
33. Shirota A. Studies on the mouth size of fish larvae. II. Specific characteristics of the upper jaw length. Bull Japan Soc Sci Fish 1978; 44: 1171- 1177. [ Links ]

