










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista MVZ Córdoba
Print version ISSN 0122-0268
Rev.MVZ Cordoba vol.18 no.3 Córdoba Sept./Dec. 2013
ORIGINAL
Efecto de la suplementación con grasas protegidas sobre parámetros productivos y reproductivos en vacas lactantes
Effect of protected fat supplementation on productive and reproductive performance in lactating cow
Mónica Duque Q,1,2* M.Sc, Ricardo Rosero N,2 Ph.D. Jorge Gallo,1,3 Esp. Martha Olivera A,1 Dr. Sci. Agr.
1Universidad de Antioquia, Facultad de Ciencias Agrarias, Grupo Biogénesis. Medellín, Colombia.
2Universidad de Antioquia, Facultad de Ciencias Agrarias, Grupo GRICA. Medellín, Colombia.
3Nutraceutika S.A. Medellín, Colombia.
*Correspondencia: mduque82@yahoo.com
Recibido: Abril de 2012; Aceptado: Marzo de 2013.
RESUMEN
Objetivo. Investigar el efecto de la suplementación con jabones cálcicos protegidos, conteniendo ácidos grasos omega 3 ó 6 sobre la producción, composición de la leche y desempeño reproductivo. Materiales y métodos. Se utilizaron 24 vacas en un diseño mixto desde 20 días antes de la fecha prevista del parto hasta 105 días postparto, asignadas a los diferentes tratamientos; C: dieta control, O6: animales suplementados con grasas protegidas conteniendo omega 6 y O3: animales suplementados con grasas protegidas conteniendo omega 3. Se determinó la producción y la composición de la leche, así como la condición corporal (CC), el primer calor y primera ovulación posparto. Resultados. La suplementación con omega 3 ó 6 no afectó la producción láctea, ni los porcentajes de grasa, proteína, sólidos no grasos (SNG) y sólidos totales (ST). Sin embargo, hubo un efecto positivo a los 45 días posparto sobre la CC cuando los animales fueron suplementados con omega 6 (p<0.05) lo que se reflejó en una recuperación ovárica posparto más rápida (42.24±16.75 días a la primera ovulación posparto). Conclusiones. La utilización de la suplementación con omega 6 mejoró la condición corporal de los animales en el posparto temprano, disminuyendo los días a la primera ovulación y primer calor posparto, sin afectar la producción ni composición de la leche. La suplementación con omega 3 no influyó en ningún parámetro productivo en el posparto temprano.
Palabras clave: Omega 3, omega 6, suplementos alimentarios, tratamiento con dieta (Fuente: CAB).
ABSTRACT
Objective. To investigate the effects of supplements with protected fat such as calcium soaps containing omega 3 or 6 milk production, composition and reproductive performance. Materials and methods. Twenty-four Holstein cows were used in a mixed model design. From 20 days before calving until 105 days postpartum, cows were assigned to different treatments; C: control diet, O6: animals supplemented with protected unsaturated fat including a higher amount of omega, 6 and O3: animals fed with protected unsaturated fat including higher amounts of omega 3. Milk volume and composition, and body condition score (CC) were determined, along with the first postpartum heat and ovulation. Results. Supplementation with omega 3 or 6 had no effect on milk volume, and fat percentages, protein, milk non-fat solids (SNF) and total solids (ST). However, there is a positive effect of omega 6 supplementation on the CC at 45 days postpartum (p<0.05). This was also reflected in a faster ovarian recovery (42.24±16.75 days to first postpartum ovulation). Conclusions. The use of omega 6 supplementation improves body condition score of animals during early postpartum, decreasing the days to the first ovulation and postpartum heat without affecting the volume or composition of the milk. Supplementation with omega 3 does not affect any productive parameters in the early postpartum.
Key words: Diet prescription, food supplements omega 3, omega 6 (Source: CAB).
INTRODUCCIÓN
Durante el primer tercio de lactancia, las vacas lecheras sufren un déficit energético importante (1), debido al bajo consumo de materia seca y los altos requerimientos para satisfacer no sólo las necesidades de mantenimiento sino para producción, lo que trae como consecuencia un incremento en la movilización de reservas corporales y, en muchos casos, una reducción en el desempeño productivo y reproductivo de los animales. Con el fin de incrementar el consumo de energía de las vacas en producción, la suplementación con grasas protegidas parecen ser una buena alternativa, al ser grasas inertes a nivel ruminal (2, 3) que no alteran la digestión de la fibra, la concentración de los ácidos grasos volátiles y la eficiencia de síntesis microbiana, pero que pueden cambiar la composición de la grasa de la leche o de los distintos depósitos adiposos a nivel de la canal del animal, que en mayor o menor medida logran pasar a la leche o se depositan a nivel corporal (4). Además, ácidos grasos esenciales como el linoleico (omega 6) y linolénico (omega 3) tienen efectos directos sobre los procesos fisiológicos; alterando positivamente la reproducción alterando las funciones del folículo ovárico y el cuerpo lúteo vía mejoramiento del estatus energético del animal y por el incremento de precursores para la síntesis de hormonas reproductivas como los esteroides y prostaglandinas. Por ésta razón su importancia para la salud uterina, acompañada finalmente por ovulaciones más tempranas (5, 6).
Se espera por lo tanto, que la suplementación con ácidos grasos omega 3 u omega 6 protegidos, disminuya la movilización de reservas corporales y de esta forma se mejore la condición corporal de los animales, se disminuya el balance energético negativo en la lactancia temprana y finalmente, se mejore la composición de la leche y se alcance un restablecimiento ovárico posparto mucho más temprano. Adicionalmente, se quiere determinar si la suplementación de cada ácido graso sigue una vía diferente o sus respuestas reproductivas son similares.
El objetivo de éste trabajo fue evaluar el efecto de las grasas protegidas conteniendo omega 3 ó 6 sobre la producción y composición de la leche, así como la condición corporal, primer calor y primera ovulación posparto de vacas Holstein en pastoreo, en el primer tercio de lactancia.
MATERIALES Y MÉTODOS
Sitio de estudio. La investigación fue realizada en la hacienda Yerbabuena, localizada en el municipio de Entrerríos (Antioquia), a una altura de 2300 msnm, temperatura promedio de 16°C, humedad relativa del 72% y clasificada como una zona de vida bosque húmedo montano bajo (bh-MB).
Animales. 24 Vacas multíparas Holstein preñadas, entre 3 y 5 lactancias fueron aleatoriamente asignadas 20+9 días antes de la fecha prevista de parto a uno de los 3 tratamientos hasta los 105 días posparto: Control: Kikuyo y alimento concentrado, Omega 3: kikuyo, alimento concentrado y suplementación con grasa protegida conteniendo omega 3 y Omega 6: kikuyo, concentrado y suplementación con grasa protegida conteniendo omega 6.
Manejo de la nutrición. Los animales fueron mantenidos bajo un sistema intensivo, pastoreando kikuyo (Pennisetum clandestinum) en un sistema rotacional por franjas. Los tratamientos se diferenciaron por el tipo de concentrado ofrecido a los animales: Control: pasto Kikuyo y alimento concentrado sin adición de grasa insaturada protegida, tratamiento Omega3: pasto Kikuyo más alimento concentrado y enriquecido con grasa protegida insaturada conteniendo ácido graso omega 3 y tratamiento Omega6: pasto Kikuyo más alimento concentrado enriquecido con grasa protegida conteniendo ácidos grasos omega 6. A todos los animales se les adaptó gradualmente al consumo de suplemento concentrado con grasa protegida 20 días antes del parto, incrementando 100 g/día paulatinamente hasta llegar a la cantidad total que se suplementaría.
Los concentrados experimentales (8 Kg/vaca/día) fueron ofrecidos en dos cantidades iguales durante el ordeño mecánico de la mañana y de la tarde a razón de 4 Kg/vaca/día y de 280 g de grasa protegida para asegurar un consumo total de 560 g/vaca/día de la grasa protegida sin superar el 6% de la materia seca (MS) del concentrado. En los grupos omega3 y omega6 la grasa protegida y la torta de soya fueron adicionadas al concentrado comercial con el fin de incrementar la densidad energética y de no afectar el nivel de proteína de la dieta total por efecto de dilución.
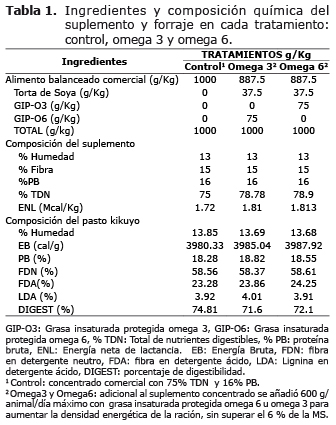
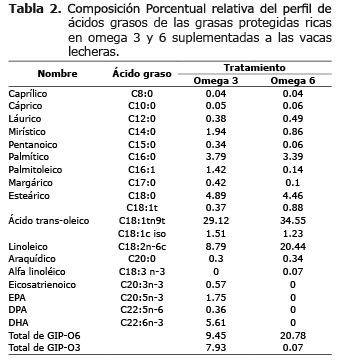
Los tratamientos control, Omega3 y Omega6 fueron isoprotéicos (16.6, 17.1, 17.4% PB respectivamente) y los tratamientos omega 3 y omega 6 isoenergéticos (78.78 y 78.9 % TDN), (Tabla 1). El perfil de ácidos grasos de las grasas ricas en omega 6 y 3 se presenta en la tabla 2 y su determinación se realizó según la metodología de Rico y colaboradores (7).
Recolección de muestras. Muestras de pasto kikuyo fueron cortadas del potrero a los 30 y 60 días después del parto de cada uno de los animales. La composición final de la pastura fue estimada a partir de la obtención de un pool de muestras que fueron colectadas varias veces en estos períodos.
Para determinar el porcentaje de digestibilidad de la materia seca (DIGEST) fue utilizado como marcador interno, la lignina en detergente ácido indigestible (LDA) (8), para éste propósito se recolectaron muestras de heces durante 3 días de cada unidad experimental a los 30 y 60 días posparto, obteniéndose un pool de muestras para cada animal en cada período experimental.
La producción y composición de la leche fue registrada en el ordeño de la mañana y la tarde, cada 15 días después del parto hasta los 105 días posparto. La condición corporal fue registrada por el mismo observador durante todo el período experimental, usando una escala de 1 a 5 puntos (1=muy flaco, 2=delgada, 3=promedio, 4=engrasada, 5=obeso), desde 20 días antes del parto, al parto y cada 15 días durante 105 días (9).
Para la determinación de la primera ovulación posparto por concentración de progesterona, fueron tomadas muestras de sangre de la vena coccígea 2 veces por semana después del día 10 posparto hasta 2 semanas después que el animal presentara el primer calor posparto detectado en la finca. El primer calor posparto fue evaluado por ayuda de los detectores de monta Kamar™, el cual fue colocado sobre el sacro de la vaca.
Análisis de muestras y datos. Las muestras de forraje y de los suplementos con omega 3 y 6 fueron secadas a 60°C en una estufa de ventilación forzada, molidas posteriormente a 1 mm y analizadas para determinar sus concentraciones de MS, energía bruta (EB), Proteína Bruta (PB). Se determinó finalmente para las muestras de kikuyo, fibra en detergente neutro (FDN), fibra en detergente ácida (FDA) y lignina en detergente ácido indigestible (LDA), para determinar la digestibilidad del forraje (8). Las muestras de heces fueron secadas a 60°C en una estufa de ventilación forzada, molidas posteriormente a 1 mm y analizadas para determinar sus concentraciones de MS y (LDA).
Los requerimientos energía neta de lactancia (ENL) y el consumo total de MS (CMSt) para cada uno de los animales fueron estimados con base en las recomendaciones del NRC (2001). El CMS del forraje (CMSf) se estimó como la diferencia entre CMSt y el consumo MS del suplemento (CMSs). Con base en estas determinaciones fueron calculados los aportes de ENL (Mcal/Kg), PB (g/día) al momento del parto, 30 y 60 días posparto; el balance energético (BE) y proteico (BP) de los animales.
La composición de la leche fue analizada por espectrofotometría infrarroja (EKOMILK) determinándose su concentración de grasa, proteína, sólidos no grasos (SNG) y sólidos totales (ST).
Para determinar la leche corregida al 4% de grasa se utilizó la siguiente fórmula (NRC, 1989):
FCL = 0.4 x kg Leche + 15 x kg Grasa
Para determinar las concentraciones de progesterona las muestras fueron procesadas mediante inmunoensayo de electroquimioluminiscencia utilizando para ello el kit de diagnóstico de Progesterone II (Roche Elecsys Cobas número de catálogo: 12145383), (laboratorio Centrolab, Medellín, Colombia). El coeficiente de variación intraensayo fue de 1.38% y la cantidad mínima detectable fue 0.030 ng/ml. El primer aumento en la progesterona postparto, definido como mayor a 1.5 ng/ml indicó el inicio de la actividad luteal, pero siempre y cuando este aumento estuviera seguido de las siguientes dos muestras con valores mayores a 1.5 ng/ml también. La detección del calor se determinó cuando el detector KAMAR cambiara a un color rojo. Conjuntamente con el cambio del color a rojo del detector KAMAR, otros signos de estro en los animales fueron observados (monta a otros animales, vulva enrojecida y presencia moco cérvico-vaginal).
Análisis estadístico. La producción, composición de leche y la condición corporal fueron evaluadas por medidas repetidas en el tiempo, y el consumo de MS, características del forraje y balance energético por el procedimiento PROC GLM (SAS 9.1.3 Versión, SAS Institute Inc.). Como covariables se emplearon la condición corporal previa al experimento, el promedio de leche producida obtenido en los primeros 100 días de la lactancia previa, días de secado y edad, para reducir la variación y los posibles efectos no relacionados al tratamiento. Para establecer el modelo más idóneo se utilizaron los criterios Akaike (AIC) y BIC Bayesiano (BIC).
El experimento fue de tipo desbalanceado analizando las fuentes de variabilidad asociadas con el diseño de estructura experimental tipo III. El análisis se complementó mediante la técnica descriptiva unidimensional por tratamiento y período con el objetivo de establecer los estadísticos (media aritmética, desviación típica y coeficiente de variación) y mediante la técnica ANOVA con contraste canónico de índole ortogonal estableciendo por el método de la máxima verosimilitud la dimensionalidad del contraste para las variables de respuesta que estaban asociadas a la producción, composición de leche, condición corporal y variables reproductivas.
El modelo para el análisis de medidas repetidas en el tiempo se presenta a continuación:
Yijk = µ + αi + dij + Tk + (αr)jk + εijk
Donde:
Yijk: Es la respuesta en el tiempo k en el animal j en el grupo de tratamiento i
µ: Es la media poblacional
αi: Es el efecto fijo del tratamiento i
dij: Es el efecto aleatorio del animal j en el tratamiento i
Tk: Es el efecto fijo en el tiempo k
(α r)ik: Es la interacción del efecto fijo del tratamiento i con el tiempo k
εijk: Es el error aleatorio en el tiempo k en el animal j en el tratamiento i.
RESULTADOS
Consumo de MS, características del forraje, balance energético y proteico. Las muestras de pasto kikuyo tomadas a los 30 y 60 días posparto no variaron en su composición en cuanto a sus concentraciones de EB, PB, FDN, FDA, LDA y digestibilidad de la MS (Tabla 1), siendo por lo tanto el aporte del alimento el mismo en los dos períodos.
El consumo de MS total difiere a los 30 y 60 días posparto en un mismo tratamiento (Tabla 3). Sin embargo, no se presentaron diferencias significativas cuando se evaluó en un mismo período los diferentes tratamientos. A los 60 días posparto, en todos los tratamientos, los animales presentaron un mayor consumo de MS total por el mayor incremento en el consumo de MS del forraje ya que no se encontró cambios en el CMS del suplemento que permaneció sin diferencias significativas a los 30 y 60 días posparto (Tabla 3).

Se encontró diferencias significativas a los 30 y 60 días en un mismo tratamiento para el BE y BP de los animales. No se encontró diferencias significativas para un mismo período en los diferentes tratamientos (p>0.05). La producción de leche no tuvo diferencias significativas entre tratamientos para un mismo período, ni entre períodos en un mismo tratamiento.
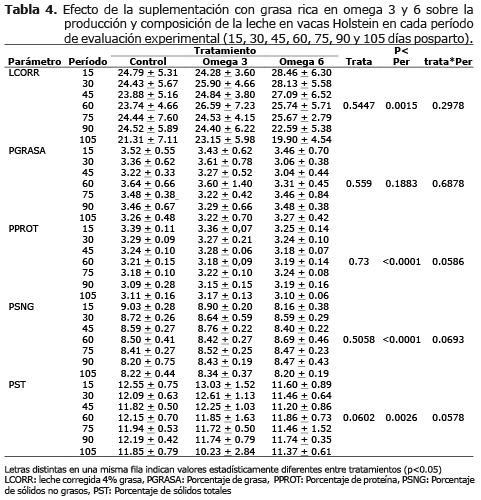
Producción de leche y composición. En todo el período experimental (105 días de evaluación), las producciones medias de leche corregidas al 4% de grasa fueron estadísticamente iguales en todos los tratamientos. La suplementación con grasa protegida omega 3 y omega 6 no tuvo efecto en la composición de la leche en cuanto a porcentaje de grasa, proteína, SNG, y ST (p>0.05), (Tabla 4).
Condición corporal. El comportamiento de la condición corporal de los animales suplementados mostró diferencias significativas (p<0.05) a partir del día 45 posparto en animales suplementados con omega 6 y a partir del día 90 en animales suplementados con omega 3, respecto al tratamiento control (Tabla 5).
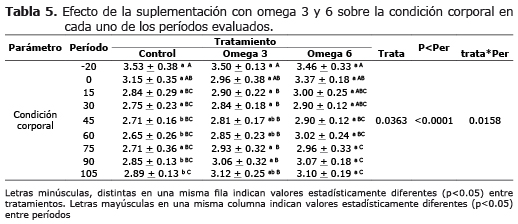
Los animales suplementados con grasas protegidas perdieron menos condición corporal a lo largo del tiempo que los animales del tratamiento control, aunque las diferencias estadísticas sólo se observan después del día 45 posparto. Los animales del tratamiento omega 6 al perder menos condición corporal (CC) en los diferentes períodos, la recuperaron fue más rápida después del pico de producción.
Primera ovulación y primer calor posparto. La condición corporal fue equivalente entre los tratamientos omega 3 y control entre los 15 y 75 días posparto. Discrepancias estadísticas entre estos tratamientos solo fueron verificadas a partir del día 90 posparto (p<0.05) (Tabla 6).
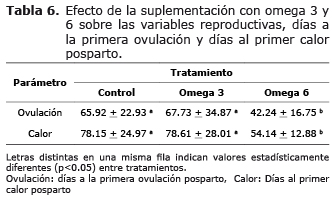
La suplementación con grasa omega 6, permitió mejorar la condición corporal de los animales, disminuir los días a la primera ovulación y el primer calor posparto cuando fue comparado con los tratamientos control y omega 3 (p<0.05). La suplementación con omega 3, no tuvo efecto sobre estas variables al igual que el tratamiento control.
DISCUSIÓN
Características de la dieta y consumo de alimento. Los animales en cada uno de los tratamientos presentaron un balance energético negativo (BEN) a los 30 días posparto. Esta condición no se presentó a los 60 días posparto. Este mejor BE a los 60 días posparto se atribuyó a un aumento en el CMS proveniente del forraje. El consumo de materia seca fue estimado de acuerdo con las recomendaciones del NRC de 2001, el cual basa sus predicciones llevando en consideración la composición de la leche y el peso vivo de los animales. En este experimento no fueron encontradas diferencias significativas para esta variable entre los tratamientos, en todos los períodos analizados. Sin embargo, cuando se comparó el CMS al interior de cada tratamiento se verifica que el consumo aumentó al día 60 posparto en todos los tratamientos. Esto ha sido reportado por diversos autores, donde el CMS se mejora después del día 45 posparto (10,11).
Salado et al (12) al suplementar vacas Holstein con grasa, encontraron que el CMSf no fue afectado acorde con nuestro estudio y tampoco evidenciaron efectos negativos de la suplementación en la función ruminal. Aunque, se ha reportado que en vacas lactantes, la grasa protegida puede reducir el consumo de MS (13, 14), aun cuando la grasa se añade directamente en el duodeno. Parece ser que la razón para esta disminución en el consumo podría estar asociada a un aumento en la concentración de los ácidos grasos libres (AGL) circulantes que proceden de la captación tisular incompleta de la hidrólisis de los triglicéridos de quilomicrones y VLDL por la lipoproteína lipasa (15). Sin embargo, la relación entre el consumo de lípidos protegidos y la disminución del CMS por un alto nivel de AGL es muy contradictoria en la literatura, debido a que algunos trabajos presentan niveles elevados mientras otros presentan niveles bajos, y aun así el consumo disminuye (15).
Los animales suplementados con omega 6 y omega 3 presentaron un BEN sin diferencias estadísticamente significativas con respecto a los animales control en el día 30 posparto y al día 60 posparto entre los tratamientos. Esta situación puede atribuirse a que aunque se incrementa la energía total de la dieta, los animales no alcanzan a llenar sus requerimientos energéticos en los tratamientos omega 6 y 3 principalmente debido a la mayor producción de leche 3.70 y 1.47 litros más que el tratamiento control respectivamente, aunque estadísticamente no hubieran diferencias significativas. Se observa también que a los 60 días posparto, el BE es positivo para todos los tratamientos y que el balance de proteína (g/día) excede los requerimientos (Tabla 3); esto puede incrementar las necesidades energéticas de los animales para eliminar excesos de proteína cruda para la formación de urea en el hígado (16) y por lo tanto, uno de los destinos de la energía añadida a la dieta sea hacia esta vía.
Condición corporal, producción y composición de la leche. Condición corporal, producción y composición de la leche. El hecho de no haber encontrado diferencias significativas entre los tratamientos en cuanto al porcentaje de grasa y proteína láctea está señalando que las dietas formuladas aportaron al rumen una cantidad suficiente de nutrientes para una actividad fermentativa ruminal normal y que la inclusión de grasas protegidas omega 3 y 6 hasta niveles del 6% no reflejó problemas ruminales que pudieran haber alterado la síntesis láctea. Además, la falta de efectos de los tratamientos con omega 3 y 6 sobre el porcentaje de grasa, proteína, SNG y ST entre los tratamientos, pudo deberse a que la suplementación con grasa fue utilizada para producir energía por los tejidos; disminuyendo la movilización de grasa en el animal, lo que se vio reflejado en una mejor condición corporal después del día 45 posparto en animales suplementados con omega 6 y después del día 90 posparto en animales suplementados con omega 3.
Éstos resultados estuvieron acordes con estudios donde no se encontraron efectos significativos en producción de leche, porcentaje de grasa y proteína cuando se suplementó los animales con jabones cálcicos de ácidos grasos omega 3 (17, 18). Contrario a lo encontrado en este trabajo, algunos estudios muestran que hay un efecto de las grasas insaturadas en la producción de la grasa en la leche de animales en pastoreo, explicado por una disminución en la concentración de grasa láctea y el aumento de la producción de leche (19), atribuyéndose un efecto directo de intermediarios de ácidos grasos formados en el rumen durante la biohidrogenación que causan disminución en la síntesis de novo de los ácidos grasos en la glándula mamaria cuando las grasas no están bien protegidas o cuando su inclusión en la dieta excede el 8% (20-22). En general, algunos autores sustentan que la suplementación con grasa de la dieta parece aumentar la producción sin afectar la composición de la leche de vacas alimentadas con pastos de alta calidad. Estos aumentos en la producción de leche pueden estar relacionados con una mejor utilización de la energía en lugar de un aumento de la ingestión energética. Los autores sugieren que el ahorro en la utilización de la glucosa podría ser, al menos parcialmente, el causante de este resultado, ya que al administrar una dieta suplementada con grasa, los ácidos grasos resultan ser una fuente de energía para los tejidos, así como para la síntesis de triglicéridos en la mama (2, 22).
Respecto a la condición corporal (Tabla 5), la mayor pérdida se produjo durante las cuatro primeras semanas postparto, siendo el grupo control, quien presentó la mayor disminución durante este período. Sin embargo, esta pérdida continuó hasta el día 45 de lactancia para el grupo omega 6, a partir de la cual se observó una recuperación de la CC hasta el final del período experimental y sólo a partir del día 90 en los animales con omega 3 con respecto al control. Estos resultados coinciden con lo afirmado por especialistas en cuanto a que el manejo de los animales debe enfocarse hacia que la máxima pérdida de CC se produzca no más allá de la tercera semana postparto e iniciar su recuperación y de esta manera disminuir la duración del balance energético negativo, evitando trastornos metabólicos y reproductivos asociados (23).
En éste trabajo, la partición de nutrientes estuvo dirigida a la menor movilización de grasa corporal principalmente en el tratamiento con omega 6, ya que con la suplementación de grasa protegida al aumentar la densidad energética de la ración mejora la eficiencia de utilización de la energía (disminución en la pérdida de energía como calor y metano, uso directo de ácidos grasos secretados en la grasa de la leche y mejor eficiencia en la producción de ATP desde AG de cadena larga que desde el acetato) (24, 25). Por lo cual, el animal mantiene la misma producción y composición de la leche sin necesidad de perder tanta CC (Tabla 5) al utilizar la grasa protegida como fuente de energía para los tejidos y para la producción y composición láctea. Acorde con nuestros hallazgos, otros estudios encontraron que los animales que se suplementaron con grasa protegida presentan una menor pérdida de CC (22, 23). Otros autores reportan en sus trabajos la pérdida de condición corporal cuando utilizan grasas protegidas, para apoyar una mayor producción de leche (26).
Condición corporal, primera ovulación y primer calor posparto. El hecho de encontrar la condición corporal por debajo de 3.0 muestra que todos los animales de los diferentes tratamientos estaban en un balance energético negativo. La suplementación con grasa omega 6 mejora la condición corporal de los animales después del día 45 (2.90+0.12), y en el omega 3 en el día 90 posparto con respecto al tratamiento control (3.06+0.32), por lo tanto se ve una presentación de la primera ovulación posparto mucho más rápida (45 días) y el primer calor a los 54 días en el tratamiento con omega 6, con un cuerpo lúteo de corta duración en todos los tratamientos.
Diversos autores reportan que hay una relación inversa entre CC y PL (27) donde la mayor producción de leche en la lactancia temprana disminuye el grado de condición corporal de los animales al movilizar ácidos grasos desde tejido adiposo para la obtención de energía. Sin embargo, si la pérdida de condición corporal es menor, los animales podrían recuperarla más rápidamente y tener efectos benéficos en la reproducción al restablecerse el eje somatotrópico, como lo confirman los resultados encontrados en este estudio. Además, se sabe que el incremento en el intervalo para la primera ovulación en vacas se debe a pérdidas mayores en la condición corporal.
González y Bas (22) atribuyen que una mejor fertilidad se obtiene cuando el animal utiliza la energía de los alimentos en lugar de la energía de las reservas de grasa corporal, lo que da como resultado un adelanto en el reinicio de su ciclo estral y una reducción de los días abiertos. Además, una alta concentración de progesterona antes y después de la inseminación está relacionada con una mayor tasa de preñez. Esta alta concentración de progesterona está asociada con el contenido de lípidos en las células luteales, atribuible a la suplementación de grasas en la dieta, que mejora la función del cuerpo lúteo en los ovarios y un mayor desarrollo folicular (28).
Otro de los efectos por los cuales la alimentación con grasa protegida pudo mejorar la respuesta sobre la primera ovulación y el primer calor posparto es por el aumento en la producción de estrógenos. El colesterol estaría influyendo en la síntesis de estrógenos derivado de la absorción de lipoproteínas de colesterol en plasma principalmente en los animales suplementados con omega 6. Además, la ruta de síntesis de hormonas de éste ácido graso es la producción de prostaglandinas F2 alfa que permiten la disminución de los días a la primera ovulación, mientras que la ruta de los omega 3 puede por el contrario, inhibir la síntesis de prostaglandina F2 alfa a través de mecanismos tales como la disminución de la disponibilidad de su precursor el ácido araquidónico, un aumento de la competencia por estos ácidos grasos con el ácido araquidónico por la unión a prostaglandina H sintasa, y la inhibición de la síntesis de prostaglandina sintasa H y su actividad. Está inhibición finalmente, parece ser importante para la amplificación de la supresión de la síntesis de prostaglandina F2 alfa uterina durante la preñez temprana, lo que puede contribuir a una reducción en la mortalidad embrionaria, pero no para disminuir principalmente los días a la primera ovulación posparto (29,30).
En conclusión, la suplementación con grasas protegidas de jabones cálcicos conteniendo omega 6 y 3 no afecta significativamente la producción y composición de leche. La suplementación con grasa protegida mejora significativamente la CC de los animales en la lactancia temprana, disminuyendo los días al primer calor y primera ovulación posparto. La suplementación con omega 3 no influye en ningún parámetro productivo en el posparto temprano.
Agradecimientos
Fundación Universitaria San Martín, por la financiación del proyecto “Efecto de la suplementación con ácidos grasos omega 3 y 6 protegidos sobre parámetros productivos, reproductivos y balance energético en vacas Holstein en el trópico alto” y a Colciencias por la beca Jóvenes investigadores e innovadores para el año 2009.
REFERENCIAS
1. Bargo F, Muller L, Kolver E, Delahoy J. Invited review: production and digestion of supplemented dairy cows on pasture. J Dairy Sci 2003;86(1):1-42. [ Links ]
2. Schroeder G, Gagliostro GA, Bargo F, Delahoy J, Muller L. Effects of fat supplementation on milk production and composition by dairy cows on pasture: a review. Livest Prod Sci 2004;86(1-3):1-18. [ Links ]
3. Gagliostro G, Schroeder G. Efectos de la suplementación con sales cálcicas de ácidos grasos insaturados sobre la digestión ruminal en vacas lecheras en pastoreo. Arch Latinoamer Prod Anim 2007;15:88-99. [ Links ]
4. Jenkins TC, Bridges Jr WC. Protection of fatty acids against ruminal biohydrogenation in cattle. Eur J Lipid Sci Technol 2007;109(8):778-89. [ Links ]
5. Bilby T, Jenkins T, Staples C, Thatcher W. Pregnancy, bovine somatotropin, and dietary n-3 fatty acids in lactating dairy cows: III. Fatty acid distribution. J Dairy Sci 2006;89(9):3386-99. [ Links ]
6. Bilby T, Sozzi A, Lopez M, Silvestre F, Ealy A, Staples C, et al. Pregnancy, bovine somatotropin, and dietary n-3 fatty acids in lactating dairy cows: I. ovarian, conceptus, and growth hormone "insulin-like growth factor system responses. J Dairy Sci 2006;89(9):3360-74. [ Links ]
7. Rico JE, Moreno B, Pabón ML, Carulla JE. Composición de la grasa láctea en la sabana de Bogotá con énfasis en ácido ruménico-CLA cis-9, trans-11. Rev Colomb Cien Pecu 2009;20(1):30-9. [ Links ]
8. Goering H, Van Soest P. Forage fiber analyses (apparatus, reagents, procedures, and some applications). Agric Handbook 1970; 379:1-20. [ Links ]
9. Roche J, Macdonald K, Burke C, Lee J, Berry D. Associations among body condition score, body weight, and reproductive performance in seasonal-calving dairy cattle. J Dairy Sci 2007;90(1):376-91. [ Links ]
10. Grummer RR, Mashek DG, Hayirli A. Dry matter intake and energy balance in the transition period. Vet Clin North Am Food Anim Pract 2004;20(3). [ Links ]
11. Nielsen N, Ingvartsen K. Propylene glycol for dairy cows: A review of the metabolism of propylene glycol and its effects on physiological parameters, feed intake, milk production and risk of ketosis. Anim Feed Sci Technol 2004;115(3-4):191-213. [ Links ]
12. Salado E, Gagliostro G, Becu-Villalobos D, Lacau-Mengido I. Partial replacement of corn grain by hydrogenated oil in grazing dairy cows in early lactation. J Dairy Sci 2004; 87(5):1265-78. [ Links ]
13. Schroeder G, Gagliostro G, Bargo F, Delahoy J, Muller L. Effects of fat supplementation on milk production and composition by dairy cows on pasture: a review. Livest Prod Sci 2004; 86(1-3):1-18. [ Links ]
14. Hammon H, Metges C, Junghans P, Becker F, Bellmann O, Schneider F, et al. Metabolic changes and net portal flux in dairy cows fed a ration containing rumen-protected fat as compared to a control diet. J Dairy Sci 2008; 91(1):208. [ Links ]
15. Adewuyi A, Gruys E, Van Eerdenburg F. Non esterified fatty acids (NEFA) in dairy cattle. A review. Vet Quart 2005; 27(3):117-26. [ Links ]
16. Doepel L, Lapierre H, Kennelly JJ. Peripartum Performance and Metabolism of Dairy Cows in Response to Prepartum Energy and Protein Intake. J Dairy Sci 2002; 85(9):2315-34. [ Links ]
17. Heravi Moussavi A, Gilbert R, Overton T, Bauman D, Butler W. Effects of feeding fish meal and n-3 fatty acids on ovarian and uterine responses in early lactating dairy cows. J Dairy Sci 2007; 90(1):145-54. [ Links ]
18. Kitessa S, Gulati S, Simos G, Ashes J, Scott T, Fleck E, et al. Supplementation of grazing dairy cows with rumen-protected tuna oil enriches milk fat with n-3 fatty acids without affecting milk production or sensory characteristics. British J Nutrit 2007; 91(02):271-7. [ Links ]
19. Schroeder G, Delahoy J, Vidaurreta I, Bargo F, Gagliostro G, Muller L. Milk fatty acid composition of cows fed a total mixed ration or pasture plus concentrates replacing corn with fat. J Dairy Sci 2003; 86(10):3237-48. [ Links ]
20. Harvatine K, Allen M. Effects of fatty acid supplements on milk yield and energy balance of lactating dairy cows. J Dairy Sci 2006; 89(3):1081. [ Links ]
21. Doreau M, Chilliard Y. Digestion and metabolism of dietary fat in farm animals. British J Nutrit 2007; 78(01):15-35. [ Links ]
22. González F, Bas F. Efecto de la suplementación con un aceite hidrogenado de pescado sobre la producción de leche en vacas Holstein Friesian. Cien Invest Agrop 2002; 29(2):73-82. [ Links ]
23. Calvopiña AI, León V. Estudio de la suplementación de tres niveles de grasa sobrepasante en la alimentación de vacas lactantes Holstein Friesian. Rumipamba 2007; XXI(1):1-12. [ Links ]
24. Rudolph MC, McManaman JL, Phang TL, Russell T, Kominsky DJ, Serkova NJ, et al. Metabolic regulation in the lactating mammary gland: a lipid synthesizing machine. Physiolog Genom 2007; 28(3):323-36. [ Links ]
25. Bernard L, Rouel J, Leroux C, Ferlay A, Faulconnier Y, Legrand P, et al. Mammary lipid metabolism and milk fatty acid secretion in alpine goats fed vegetable lipids. J Dairy Sci 2005; 88(4):1478-89. [ Links ]
26. Perfield JW, Bernal-Santos G, Overton TR, Bauman DE. Effects of Dietary Supplementation of Rumen-Protected Conjugated Linoleic Acid in Dairy Cows during Established Lactation. J Dairy Sci 2002; 85(10):2609-17. [ Links ]
27. Ceballos A, Gómez P, Vélez M, Villa N, López L. Variación de los indicadores bioquímicos del balance de energía según el estado productivo en bovinos lecheros de Manizales, Colombia. Rev Colomb Cienc Pecu 2009; 15(1):13. [ Links ]
28. Castañeda-Gutiérrez E, Pelton S, Gilbert R, Butler W. Effect of peripartum dietary energy supplementation of dairy cows on metabolites, liver function and reproductive variables. Ani Reprod Sci 2009; 112(3-4):301-15. [ Links ]
29. Leroy J, Van Soom A, Opsomer G, Bols R. The consequences of metabolic changes in high-yielding dairy cows on oocyte and embryo quality. Animal 2008; 2:1120-7. [ Links ]
30. Mattos R, Staples CR, Thatcher WW. Effects of dietary fatty acids on reproduction in ruminants. Rev Reprod 2000; 5(1):38-45. [ Links ]
