










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista MVZ Córdoba
Print version ISSN 0122-0268
Rev.MVZ Cordoba vol.18 supl.1 Córdoba Dec. 2013
ORIGINAL
Oferta de pasto kikuyo (Pennisetum clandestinum), tercio de lactancia y perfil de ácidos grasos lácteos
Kikuyo grass (Pennisetum clandestinum) allowance, state of lactation and milk fatty acid profile
Juan Vargas M,1 Zoot, José Mojica R,2 M.Sc, Martha Pabón R,3 Ph.D, Juan Carulla F,1* Ph.D..
1Universidad Nacional de Colombia, Facultad de Medicina Veterinaria y Zootecnia, Departamento de Ciencias para la Producción Animal, Bogotá, Colombia.
2Centro de investigación Motilonia, Corporación Colombiana de Investigación, Corpoica. Bogotá, Colombia
3Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Química, Bogotá, Colombia.
*Correspondencia: jecarullaf@unal.edu.co
Recibido: Marzo de 2012; Aceptado: Febrero de 2013.
RESUMEN
Objetivo. Evaluar el efecto de la oferta de kikuyo y el tercio de lactancia sobre el perfil de ácidos grasos en la grasa láctea de bovinos en pastoreo. Materiales y métodos. Se utilizaron 18 vacas de la raza Holstein con un peso promedio de 585 ±10 kg entre dos y cuatro partos, en primer y segundo tercio de lactancia. Las vacas se dividieron en tres grupos y fueron asignados al azar a uno de los tres tratamientos (ofertas de 2.6; 3.3 ó 4.0 kg MS/100 kg PV). Se determinó el perfil de ácidos grasos en la leche los días 14 y 21 del periodo experimental. Resultados. Los ácidos grasos C6:0, C16:0, C18:1c9 y C18:3 permanecieron constantes entre las diferentes ofertas de forraje y tercios de lactancia. En las mayores ofertas se presentó un aumento entre el día 14 y 21 de los ácidos C10:0 y C12:0 mientras que disminuyeron su concentración en estos mismos días los ácidos C18:1t11, C18:2c9c12, C18:2c9t11 y poliinsaturados. Se presentó una mayor concentración del ALC en animales de segundo tercio respecto a los de primero. Conclusiones. Debido a las variaciones entre los días de muestreo en el perfil de ácidos grasos de la grasa láctea fue imposible determinar si el aumento en la oferta mejora las concentraciones de ácidos grasos que han presentado efecto benéfico en la salud humana. La movilización de reservas de grasa podría explicar el comportamiento diferente de la concentración de ácidos grasos entre los días de recolección de muestras.
Palabras clave: Ácido linoleico conjugado (ALC), ácido transvaccénico, ácidos poliinsaturados, grasa láctea (Fuente: CAB).ABSTRACT
Objective. To evaluate the effect of kikuyu offer and stage of lactation on fatty acids profile on milk fat of grazing dairy bovines. Materials and methods. Eighteen Holstein cows with average weight of 585±10 Kg in two to four calvings on first and second lactation stages. Cows were divided in three groups and assigned randomly to one of three treatments (pasture allowances of 2.6, 3.3 or 4.0 KgDM/100 Kg LW). Milk fatty acid profiles were determined on days 14 and 21 of the experimental period. Results. Fatty acids C6:0, C16:0, C18:1cis9, C18:3 remained constant between different forage allowances and lactation stage were used. On the treatments of highest allowance, there was an increase between days 14 and 21 of C10:0 andC12:0 acids while C18:1t11,C18:2c9c12, C18:2c9,t11, and polyunsaturated fatty acids decreased their concentration. The lowest allowance had a different behavior. A higher concentration of ALC was observed in animals of second stage compared to first stage of lactation. Conclusions. It was not possible to conclude on the effects of forage allowance on the fatty acid profile of milk, due to variations between sampling dates. It is possible that mobilization of fat storage can explain the different behavior of fatty acid concentrations between sampling days.
Key words: Conjugated linoleic acid (CLA), milk fat, polyunsaturatedfattyacids, transvaccenic acid (Source: CAB).INTRODUCCIÓN
En las últimas décadas se han reconocido en la grasa láctea biomoléculas benéficas para la salud, entre las que se destacan el ácido butírico (C4:0), caprílico (C8:0), cáprico (C10:0), oléico (C18:1), ácidos omega 3 y 6, ácidovaccénico (C18:1 trans-11) y ácido linoléico conjugado (ALC C18:2 c9 t11), (1,2).
Algunos investigadores han sugerido que al aumentar los días de lactancia se presenta variación en la composición de la grasa láctea, evidenciándose en los primeros días una mayor concentración de ácidos grasos de cadena larga provenientes de la movilización de las reservas corporales, y posteriormente el incremento en los ácidos que se obtienen por síntesis de novo (3,4). Kay et al (5), reportaron un incremento (51%) en la concentración de ácidos provenientes de la síntesis de novo (4:0 a 14:0), mientras que los ácidos grasos preformados (≥17:0) disminuyeron (15%) al aumentar los días en lactancia. Stoopet et al (6), reportaron que en la primera parte de la lactancia aumentan las concentraciones de ácidos saturados y disminuyen en la segunda mitad. Estos autores encontraron un aumento en la concentración de ALC en la medida que se incrementaban los días en lactancia.
La literatura reporta algunas estrategias que permiten modificar el perfil lipídico de la grasa láctea de bovinos, tanto en dietas TMR como en pastoreo (7,8). Los sistemas pastoriles han presentado mayor concentración de ciertos ácidos grasos benéficos para la salud como el ALC y ácidos poliinsaturados y una menor de ácidos grasos saturados (7,9). Sin embargo, en pocos trabajos se ha determinado el efecto de la oferta forrajera sobre el perfil lipídico de la grasa láctea. Bargo et al (3) reportaron que con un aumento en la oferta forrajera de 3.96 a 6.34 kg de materia seca (MS)/ 100 kg peso vivo (PV), aumentaban las concentraciones de ácidos grasos de cadena media y corta, saturados e insaturados y que disminuían las concentraciones de ácidos grasos de cadena larga, pero no se encontraron diferencias en las concentraciones de ALC.
El objetivo del presente estudio fue evaluar el efecto de la oferta forrajera de kikuyo y el tercio de lactancia, sobre el perfil lipídico de la grasa láctea, principalmente la concentración de ácidos grasos benéficos para la salud (ácido vaccénico, ácido linoleico conjugado y ácidos grasos poliinsaturados).
MATERIALES Y MÉTODOS
Localización. El experimento se realizó en el Centro Agropecuario Marengo de la Universidad Nacional sede Bogotá, localizado en la vereda San José, municipio de Mosquera (Cundinamarca), 4°42’ de latitud norte y 74°12’ de longitud oeste a una altitud de 2650 msnm con una temperatura promedio de 13°C con fluctuaciones entre 0 y 20°C y una humedad relativa de 80 a 85%. La precipitación anual promedio fue de 528.9 mm, con distribución bimodal del período lluvioso; uno entre los meses de abril y mayo y el otro desde septiembre hasta noviembre.
Diseño experimental. Se evaluaron ofertas de 2.6; 3.3 ó 4.0 Kg MS/100 Kg PV de pasto kikuyo. Los animales se suplementaron de acuerdo con la producción de leche (1 Kg por 4.2 L de leche) utilizando un suplemento diseñado para el experimento (10).
Se seleccionaron 18 vacas Holstein con un peso de 585±10 kg entre dos y cuatro partos, en primer y segundo tercio de lactancia. Las vacas se dividieron en tres grupos teniendo en cuenta la producción de leche y los días en lactancia donde cada uno de los grupos tenía tres vacas en primer tercio y tres en segundo tercio. Uno de los tres tratamientos experimentales fue asignado al azar a cada uno de los grupos.
La duración del período experimental fue de 21 días. Los primeros 7 días se utilizaron como período de ajuste de los animales al suplemento, los 7 días siguientes de ajuste a la oferta de pasto kikuyo y los últimos 7 días fueron utilizados para el período de medición.
Manejo del pastoreo. Se utilizó una pastura de kikuyo (Pennisetum clandestinum) con un área aproximada de 3 ha dividida en tres áreas de 1 ha, donde se distribuyeron al azar los tres grupos de animales. El kikuyo fue pastoreado a una edad de rebrote de 45 días. Para la evaluación de la producción de forraje verde se realizaron cortes a una altura de pastoreo de 10 cm por delante de la cuerda eléctrica utilizando un marco plástico de 1 m2. Se tomaron muestras de forraje para análisis de materia seca (MS), cada segundo día a partir del período de acostumbramiento y para el análisis de la composición química del forraje a los días 14, 17 y 20 del período experimental (10). La producción de forraje promedio fue de 2000 kg de MS/ha.
Se ajustó diariamente la oferta forrajera mediante el uso de cuerda eléctrica ofreciendo el área de pastoreo en metros cuadrados de superficie de pasto kikuyo en cada grupo experimental correspondiente a las ofertas de 2.6; 3.3 y 4.0 kg MS/100 kg PV, respectivamente, moviendo dos veces diarias la cuerda eléctrica, luego de cada ordeño (a.m. y p.m.).
Toma de muestras y análisis químicos. Muestras de forraje y concentrado fueron secadas en horno de aire forzado, a 60°C durante 48 horas. Posteriormente se molieron usando una criba de 1 mm de diámetro. Se determinó humedad (11), fibra detergente neutro (FDN), fibra detergente ácido (FDA) (12), proteína cruda por el método de Kjeldahl (11) y grasa total por extracción con éter (11).
La producción de leche se midió los días 0, 14 y 21 en el ordeño de la mañana y de la tarde. Para cada día de muestreo, se tomaron muestras de leche en cada ordeño (a.m. y p.m.) y posteriormente se mezclaron en partes iguales. Estas muestras fueron conservadas con bicromato de potasio en una concentración de 600 ppm. Para obtener la crema, la leche se centrifugó a 5000 rpm durante 15 min y la crema se congeló a -20°C hasta su análisis posterior (13).
Extracción de grasa y metilación de los ácidos grasos. Para la extracción de la grasa láctea, se descongeló la crema a 30° centígrados. Se extrajo la grasa de la leche a través de una técnica de separación mecánica modificada (14). Se colocaron 500 µl de la crema homogenizada en un tubo de ensayo. Se adicionaron 500 µl de una solución detergente preparada con 50 gramos de hexametafosfato de sodio y 24 ml de Tritón X-100® disueltos en un litro de agua. El tubo de ensayo se agitó vigorosamente, se colocó en un baño de agua a 90°C y se extrajo la capa superior que corresponde a la fracción lipídica. De ésta se tomaron 100 µl y se disolvieron en 1000 µl de una solución cloroformo: metanol, en relación 1:1, y se agitó con vortex. Para preparar los esteres metílicos, se tomaron 20 µl de la solución anterior y se mezclaron con 160 µl de la solución cloroformo: metanol, y 20 µl de MethPrep II®. La muestra se conservó a 20°C para su posterior análisis por cromatografía de gases.
Para la extracción de los lípidos del kikuyo (Pennisetum clandestinum) se utilizó el protocolo adaptado de Palmquist (15). Se pesó un gramo del forraje seco dentro de un tubo de ensayo. Luego se adicionaron 2 ml de tolueno y 3 ml de solución HClmetanólica al 10% (v/v), (se adicionaron 10 ml de HCl a 90 ml de metanol), se mezcló y se colocó en un baño de maría a 65°C durante dos horas. Después de este tiempo se dejó enfriar, se adicionaron 5 ml de hexano y 10 ml de una solución de carbonato de potasio al 6% (p/v), se mezcló y se centrifugó por 5 minutos a 500 g. La capa superior (solvente) se transfirió a tubos de ensayo que contenían 1 g de carbón activado y 1 g de sulfato de sodio y se centrifugó por 5 minutos a 500 g. El sobrenadante que contiene los lípidos se pasó por una micro columna que contenía 0.5 cm de sulfato de sodio en la parte superior de la columna, 6 cm de sílica gel para cromatografía en la parte intermedia y finalmente 2 cm de carbón activado, en la parte inferior. La columna se eluyó con hexano, se eliminó el exceso de hexano del eluido, la muestra se predisolvió en 0.5 ml de hexano y se transfirió a viales ámbar para su posterior análisis cromatográfico.
Análisis de los esteres metílicos mediante cromatografía de gases. Se hicieron dos análisis cromatográficos. El primero, para cuantificar los ácidos grasos diferentes al ácido vaccénico y a los ALC’s, fue realizado con una fase móvil de helio, en una columna capilar Omega wax 320 Supelco® de 30 metros de longitud y 0.25 mm de diámetro interno. Se utilizó un programa isotérmico de 80°C por 2 min, pendiente de 10°C/min hasta los 190°C, durante 20 min, pendiente de 2°C hasta los 220°C, durante 10 min y un tiempo total de corrida de 58 min. En el segundo análisis cromatográfico se determinó el ácido vaccénico (C18:1 trans11) y el ALC (C18:2 c9t11), utilizando una columna capilar en sílica fundida con fase polar D-Wax Agilen®, de 30 metros de longitud y 0.25 mm de diámetro interno con un programa de temperatura de 190°C isotérmica por 2 min, pendiente 3°C por min hasta los 220°C, durante 3 min y un tiempo total de la corrida de 15 min.
La muestra que contenía los esteres metílicos de los ácidos grasos fue inyectada en un cromatógrafo (Shimadzu C-R4A) equipado con un detector de ionización de llama (FID). Cada pico fue identificado y cuantificado utilizando como referencia los tiempos de retención de los patrones. El cálculo de la concentración de cada ácido graso se realizó tomando el área de cada pico de manera individual y relacionando con el número de áreas detectadas. Se utilizaron patrones comerciales para la determinación del perfil lipídico (SupelcoTM.Bellefonte, PA), vaccénicotrans 11 (Nu-Check®.Elysian, MA), y ALC’s, c9t11 y t10c12. (Nu-Check®. Elysian, MA).
Análisis estadístico. Para el análisis del efecto de la oferta forrajera y el tercio de lactancia sobre la concentración de los ácidos grasos se utilizó un modelo de medidas repetidas con arreglo factorial 2x3 (dos tercios de lactancia y tres tratamientos). Se analizaron los efectos principales de los tratamientos, el tercio de lactancia y el día de toma de muestra; además, de las interacciones utilizando el procedimiento mixed del programa SAS (16). La comparación de medias se realizó por medio de la prueba de Duncan con un nivel de significancia del 5% (17).
RESULTADOS
Perfil lipídico del forraje y suplemento alimenticio. Los ácidos grasos más abundantes en el kikuyo (Pennisetum clandestinum) fueron en su orden linolénico (18:3), palmítico (16:0), linoleico (C18:2c9c12) y esteárico (C18:0). El suplemento alimenticio presentó una mayor concentración de linoléico (C18:2c9c12), oleico (C18:1) y palmítico (C16:0) (Tabla 1).
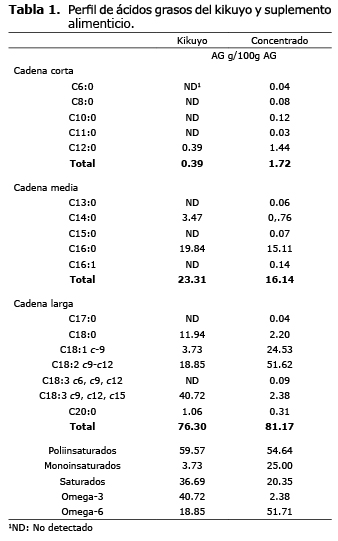
Perfil lipídico de la grasa láctea. El tercio de lactancia, la oferta forrajera y el día de la toma de muestra no tuvieron ningún efecto sobre las concentraciones de los ácidos capróico (C6:0), palmítico (C16:0), oleico (C18:1c9) y linolénico (C18:3 c9c12c15) en la leche. Adicionalmente, el efecto de los factores sobre las concentraciones de los demás ácidos grasos no se explicaron de manera independiente sino que se presentaron interacciones, principalmente entre la oferta forrajera y el día de muestreo. La respuesta del ácido mirístico (C14:0) a los tratamientos no fue clara pues se presentó una interacción triple (p<0.05)(Tabla 2).
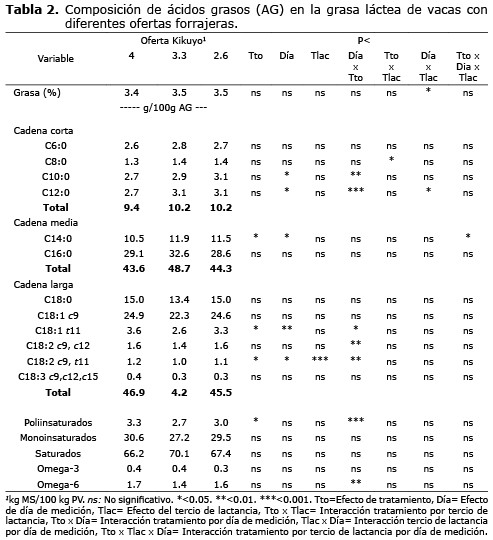
Con una oferta de forraje del 4% se presentó una menor concentración de ácido caprílico (C8:0) en animales de primer tercio de lactancia (1.29 g/100 g de AG ) (p<0.05) mientras que no hubo diferencias entre ofertas de 3.3 y 2.6% (1.46 y 1.43 mg/g de grasa respectivamente). En animales de segundo tercio de lactancia, se evidenció diferencias entre todos los niveles de oferta (p<0.05) resultando en una menor concentración para la mayor oferta (1.32 g/100 g de AG) y un aumento de la concentración conforme disminuía la oferta de forraje (1.37 y 1.41 g/100 g de AG , para la oferta media y baja, respectivamente).
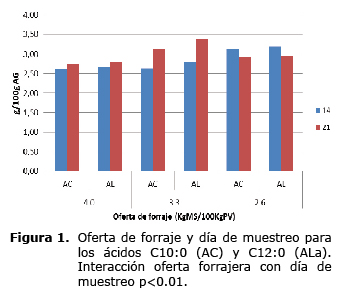
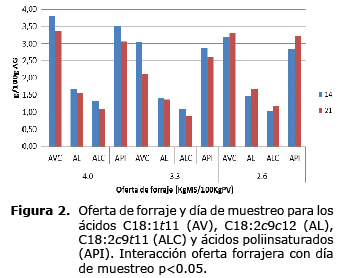
La respuesta a la oferta en la concentración de los ácidos grasos cáprico (C10:0), laúrico (C12:0), vaccénico (C18:1t11), linoléico (C18:2c9c12), ALC (C18:2c9t11) y poliinsaturados, dependieron del día de muestreo. En el día 14, la concentración de ácido cáprico (C10:0) y laúrico (C12:0) fueron menores para las ofertas de 4.0 y 3.3% y mayores para la oferta de 2.6% (p<0.01). Para el día 21 se observó un aumento en las concentraciones de éstos ácidos para las ofertas de 4.0 y 3.3% y una disminución en la oferta de 2.6% (Figura 1). En el día 14, la concentración de los ácidos grasos vaccénico (C18:1t11), linoléico (C18:2c9c12), ALC (C18:2c9t11) y poliinsaturados, fueron mayores en la oferta de 4% que en las menores ofertas. En el día 21, se presentó un aumento en la concentración de éstos ácidos grasos en la menor oferta de forraje (2.6%) mientras la concentración disminuyó en las mayores ofertas (3.3 y 4%) (Figura 2).
Se evidenció un aumento en las concentraciones de ALC en animales de segundo tercio respecto a los del primero (Tabla 2). También se presentaron variaciones en las concentraciones del ácido laúrico (C12:0) asociadas al día de muestreo y el tercio de lactancia (interacción p<0.05). En el día 14 no se presentaron diferencias en la concentración de ácido láurico entre animales de primero y segundo tercio. Sin embargo, en el día 21 el primer tercio aumentó las concentraciones del ácido laúrico (3.2 g/100 g de AG) respecto al segundo tercio (2.9 g/100 g de AG).
Se presentó una correlación negativa entre los ácidos de cadena corta frente a los de cadena larga y poliinsaturados (r=-0.51, p<0.03 y r=-0.63, p<0.004, respectivamente).
DISCUSIÓN
Forrajes y suplemento. En el presente estudio la proporción de ácido linolénico en los ácidos grasos del kikuyo fue de 41%. Sin embargo, Elgersma et al (18) reportaron que el contenido de ácido linolénico en forrajes verdes puede oscilar entre el 50 y el 70% del total de ácidos grasos. Estas variaciones pueden deberse a la edad de corte y/o condiciones ambientales (8,18). Es el caso de Aguilar et al (8), quienes reportaron una disminución del 10% en la concentración de ácido linolénico en kikuyo (Pennisetum clandestinum) de 70 días respecto a los de 50 días. La concentración de ácidos saturados fue similar a lo reportado por Aguilar et al (8) en pasto kikuyo y Bargo et al (19) en una mezcla de diferentes especies forrajeras.
El perfil lipídico del suplemento fue similar a lo encontrado por Elgersma et al (20,21). Sin embargo, Bargo et al (19) reportaron una mayor concentración de palmítico (C16:0) y oléico (C18:1) que de linoléico. Estas diferencias pueden ser debidas a las materias primas usadas en la preparación del suplemento alimenticio (10).
Grasa láctea. En el presente estudio, la concentración de ácidos grasos de cadena corta y media fueron similares a los reportados por Aguilar et al, (8) y Wijesundera et al (20), los de cadena larga, a lo reportados por Kelsey et al (4) y Stockdale et al (22). Sin embargo, se presentaron variaciones de acuerdo con los tratamientos evaluados, aunque la concentración de grasa fue similar entre estudios.
Oferta forrajera. La mayoría de los ácidos grasos en la leche no presentaron variaciones debido a la oferta forrajera únicamente, los ácidos grasos caprílico (C8:0), mirístico (C14:0), vaccénico (C18:1t11), ALC (C18:2c9t11) y poliinsaturados mostraron variaciones asociadas a la oferta que dependieron del día de muestreo o del tercio de lactancia. Aunque para el primer muestreo las concentraciones de los ácidos grasos deseables para la salud humana (vaccénico, ALC y poliinsaturados) fueron mayores en el tratamiento de mayor oferta estos disminuyeron su concentración entre los dos días de muestreo contrario al comportamiento de los animales que recibieron la menor oferta. Por lo tanto, es difícil concluir sobre la bondad de aumentar la oferta de kikuyo para aumentar estos ácidos en la leche. Lo hallazgos en la literatura también parecen contradictorios. Wisejundera et al (20) encontraron un aumento del 6 y 13% en la concentración de ácido vaccénico y ALC respectivamente y una disminución de 14% en la concentración de linoleico, cuando aumentaba la oferta forrajera en una pastura de mezcla de gramíneas desde 3.7 a 7.6 kg de MS/ 100 kg PV. Bargo et al (19) también encontraron una disminución en la concentración de linoleico y oleico y no observaron diferencias en el vaccénico o ALC. Estos resultados difieren a los encontrados por Stockdale et al (22), quienes no reportan efecto de la oferta forrajera sobre los ácidos grasos antes mencionados, debido quizás a una menor oferta de forraje en sus experimentos, (4 vs. 6 kg de MS/ 100 kg PV) o un mejor estatus energético de los animales. Estos autores sugieren que el aumento en la concentración de los ácidos grasos de cadena larga en animales que consumen menor cantidad de forraje, podría deberse a un balance energético negativo, lo que conduce a una movilización de tejido graso y mayor concentración de éstos ácidos en la grasa láctea (23).
La concentración de los ácidos caprílico (C8:0), cáprico (C10:0), laúrico (C12:0) y mirístico (C14:0), fueron menores en animales que consumieron una mayor oferta de forraje. Este efecto se podría explicar por un mayor consumo de ácidos grasos de cadena larga. Algunos autores sugieren que un mayor consumo de ácidos grasos de cadena larga inhibe la síntesis de ácidos grasos de cadena corta; de esta manera un mayor consumo de forraje podría disminuir la concentración de ácidos grasos de cadena corta (23).
Con la menor oferta de forraje se aumentaron las concentraciones de los ácidos vaccénico (C18:1t11), linoléico (C18:2c9c12), ALC (C18:2c9t11) y poliinsaturados entre la primera y la segunda toma; por el contrario con mayores ofertas se disminuyeron las concentraciones de éstos ácidos grasos. La disminución de estos ácidos en aquellos animales que recibieron una mayor oferta podría ser debida a un descenso en la concentración de ácidos grasos precursores en el forraje, ya que el forraje ofrecido en la primera toma, era 7 días más joven que el de la segunda. Aguilar et al (8), reportaron que en pasto kikuyo a mayor edad disminuye la concentración de precursores, especialmente ácido linolénico (C18:3) y se presenta un aumento en ácidos grasos saturados. Sin embargo, esto no explicaría el aumento en la menor oferta de forraje, lo que sugeriría un aumento en el aporte endógeno por posible movilización de tejido graso en animales que recibieron una menor oferta.
Bargo et al (19), Griinari et al (24) y Moate et al (7), sugieren que la concentración de ácidos grasos provenientes de la síntesis de novo presenta una correlación negativa en relación a la concentración de ácidos grasos poliinsaturados; resultados similares se encontraron en este trabajo. Sin embargo, estos resultados difieren del trabajo realizado por Aguilar et al (8), en el cual la síntesis de ácidos grasos de cadena corta no presentó una correlación negativa con las concentraciones de ácidos grasos poliinsaturados.
Tercio de lactancia. La literatura reporta que a medida que avanzan los días en lactancia, aumenta la concentración de ácidos grasos de cadena corta (25) pero en la semana 8 se estabiliza la concentración de éstos ácidos grasos seguido de un descenso que ocurre aproximadamente a los 3 meses de lactancia (26). Lo anterior podría explicar la mayor concentración de ácido caprílico (C8:0) en animales de primer tercio, respecto a los de segundo, debido a que los animales de primer tercio de lactancia superaban los 56 días de lactancia, logrando una mayor concentración, y los de segundo tercio superaban los 120 días, disminuyéndola levemente.
En este estudio, el perfil de ácidos grasos, con excepción al ALC, no fue modificado por el tercio de lactancia. El ALC presentó mayores concentraciones en animales de segundo tercio respecto animales de primer tercio de lactancia. Resultados similares reportó Kay et al (25) y Stoop et al (26) en donde las concentraciones de ALC incrementaron al aumentar los días de lactancia. Los resultados de este trabajo sugerirían que los ácidos grasos varían poco debido al tercio de lactancia, contrario a lo reportado por otros autores que proponen que el aporte de ácidos grasos endógenos es mayor en el primer tercio de lactancia y los provenientes de la síntesis de novo aumentan en el segundo y tercer tercio, relacionado con un mejoramiento en el balance energético (25).
Este trabajo intentó determinar el efecto de la oferta forrajera y el tercio de lactancia sobre la composición de la grasa de la leche. Los resultados sugieren que la respuesta varía dependiendo del ácido graso y del día de toma de muestra. Mientras los ácidos grasos C6:0, C16:0, C18:1 c9 y C18:3 permanecieron constantes entre las diferentes ofertas de forraje y tercios de lactancia, los ácidos grasos C10:0, C12:0, C18:1t11, C18:2 c9c12, C18:2c9t11 y poliinsaturados fueron afectados por las diferentes ofertas de forraje y el día de toma de muestra. En los tratamientos de mayor oferta se presentó un aumento entre el día 14 y 21 de para los ácidos C10:0 y C12:0 mientras que disminuyeron su concentración en estos mismos días los ácidos C18:1t11, C18:2c9c12, C18:2c9t11 y poliinsaturados. Diferente comportamiento presentó la menor oferta forrajera. El tercio de lactancia afectó la respuesta del ácido C18:2c9t11 presentándose mayor concentración en animales de segundo tercio respecto a los de primero. Posiblemente, la movilización de reservas corporales, puede explicar el diferente comportamiento, entre los días de toma de muestra, para los animales que recibieron una menor oferta de forraje respecto a aquellos de mayor oferta.
REFERENCIAS
1. Rico J, Moreno B, Pabón M, Carulla J. Composición de la grasa láctea de la sabana de Bogotá con énfasis en ácido ruménico- CLA cis9 trans11. Rev Col Cienc Pecu 2007; 20:30-39. [ Links ]
2. Gagliostro G. Control nutricional del contenido de ácido linoléico conjugado (CLA) en leche y su presencia en alimentos naturales funcionales. 2. Producción de leche alto CLA de vaca. Rev Arg Prod Anim 2005; 24(3-4):137-163. [ Links ]
3. Bargo F, Delahoy J, Schroeder G, Muller L. Milk fatty acid composition of dairy cows grazing at two pasture allowances and supplemented with different levels and sources of concentrate. Anim Feed Sci Technol 2006; 125:17-31. [ Links ]
4. Kelsey J, Corl B, Collier R, Bauman D. The effect of breed, parity and stage of lactation on conjugated linoleic acid (CLA) in milk fat from dairy cows. J Dairy Sci 2003; 86:2588-2597. [ Links ]
5. Kay JK, Weber WJ, Moore CE, Bauman DE, Hansen LB, Chester-Jones H, et al. Effects of week of lactation and genetic selection for milk yield on milk fatty acid composition in Holstein Cows. J Dairy Sci 2005; 88:3886-3893. [ Links ]
6. Stoop W, Bovenhuis H, Heck J, van Arendonk J. Effect of lactation stage and enegy status on milk fat composition of Holstein-Friesian cows. J Dairy Sci 2009; 92:1469-1478. [ Links ]
7. Moate P, Chalupa E, Bostom R, Lean I. Milk fatty acids I: Variation in the concentration of individual fatty acids bovine milk. J Dairy Sci 2007; 90:4730-4739. [ Links ]
8. Aguilar O, Moreno B, Cárdenas E, Pabón M Carulla J. Efecto del consumo de kikuyo (Pennisetumclandestinum) o ryegrass (Lollium spp.) sobre la concentración de ácido linoléico conjugado y el perfil de ácidos grasos de la grasa láctea. Livest Res Rural Dev 2009; 21(49). [ Links ]
9. Elgersma A, Tamminga S, Ellen G. Effect of grazing versus stall-feeding of cut grass on milk fatty acid composition of dairy cows. Proceedings of the Int. Occ. Symp. of the European Grassland Federation, Pleven, Bulgaria. Grass Sci Eur 2003; 8:271-274. [ Links ]
10. Mojica JE, Castro E, León JM, Cárdenas EA, Pabón ML, Carulla JE. Efecto de la oferta de pasto kikuyo y ensilaje de avena sobre la producción y calidad composicional de la leche bovina. Rev Corpoica 2009; 10:81-90. [ Links ]
11. A.O.A.C. Official methods of analysis. Washington, D.C., USA: Association of Official Agricultural Chemists. 2005. [ Links ]
12. Van Soest P, Roberton L. Methods for dietary fiber, neutral fiber and no starch polysaccharides in relation to nutrition. J Dairy Sci 1991; 74:3583-3597. [ Links ]
13. Tilley J, Terry A. Two-stage technique for in vitro digestion of forage crops. J Br Grass Soc 1963; 18:104-111. [ Links ]
14. Díaz-González G, Gutiérrez R, Pérez N, Vega S, González M, Prado G, et al. Detección de adulteraciones de la grasa de la leche pasteurizada mexicana. Rev Salud Anim 2002; 24:54-59. [ Links ]
15. Palmquist DL. The feed value of fats. In: Orskov, E Editor. Feed Science. Amsterdam: Elsevier Science Publishing. 1983. [ Links ]
16. SAS. OnlineDoc 9.1.3. Cary, USA: SAS Institute Inc; 2007. [ Links ]
17. Martínez R, Martínez N, Martínez MV. Diseños de experimentos en ciencias agropecuarias y biológicas con SAS, SPSS, R y STATISTIX. 1 Ed. Bogotá (Colombia): Cooperativa de Profesores de la Universidad Nacional de Colombia; 2011. [ Links ]
18. Elgersma A, Tamminga S, Ellen, G. Modifying milk composition through forage. Anim Feed Sci 2006; 131:207-225. [ Links ]
19. Bargo F, Delahoy J, Schroeder G, Muller L. Milk fatty acid composition of dairy cows grazing at two pasture allowances and supplemented with different levels and sources of concentrate. Anim Feed Sci Technol 2006; 125:17-31. [ Links ]
20. Wijesundera C, Shen Z, Wales W, Dalley D. Effect of cereal grain and fibre supplements on the fatty acid composition of milk fat of grazing dairy cows in early lactation. J Dairy Res 2003; 70:257-265. [ Links ]
21. Elgersma A, Ellen G, van der Horst H, Boer H, Dekker P y Tamminga S. Quick changes in milk fat composition from cows after transition from fresh grass to a silage diet. Anim Feed Sci Technol 2004; 117:13-27. [ Links ]
22. Stockdale R, Walker G, Wales W, Dalley D, Birkett A, Shen Z et al. Influence of pasture and concentrates in the diet of grazing dairy cows on the fatty acid composition of milk. J Dairy Res 2003; 70:267-276. [ Links ]
23. Khanal R, Dhiman T, Boman R. Changes in fatty acid composition of milk from lactating dairy cows during transition to and from pasture. Livest Sci 2007; 114:164-175. [ Links ]
24. Griinari J, Corl B, Lacy S, Chouinard P, Nurmela K, Bauman D. Conjugated linoleic acid is synthesized endogenously in lactation dairy cows by Δ9 desaturase. J Nutr 2000; 130:2285-2291. [ Links ]
25. Kay JK, Weber WJ, Moore CE, Bauman DE, Hansen LB, Chester-Jones H, et al. Effects of week of lactation and genetic selection for milk yield on milk fatty acid composition in Holstein Cows. J Dairy Sci 2005; 88:3886-3893. [ Links ]
26. Stoop WM, Bovenhuis H, Heck JML, van Arendonk JAM. Effect of lactation stage and energy status on milk fat composition of Holstein Friesian cows. J Dairy Sci 2009; 92:1469-1478. [ Links ]
