










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista MVZ Córdoba
Print version ISSN 0122-0268On-line version ISSN 1909-0544
Rev.MVZ Cordoba vol.14 no.3 Córdoba Sept./Dec. 2009
ORIGINAL
IN VITRO ASSESSMENT OF NEMATOPHAGUS FUNGI FOR BIOLOGICAL CONTROL OF GASTROINTESTINAL NEMATODES OF RUMINANTS
Martha Orozco A1*, M.Sc, Víctor Álvarez C2, M.Sc, Ana Jiménez R3, Ph.D, Óscar Acuña N4, M.Sc.
1Universidad Nacional de Costa Rica. Escuela de Ciencias Agrarias. Laboratorio de Ecología
de los organismos del suelo y control biológico, Heredia, Costa Rica. 2Universidad Nacional
de Costa Rica. Escuela de Medicina Veterinaria. Laboratorio de Parasitología. Heredia,
Costa Rica. 3Universidad Nacional de Costa Rica. Escuela de Medicina Veterinaria.
Laboratorio de Parasitología, Heredia, Costa Rica. 4Universidad de Costa Rica. Centro de
Investigaciones Agronómicas. Laboratorio de Bioquímica de procesos orgánicos. San José,
Costa Rica. *Correspondencia: morozc@una.ac.cr
RESUMEN
Objetivo. Determinar la actividad depredadora in vitro de cepas de hongos nematófagos autóctonos de Costa Rica. Materiales y métodos. Dieciséis cepas de hongos nematófagos nativos de Costa Rica de los géneros Candelabrella musiformis, Arthrobotrys oligospora y Dactylella sp. Se evaluó in vitro la actividad depredadora de los hongos frente a nemátodos gastrointestinales como Haemonchus spp., Oesophagostomum sp. y Trichostrongylus sp aislados de pequeños rumiantes. Resultados. La actividad de los hongos fue muy heterogénea, presentándose resultados que van desde 9.8 a 99.6% de depredación. Conclusiones. El análisis estadístico demostró diferencias significativas en la actividad de los hongos; sin embargo, las cepas de A. oligospora mostraron un mejor desempeño en la captura de nemátodos comparado con las cepas de C. musiformis y Dactylella sp. La heterogeneidad encontrada responde a factores como: origen de las cepas, características intrínsecas de las mismas y especies de larvas utilizadas.
Palabras clave: Biocontrol, hongos, Artrhobotrys oligospora, nemátodos, rumiantes.
ABSTRACT
Objetive. To determine in vitro predatory activity of fungi strains nematófagos native of Costa Rica. Materials and methods. Sixteen native strains of nematophagus fungi from Costa Rica isolated from different substrata and identified as Candelabrella musiformis, Arthrobotrys oligospora and Dactylella sp. In order to assess their fungi predatory activity, it was evaluated in vitro assay gastrointestinal nemátodos as Haemonchus spp., Oesophagostomum sp. and Trichostrongylus sp, isolated from small ruminants. Results. The activity of fungi was very heterogeneous, ranging between 9.8 and 99.6%. Conclusions. The statistical analysis showed significant differences; however, A. oligospora strains were more effective in capturing nematodes in comparison with C. musiformis and Dactylella sp. strains. Heterogeneity is explained by factors like strains origin, their own characteristics and type of larvae used in the test.
Key words: Biocontrol, fungi, Arthrobotrys oligospora, nematodes, ruminants.
INTRODUCCIÓN
Los últimos treinta años se han caracterizado por el desarrollo y aplicación en distintas áreas del mundo de numerosas estrategias químicas (antiparasitarios o antihelmínticos) para el control de parásitos gastrointestinales y pulmonares en el ganado. Sin embargo, dichas estrategias han provocado una gran presión de selección en las poblaciones parasitarias, produciendo resistencia.
En trabajos llevados a cabo por la FAO a solicitud de la Organización Mundial de Sanidad Animal, se muestra que el 55% de los países miembros de dicha organización (77 de 151 países) tienen problemas de resistencia en especies de endo y ectoparásitos de importancia económica en rumiantes. El 22% de estos países presentan dos o más especies con resistencia (1). Actualmente se ha presentado resistencia a los antiparasitarios en los cinco continentes llegando a un estado de crisis en países como Argentina, Brasil, Uruguay y Paraguay (2). Otros efectos importantes de los desparasitantes químicos son la residualidad en productos de consumo humano y el posible impacto negativo en organismos no blanco (3).
Actualmente el manejo de la sanidad animal en lo que a parasitosis se refiere se ha dirigido hacia el llamado "Combate Integrado de Parásitos" o CIP, cuyo objetivo es combinar adecuadamente varias herramientas de control a efectos de desestabilizar la formación de aquellas poblaciones parasitarias con mayor proporción de individuos genéticamente resistentes, manteniendo un nivel adecuado de producción (1).
El CIP contempla el uso de hongos nematófagos depredadores para el control biológico de nemátodos gastrointestinales y pulmonares en pasturas o suelo. Los hongos nematófagos son microorganismos que atrapan, destruyen y se alimentan de nemátodos vivos en el suelo (4). Pertenecen en su mayoría al grupo de los Hyphomycetes (Deuteromycetes) (5) y se clasifican en dos grandes grupos de acuerdo a su estrategia de captura de nemátodos, estos son: depredadores y endoparásitos. Actualmente el grupo de los hongos nematófagos depredadores es el que ha tenido mayor relevancia en la investigación sobre opciones de control biológico de nemátodos gastrointestinales y pulmonares que afectan rumiantes. Los hongos depredadores se caracterizan por producir un extenso sistema de hifas y a todo lo largo de las hifas a ciertos intervalos presentan estructuras especializadas para atrapar y sostener nemátodos vivos (6). Estas estructuras pueden ser perillas pegajosas, anillos no adhesivos, redes adhesivas, anillos constrictores, entre otros. Algunos ejemplos de hongos depredadores investigados mundialmente son el género Arthrobotrys con varias especies y Duddingtonia flagrans. Estos han presentado resultados variables contra Haemonchus contortus, Teladorsagia circumcincta, Trichostrongylus colubriformis, Cooperia sp., Oesophagostomum sp., Dictyocaulus viviparus, Strongyloides papillosus, entre otros (7-10).
Un primer paso para determinar el potencial de los hongos nematófagos depredadores como controladores biológicos consiste en realizar ensayos in vitro donde éstos son puestos en contacto con los nemátodos gastrointestinales para posteriormente evaluar la disminución en la población de los mismos. Los ensayos in vitro proveen información que ayudará a predecir si un hongo podrá o no ser un depredador eficiente en la práctica, ya que un pobre desempeño en la captura de nemátodos en condiciones óptimas, da la pauta para descartarlo como potencial controlador biológico de nemátodos en el ambiente.
El estudio de los hongos nematófagos ha permitido usar micelios o esporas de hongos nematófagos administradas vía oral para inocular de manera indirecta heces animales y propiciar un contacto estrecho entre larvas recién eclosionadas y los hongos, evitando que las larvas pasen al suelo (11).
El objetivo del presente trabajo fue determinar la actividad depredadora in vitro de cepas de hongos nematófagos autóctonos de Costa Rica, enfrentándolos a nemátodos gastrointestinales de rumiantes para observar su efectividad en la reducción de las poblaciones de este tipo de nemátodos y así seleccionar las cepas más eficientes como candidatos a controladores biológicos.
MATERIALES Y MÉTODOS
Larvas de nemátodos gastrointestinales. Las larvas de nemátodos gastrointestinales (NG) se aislaron de estiércol de ovejas y cabras tomado directamente del recto de los animales. Las ovejas provenían de un sistema productivo de engorde ubicado en el cantón de Jiménez, Cartago y las cabras del proyecto caprino de la Estación Experimental Santa Lucía de la Universidad Nacional en Heredia. Todas las muestras de estiércol fueron procesadas mediante la técnica de flotación de Sheater (12) para observar la presencia de huevos de nemátodos gastrointestinales. Cuando las muestras resultaron positivas y presentaron cargas parasitarias elevadas se realizaron coprocultivos para propiciar la eclosión de los huevos y la liberación de las larvas. Los coprocultivos se establecieron mezclando heces con aserrín y colocando la mezcla en frascos de vidrio, los cuales fueron humedecidos con agua destilada e incubados a 27°C en oscuridad. A los siete días se extrajeron las larvas de tercer estadío de los frascos utilizando la técnica de migración larvaria en agua a 30°C por 2 h. De las larvas extraídas, 3 submuestras fueron tomadas y teñidas con solución de lugol para ser observadas al microscopio compuesto e identificar los géneros presentes. La identificación de los NG fue realizada por personal del Laboratorio de Parasitología Veterinaria de la Universidad Nacional de Costa Rica. La población de larvas de NG provenientes de ovejas consistió en: 51% Haemonchus contortus; 41% Trichostrongylus spp; y 8% Oesophagostomum spp. La población de larvas de NG provenientes de cabras consistió en 100% Haemonchus contortus. El resto de las larvas extraídas fueron desinfectadas con tetraciclina (0.01 g/ml) para ser utilizadas en la determinación de actividad depredadora de los hongos nematófagos a probar.
Es importante señalar que no se trabajó con poblaciones homogéneas de nemátodos ya que resulta complicado establecer cultivos puros de estos organismos debido a su carácter de parásitos obligados.
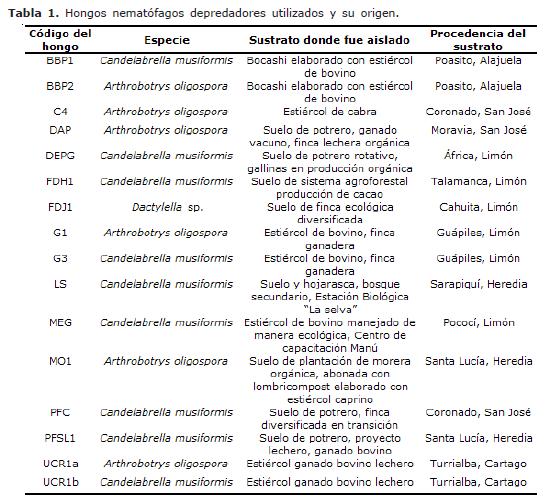
Hongos nematófagos. Como se indica en la tabla 1, dieciséis cepas de hongos nematófagos depredadores aislados a partir de diversos sustratos recolectados en varias zonas de Costa Rica fueron utilizadas en la prueba. Las 16 cepas consistieron en 9 Candelabrella musiformis, 6 Arthrobotrys oligospora y una Dactylella sp. Todos los anteriores producen redes pegajosas bi o tridimensionales que constituyen las estructuras donde los nemátodos son atrapados.
Todos los hongos probados producen esporas asexuales o conidios. Para realizar el ensayo se prepararon suspensiones acuosas de conidios, los cuales se cosecharon mediante el raspado de la superficie del medio papa-dextrosa- agar donde los hongos se sembraron previamente.
Actividad depredadora in vitro. Cajas Petri de 9 cm de diámetro conteniendo agaragua (AA) fueron inoculadas con 9000 - 11000 conidios, se establecieron 13 repeticiones por cepa. Las cajas se incubaron por tres días a temperatura ambiente (23-26°C) y luz artificial (luz del laboratorio). Pasado este tiempo se depositó en cada caja 0,5 ml de una suspensión conteniendo aproximadamente 100 larvas de NG. Las cajas se incubaron por cuatro días más en las condiciones antes descritas. Posteriormente se contó en cada caja el número de larvas libres y atrapadas en las redes pegajosas con la ayuda de estereoscopio. Es importante aclarar que con esta metodología no es posible diferenciar los géneros o especies de los nemátodos libres y atrapados en las trampas; por lo tanto, a pesar de que inicialmente se determinó la composición de la población larvaria, para efectos de análisis y discusión se considera únicamente el número total de larvas aisladas de cabras y ovejas, expresando algunas inferencias.
Se establecieron los siguientes testigos: 1) tres cajas Petri con AA conteniendo sólo larvas de NG, 2) cajas con AA conteniendo únicamente a los hongos (una caja por hongo).
El procedimiento antes descrito se realizó dos veces, uno con larvas de NG procedentes de ovejas y otro con larvas de NG procedentes de cabras.
Evaluación. Con los datos obtenidos se calculó la actividad depredadora en porcentaje por cada caja de la siguiente manera:
A= (x/x + y)/100
Donde:
A = actividad depredadora (%)
X = número de larvas atrapadas
Y = número de larvas libres
Análisis estadístico. El diseño del ensayo fue irrestricto y con los datos obtenidos se realizó un análisis de varianza (ANDEVA) y prueba de separación de medias de Tukey para determinar la presencia de diferencias significativas en la actividad depredadora desarrollada por las 16 cepas. Se realizó el mismo procedimiento para observar diferencias significativas entre cepas de la misma especie. Debido a que la actividad depredadora de los hongos nematófagos sigue una distribución binomial (3), se aplicó la transformación arcosen o angular a los datos con el fin de normalizarlos previamente al análisis estadístico.
RESULTADOS
Actividad depredadora de las 16 cepas. El análisis estadístico indicó diferencias altamente significativas en la actividad depredadora de las 16 cepas de hongos nematófagos (p=0.0001). La separación de medias de Tukey resultó en el establecimiento de 9 grupos cuando los hongos se encuentran capturando larvas de NG procedentes de ovejas y 6 grupos cuando se encuentran capturando larvas de NG procedentes de cabras (Tablas 2 y 3). Es decir, se presentaron actividades de captura más heterogéneas cuando los hongos atraparon larvas de NG procedentes de ovejas.
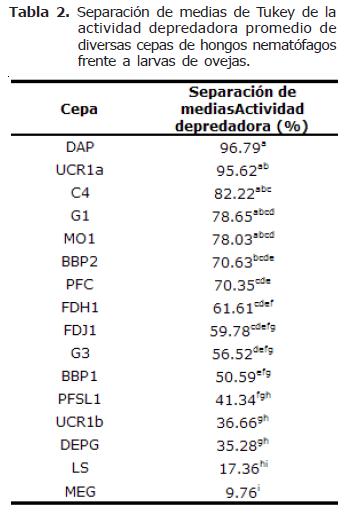
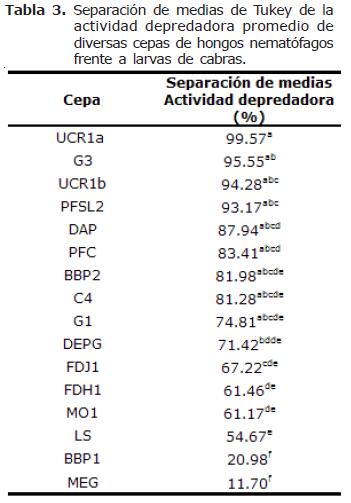
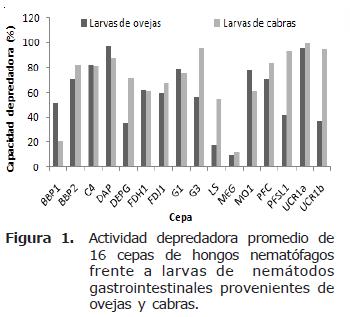
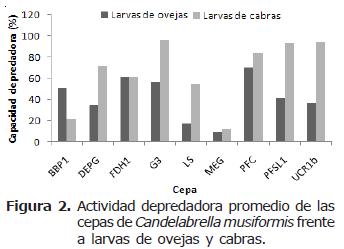
La cepa que presentó la mayor actividad depredadora fue UCR1a (A. oligospora proveniente de estiércol bovino) con un 99.6% y la que presentó la menor actividad depredadora fue MEG (C. musiformis proveniente de estiércol bovino) con 9.8%. Hubo una tendencia de las cepas de A. oligospora de atrapar preferentemente larvas de NG provenientes de ovejas, ya que de las seis cepas probadas de esta especie, cuatro desarrollaron actividades de depredación mayores con dicho tipo de larvas (cepas C4, DAP, G1 y MO1). En contraste, las cepas de C. musiformis tendieron a atrapar larvas de NG provenientes de cabras, ya que de las nueve cepas de esta especie, siete desarrollaron actividades depredadoras más elevadas al atrapar este tipo de larvas (cepas DEPG, G3, LS, MEG, PFC, PFSL1 y UCR1b). La única cepa que no presentó diferencias al atrapar larvas de NG provenientes de ovejas o cabras fue FDH1. Dactylella sp. presentó una ligera preferencia por NG de larvas provenientes de cabras (Figura 1).
Actividad depredadora entre cepas de Candelabrella musiformis. El análisis estadístico indicó diferencias altamente significativas en la actividad depredadora de las nueve cepas de C. musiformis con larvas de NG provenientes tanto de ovejas como de cabras (p=0.0001). La comparación de medias de Tukey determinó la presencia de cinco grupos que se traslapan entre sí cuando los hongos se encuentran capturando larvas provenientes de ovejas y cuatro grupos diferentes cuando se encuentran atrapando larvas provenientes de cabras (Tablas 4 y 5).
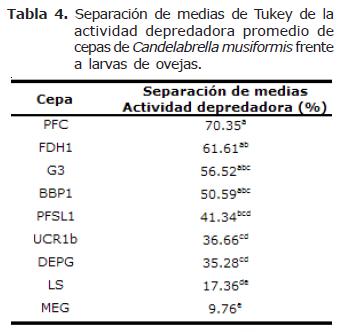
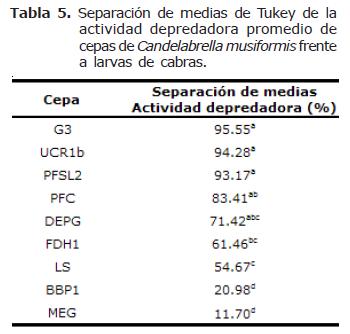
En general, la actividad depredadora de las cepas de C. musiformis fue muy heterogénea, presentándose rangos amplios; con larvas de NG provenientes de ovejas fue de 9.8% a 70.3%, siendo la cepa MEG la menos efectiva y la cepa PFC la más efectiva. Con larvas de NG provenientes de cabras el rango fue de 11.7% de la cepa MEG a 95.5% de la cepa G3. Como se menciona en la sección anterior, las cepas de este hongo presentaron una actividad depredadora más eficiente con larvas provenientes de NG de cabras que de ovejas y en algunos casos esta diferencia fue bastante marcada (cepas DEPG, G3, LS, PFSL1 y UCR1b) (Figura 2).
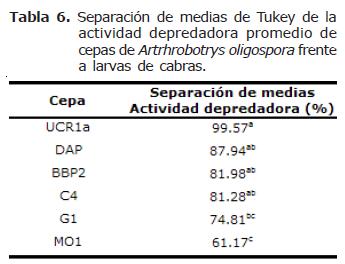
Actividad depredadora entre cepas de Arthrobotrys oligospora. Las cepas de A. oligospora presentaron diferencias altamente significativas en la actividad depredadora con larvas de NG provenientes tanto de ovejas como de cabras, (p=0.0001). La prueba de Tukey arrojó la presencia de 3 grupos tanto con NG provenientes de ovejas como de cabras (Tablas 6 y 7).
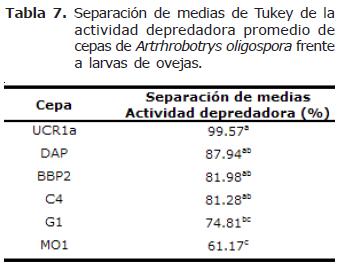
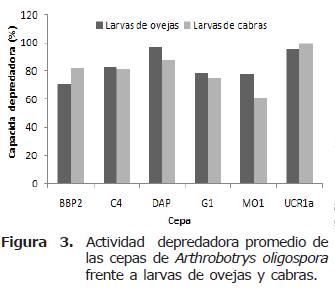
La actividad depredadora registrada por las cepas de A. oligospora presentó rangos más estrechos y fue más homogénea en comparación con las cepas de C. musiformis. Con larvas de NG provenientes de ovejas el rango fue de 70.6% a 96.8%, siendo la cepa BBP2 la menos eficiente y la cepa DAP la más eficiente. Con larvas de NG provenientes de cabras el rango fue de 61.2% a 99.6% siendo las cepas MO1 y UCR1a la menos efectiva y más efectiva respectivamente. Como se mencionó anteriormente, la mayoría de las cepas de esta especie fueron más eficientes en la depredación de larvas provenientes de ovejas; sin embargo, la diferencia en la depredación con larvas provenientes de cabras es muy estrecha en todos los casos (Figura 3).
Actividad depredadora de Dactylella sp. Dactylella sp. presentó una actividad depredadora promedio de 59.8% con larvas de NG provenientes de ovejas y 67.2% con larvas de NG provenientes de cabras. Como sólo se aisló una cepa de este género, (cepa FDJ1) no fue posible realizar alguna comparación o análisis estadístico.
Los testigos conteniendo únicamente larvas presentaron un bajo índice de mortalidad; 0.8% en larvas de NG provenientes de ovejas y 6.2% en larvas de NG provenientes de cabras. Los testigos conteniendo los hongos presentaron el crecimiento característico en AA, es decir un crecimiento muy pobre presentándose solo micelio vegetativo
DISCUSIÓN
Origen de las cepas. Teóricamente, los hongos aislados de muestras de estiércol, abonos orgánicos elaborados a partir de estiércol o de suelo que hayan estado en contacto con dicho material, ya sea directamente o abonado con fertilizantes cuya base sea estiércol, deberían presentar una mayor probabilidad de desempeñarse más eficientemente debido al contacto previo entre éstos y los nemátodos gastrointestinales (lo que se traduce en un mayor reconocimiento presa-depredador y a mayor reconocimiento mayor depredación). Este supuesto puede ser avalado por los resultados obtenidos ya que tanto en el ensayo con larvas de NG provenientes de ovejas como en el ensayo con larvas de NG provenientes de cabras, los hongos que presentaron la actividad depredadora más alta fueron precisamente los que se aislaron de muestras que estuvieron en contacto con estiércol o aislados directamente del mismo. Por ejemplo, la cepa de C. musiformis que presentó la mayor actividad depredadora con larvas de NG provenientes de ovejas (cepa PFC con 70.3%) fue aislada de suelo de potrero de pastoreo de cabras productoras de leche (Coronado, San José), donde las cabras depositan constantemente heces, existiendo un contacto directo entre este material y la biota del suelo.
El mismo caso se da para la cepa DAP de A. oligospora, cepa que registró la mayor actividad depredadora con larvas de NG provenientes de ovejas (96.8%), y que fue aislada de suelo de potrero de pastoreo de bovinos productores de leche (Moravia, San José). La misma situación se presenta en el ensayo con larvas provenientes de cabras; las cepas que presentaron mayor actividad depredadora fueron G3 (C. musiformis) con un 95.6%, aislada de estiércol de bovino de un sistema de doble propósito (Guápiles, Limón) y la cepa UCR1a (A. oligospora) con un 99.6% que también fue aislada de estiércol de bovinos (Sede del Atlántico de la Universidad de Costa Rica en Turrialba, Cartago).
En general las cepas que presentaron un mejor desempeño fueron: G3, UCR1b, UCR1a, C4 y G1 aisladas directamente de estiércol; PFSL1, PFC y DAP, aisladas de suelo de potreros y BBP2, aislada de abono orgánico elaborado a partir de estiércol de bovino y suelo de potrero.
Por otra parte, para el ensayo también se seleccionaron algunas cepas las cuales es muy probable nunca hayan estado en contacto con estiércol y mucho menos con larvas de nemátodos gastrointestinales, por ejemplo la cepa LS de C. musiformis aislada a partir de suelo de bosque secundario de la Estación Biológica "La Selva" (Sarapiquí, Heredia), la cual presentó una actividad depredadora baja (17.4 % con larvas de NG de ovejas y 21.0% con larvas de NG de cabras). Dicha ineficiencia puede deberse a la falta de reconocimiento entre el hongo y nemátodos de este tipo; sin embargo, no es posible hacer generalizaciones ya que cepas que probablemente tampoco han estado en contacto con nemátodos gastrointestinales como FDJ1 (Dactylella sp.), aislada de una finca ecológica (Talamanca) y FDH1 (C. musiformis), aislada de un sistema agroforestal de cacao (Talamanca) presentaron actividades de depredación intermedias (59.8% y 67.2% respectivamente).
Un último aspecto que es importante mencionar es que se cuenta con al menos una cepa con actividad depredadora elevada (más de 80%) por zonas específicas en cada una de las cinco provincias de estudio (Alajuela, Cartago, Heredia, Limón y San José). Uno de los aspectos a considerar en el control biológico en general, es el desempeño de antagonistas ajenos a la zona a la que se están aplicando, los cuales no siempre resultan efectivos en su función debido al estrés producido por las condiciones ambientales del lugar, a las que no están adaptados. La situación anterior podría sortearse si se aislan antagonistas autóctonos específicos de cada zona. En este caso se puede establecer que en cada una de las cinco provincias se cuenta con al menos un antagonista autóctono para ser utilizado como controlador biológico de nemátodos gastrointestinales; sin embargo, las condiciones ambientales dentro de una provincia varían, por lo que es recomendable probar en un futuro el o los hongos correspondientes en diferentes puntos de la provincia y observar si existe una eficiencia similar en la captura de larvas de NG.
Cepas. Las cepas de C. musiformis presentaron un rango bastante amplio de actividad depredadora; algunas fueron muy buenas capturando larvas, otras tuvieron un mediano desempeño y otras fueron deficientes. Por otra parte, las cepas de dicha especie parecieran capturar preferentemente larvas de NG provenientes de cabras, cuya población consistió de Haemonchus contortus en su totalidad, lo que indica que algunas cepas de C. musiformis presentan gran afinidad por dicho nemátodo, lo cual resulta beneficioso ya que este parásito es uno de los más problemáticos en pequeños rumiantes.
A pesar de que C. musiformis ha sido utilizado en ensayos de actividad depredadora contra nemátodos gastrointestinales, no existen datos concretos de su desempeño en la literatura consultada, por lo que no es posible realizar algún tipo de comparación. Por el contrario, existen algunos ensayos en los cuales se ha probado este hongo contra nemátodos fitoparásitos como Aphelenchus avenae, Neotylenchus linfordi, Meloidogyne sp., Radopholus citrophilus, Helycotylenchus sp., Pratylenchus sp., entre otros, obteniendo actividades depredadoras intermedias (13-15). Las cepas de C. musiformis utilizadas en este ensayo también desarrollaron una actividad depredadora intermedia, pero menor a la desarrollada por A. oligospora.
Las cepas de A. oligospora presentaron un rango estrecho de actividad de captura, donde la mayoría resultaron ser buenas depredadoras de larvas de NG (alrededor del 80% de actividad depredadora). Las cepas de esta especie presentaron una ligera preferencia por las larvas de NG provenientes de ovejas sobre las de cabras. Debido a que la totalidad de la población de larvas de NG provenientes de cabras era de Haemonchus contortus y que un 50% de las larvas de NG provenientes de ovejas correspondían a dicha especie, puede decirse que este hongo también es eficiente capturando este nemátodo.
Es importante señalar que las cepas costarricenses de A. oligospora presentaron actividades depredadoras más altas a las establecidas en la literatura, por ejemplo Mendoza de Gives et al (16) establecen una actividad depredadora de 35.9%; Wolstrup et al (17) indican un intervalo de 50.0% a 86.0%; Flores et al (18) un 50.1% y Flores et al (19) indica un rango de 40.2% a 64.0% para este hongo. A. oligospora es la especie aislada con más frecuencia en todo el mundo, por lo tanto en cada lugar el hongo estaría adaptado a las condiciones ambientales particulares de la zona, lo cual ejercería una influencia directa sobre su desempeño atrapando nemátodos, por lo que es de esperarse que cepas aisladas en diferentes lugares a pesar de ser la misma especie presenten actividades de captura diferentes frente a nemátodos gastrointestinales.
No se cuenta con información sobre la actividad depredadora de Dactylella sp., frente a larvas de NG; sin embargo, la actividad de depredación desarrollada por este hongo se puede considerar intermedia, similar a la indicada para A. oligospora en otras partes del mundo, por lo que puede establecerse que este hongo presenta una actividad depredadora aceptable.
Tipo de larvas. La mayoría de las cepas probadas de C. musiformis, A. oligospora y Dactylella sp., presentaron actividades de depredación mayores en presencia de la población de larvas de Haemonchus contortus provenientes de cabras en comparación con la población de larvas de NG provenientes de ovejas donde las actividades de depredación de los hongos fue menor en la mayoría de los casos.
Lo anterior indica una alta afinidad de los hongos por Haemonchus contortus. La población larvaria proveniente de ovejas consistió en un 50% de este nemátodo, por lo que pudiera establecerse que una buena parte de las larvas atrapadas en este caso pertenecían a dicha especie. Desgraciadamente la metodología empleada no permite realizar esta diferenciación; es decir, no es posible reconocer qué tipo de larvas se encuentran atrapadas en las trampas cuando se realizan las observaciones.
Existen diferencias en la eficiencia de depredación con la cual los hongos nematófagos capturan diferentes especies de nemátodos, pero en general estos hongos presentan un rango amplio de hospederos (6, 14). La interacción entre nemátodos y hongos nematófagos es un proceso complejo que envuelve polímeros superficiales, actividades enzimáticas y reconocimiento lectinas-carbohidratos entre las trampas pegajosas y los nemátodos. Las diferencias en la actividad depredadora de los hongos nematófagos están relacionadas directamente con la composición de la cutícula, que varía entre las especies de nemátodos (3, 16, 20). Otros estudios establecen que específicamente es la variación de los patrones de proteína de la cubierta antigénica la responsable del reconocimiento hongo-nemátodo (16, 19). Lo anterior determinaría la especificidad de los hongos nematófagos por sus hospederos (20).
Los resultados obtenidos muestran que existe una gran variabilidad en la actividad depredadora de las cepas de los diferentes hongos probados y que existen tendencias y no reglas que puedan generalizarse, ya que la actividad depredadora está influenciada por la especie y procedencia de los hongos nematófagos y especie de larvas a la que se los enfrenta.
En conclusión, el análisis estadístico indicó diferencias altamente significativas en la actividad de los hongos; sin embargo, las cepas de A. oligospora mostraron un mejor desempeño en la captura de nemátodos comparado con las cepas de C. musiformis y Dactylella sp. La heterogeneidad encontrada responde a factores como: origen de las cepas, características intrínsecas de las mismas y especies de larvas utilizadas.
Referencias [ Links ]
2. Waller JP. Anthelmintic resistance. Vet Parasitol 1997; 72:391–412. [ Links ]
3. González CME, Mendoza de Gives P and Quiróz RH. Comparison of the trapping ability of Arthrobotrys robusta and Monacrosporium gephyropagum on infective larvae of Strongyloides papillosus. J Helminthol 1998; 72:209–213. [ Links ]
4. García RC. Ecoepidemiología y control de parasitosis. Ganadería ecológica [en línea] 2001 [acceso: 7 octubre 2003]. URL disponible en: http://www.ganadería-ecologica.com/documentos/articulo_final.phtml?codigo=41 [ Links ]
5. Cole GT and Kendrick B. (editors). Biology of Conidial Fungi Vol. 2. EUA: Academic Press; 1981. [ Links ]
6. Barrón GL. The nematode destroying fungi. Lancaster, Pennsylvania: Editorial Lancaster Press, Inc; 1977. [ Links ]
7. Larsen M. Biological control of helminths. Int J Parasitol 1999; 29:139– 146. [ Links ]
8. Araújo JV; Assis RCL; Campos AK; Mota MA. Antagonistic effect of nematophagus fungi Monacrosporium, Arthrobotrys and Duddingtonia on infective Cooperia sp. and Oesophagostomum sp. larvae. Arq Bras Med Vet Zootec 2006; 58(3):373–380. [ Links ]
9. Paraud C; Pors I; Chicard C; Chartier C. Comparative efficacy of the nematode-trapping fungus Duddingtonia flagrans against Haemonchus contortus, Teladorsagia circumcincta and Trichostrongylus colubriformis in goat faeces: influence of the duration and the temperature of coproculture. Parasitol Res 2006; 98:207-213. [ Links ]
10. Campos AK; Araújo JV; Guimarães MP; Dias AS. Resistance of different fungal structures of Duddingtonia flagrans to the digestive process and predatory ability on larvae of Haemonchus contortus and Strongyloides papillosus in goat faeces. Parasitol Res 2009; 105: 913 – 919. [ Links ]
11. Mendoza de Gives P, Flores CJ, Herrera RD, Vázquez PV, Liébano HE, Ontiveros FGE. Biological control of Haemonchus contortus infective larvae in ovine faeces by administering an oral suspensión of Duddingtonia flagrans chlamydospores to sheep. Rev Latinoam Microbiol 1998; 72:343–347. [ Links ]
12. Hernández GJ. (Compilador). Técnicas Coproparasitológicas. Escuela de Medicina Veterinaria. Universidad Nacional de Costa Rica: Heredia; 2001. [ Links ]
13. Monoson LR. Trapping effectiveness of five species of nematophagus fungi cultured with mycophagus nematodes. Mycologia 1968; 60:788-801. [ Links ]
14. Stirling RG. Biological control of plant parasitic nematodes: progress, problems and prospects. United Kingdom, CAB International, 1991. [ Links ]
15. Durán MJ. Guía de ingredientes activos de bioplaguicidas. CATIE [en línea] 2004 [acceso: 12 septiembre 2005]. URL disponible en: http://www.bioplaguicidas.org/Bioplaguicidas04/Documentos/libros%20BPNOQ/guia%20ingredientes%20enero%202004.pdf [ Links ]
16. Mendoza de Gives P, Zavaleta ME, Herrera RD, Quiróz RH. In vitro capability of Arthrobotrys spp. on infective larvae of Haemonchus contortus and Nacobbus aberrans. J Helminthol 1994; 68:223-229. [ Links ]
17. Wolstrup J, Nansen P, Grønvold J, Henriksen SA, Larsen M. Toward practical biological control of parasitic nematodes in domestic animals. J Nematol 1996; 28(2):129-132. [ Links ]
18. Flores CJ, Herrera RD, Vázquez PV, Flores CR, Líebano HE, Mendoza de Gives P. Acción depredadora in vitro de ocho aislados de hongos contra el nemátodo Panagrellus redivivus. Rev Latinoam Microbiol 1999; 41:239-244. [ Links ]
19. Flores CJ, Herrera RD, Mendoza de Gives P, Líebano HE, Vázquez PVM, López AME. The predatory capability of three nematophagus fungi in the control of Haemonchus contortus infective larvae in ovine faeces. J Helminthol 2003; 77:1-8. [ Links ]
20. Dijksterhuis J, Veenhuis M, Harder W, Nordbring-Hertz B. Nematophagus fungi: structure/function relationships and physiological aspects. Adv Microb Physiol 1994; 36: 112-143. [ Links ]

