










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista MVZ Córdoba
Print version ISSN 0122-0268
Rev.MVZ Cordoba vol.19 no.3 Córdoba Sept./Dec. 2014
ORIGINAL
Association of the bovine growth hormone gene with Holstein cattle reproductive parameters
Asociación del gen de la hormona de crecimiento bovino con parámetros reproductivos en ganado Holstein
Juliana Arango G,* M.Sc, Julián Echeverri Z, Ph.D, Albeiro López H, Ph.D.
1Universidad Nacional de Colombia, Facultad de Ciencias Agropecuarias, Departamento de Producción Animal, Grupo BIOGEM, Sede Medellín, AA 1779, Colombia.
*Correspondence: jarangog@unal.edu.co
Received: November 2013; Accepted: March 2014.
ABSTRACT
Objective. To determine the association of intron 3 polymorphism of the bovine growth hormone (BGH) gene, with age at first service, first birth, first postpartum service and second birth in a population of Holstein cows in the state of Antioquia. Materials and methods. The study was conducted on 408 Holstein cows in 8 herds. Genotyping was performed using PCR-RFLP technique with DNA extracted from peripheral blood by the salting out technique. The Phenotypic information used was collected for 4 years, from a dairy production control program. To determine the association between characteristics and gene polymorphism, parametric statistical analyzes were performed as generalized linear models and linear regression analysis. Results. Allele frequency (positive) was 0.91 and was 0.09 for (negative) allele. Genotypic frequencies were 0.77, 0.2 and 0.03 for (positive/positive), (positive/negative) and (negative/negative) respectively. There were significant differences between the mean of age at first service, age at first birth, age at first postpartum service and age at second birth. For all of the (negative/negative) genotype characteristics there were greater ages for each event. Conclusions. These results suggest that intron-3 polymorphism of the BGH gene is associated with reproductive traits, facilitating the selection of individuals with favorable genotypes for use in breeding programs.
Key words: Animal reproduction, molecular markers, PCR-RFLP (Source: Mesh).
RESUMEN
Objetivo. Determinar la asociación del polimorfismo del intrón 3 del gen de la hormona de crecimiento bovino (BGH) con las edades al primer servicio, primer parto, primer servicio posparto y segundo parto en una población de vacas Holstein del departamento de Antioquia. Materiales y métodos. El estudio se realizó con 408 vacas Holstein ubicadas en 8 hatos. La genotipificación se llevó a cabo usando la técnica de PCR-RFLP con ADN extraído de sangre periférica mediante la técnica de salting out. La información fenotípica utilizada fue recopilada durante 4 años, a partir de un programa de control de producción lechera. Para determinar la asociación entre las características y el polimorfismo del gen, se realizaron análisis estadísticos paramétricos mediante modelos lineales generalizados y análisis de regresión lineal. Resultados. La frecuencia para el alelo (positivo) fue de 0.91 y para el alelo (negativo) fue de 0.09. Las frecuencias genotípicas fueron 0.77, 0.2 y 0.03 para (positivo/positivo), (positivo/negativo) y (negativo/negativo) respectivamente. Se presentaron diferencias significativas entre las medias de la edad al primer servicio, la edad al primer parto, edad al primer servicio postparto y edad al segundo parto. Para todas las características el genotipo (negativo/negativo) presentó edades más tardías para cada eventos. Conclusiones. Estos resultados sugieren que el polimorfismo del intrón 3 del gen BGH, está asociado con características de tipo reproductivo, facilitando la implementación de un programa de selección de individuos con genotipos favorables, para su uso en programas de mejoramiento genético animal.
Palabras clave: Marcadores moleculares, PCR-RFLP, reproducción animal (Fuente: Mesh).
INTRODUCTION
In specialized dairy herds, reproductive efficiency has a direct influence on productivity. Without efficient reproductive performance, milk production is strongly affected and thus the profitability of the herd. Cow fertility is affected by many factors, the age of the animal at puberty has a very strong influence and is transcendental due to the fact that older ages at the start of breeding and first calving reflect low reproductive efficiency (1).
A delay in the age of service or birth of each cow, or even, each cow discarded as a result of infertility affects the economic performance of the herd. When a heifer is delayed in entering the breeding program or a cow delays in returning to heat after delivery, there are expenses for the additional non-lactating period and lengthening of the generational interval (1, 2). In contrast, reduction of the heifers' rearing period and rapid growth, which brings them to the reproductive stage in a shorter amount of time, produces more births and more production during their lifetime. Therefore, it is essential to consider the factors that influence these parameters. The bovine growth hormone (BGH) gene has polymorphisms that influence these characteristics differently (2) and play a key role in the regulation of growth and development; indirectly influencing the reproductive efficiency of cattle (3-7).
BGH effect on growth is observed in several tissues including bone, muscle and adipose tissue. These effects result from both direct action of BGH on nutrient partitioning and on cell multiplication, as in the action mediated by the insulin-like growth factor 1, which stimulates cell proliferation and metabolic processes associated with protein deposition (8). In addition, significant relationships have been reported between BGH concentrations and the postpartum period, and consequently with cow fertility (9.10).
The main concern behind the analysis of this association is to determine the influence of genetic variants of growth hormone on the scope of an earlier active reproductive stage. The aim of the research was to determine the association of intron 3 polymorphism of the bovine growth hormone (BGH) gene, with ages at first service, first birth, first postpartum service and second birth, in a population of Holstein cows in the state of Antioquia.
MATERIALS AND METHODS
Study areas and population. 408 Holstein cattle were used, belonging to 8 dairy herds located in the high tropics of Antioquia, northern and eastern areas of the state, in the municipalities of San Pedro de los Milagros (height 2475 mamsl; with a temperature of 14°C, Belmira (height 2550 mamsl, T° 14°C) and Santa Elena township of Medellín (height 2500 mamsl, T° 17°C).
DNA extraction and determination of genotypic variants. To determine the genotypic variants, coccygeal vein blood was extracted and DNA extraction was performed using the salting out technique (11). Only genomic DNA with suitable purity between 1.8-2.0 was considered for the studies to be performed.
The following oligonucleotides were synthesized, of 20 base pairs (pb) F 5'CCC ACG GGC AAG AAT GAG GC 3', R5' TGA GGA ACT GCA GGG GCC CA 3', which enabled amplification of the 329bp fragment which has the restriction site for endonuclease MspI (12).
This was carried out by PCR amplification for the specific region, using a final volume of 25 µl containing 2.5 µl 10X PCR buffer (1.0-1.5 mM of MgCl2, 50 mM of KCl, 10 mM of Tris-HCl, pH of 8.3), 0.2 µM of primers; 0.4 mM of each dNTPs, 2 mM of MgCl2, 1 unit of Taq polymerase and ~30-60 ng of genomi DNA.
PCR was performed in a thermocycler (Biometra®). The conditions for amplification of the specific region of the BGH gene were: initial denaturation with heating for five minutes at 94°C, followed by 39 cycles of denaturation at 94°C for 1 minute, primer alignment (anneling) at 55°C for 1 minute, extension at 72°C for 1 min, and a final extension step of 3 minutes at 72°C to complete the reaction (12). The annealing temperature used was determined by following the recommendation of the primer supplier and through different trials that yielded more efficient alignment to said temperature (55°C) (12). As a positive control for all reactions, amplification of samples that were previously evaluated was performed and as negative control reactions in the absence of DNA. Genotypic variants were determined by using RFLP (Restriction Fragment Length Polymorphism), using the restriction enzyme MspI. A final volume of 20 µl was used, containing 5 µl of PCR product, 2 µl of buffer Tango 1X, 12.5 µl of ultra-pure water, which was subjected to digestion by 5 units of restriction enzyme MspI for three hours at 37°C. The resulting product was observed by electrophoresis in agarose gel at 2.5% (w/v) and ethidium bromide staining. The restriction pattern expected for the (negative/negative) genotype was a 329 pb fragment, for the (positive/negative) genotype three fragments (329, 224 and 105 pb) and for the (positive/positive) genotype two fragments (224 and 105 pb).
Estimation of allele frequencies. The frequency of the different alleles was estimated by determining the proportion of each form of the gene among the total number of copies in the study population. Homozygotes (two copies of the same allele) and heterozygous (one copy of each allele) were identified, and the frequency (F) of each allele was calculated by counting homozygotes and adding half the heterozygotes, with the method described by Hartl (13).
Statistical Analysis. To determine the association of each characteristic with the genotype for the BGH, several generalized linear models were adjusted based on known sources of variation for each of the dependent variables (age at first service, age at first birth, age at first postpartum service and age at second birth). The Tukey analysis was used to determine differences between the means for each of the fixed effect levels included in the models. The statistical packet SAS v9.2 was used for all analyzes (SAS Inst Inc., Cary, NC), (14).
The general model of fixed effects was carried out as follows:
Yijklmnopq= µ + Gi+ Hj + ANk + MNl + (MN*AN)m + (H*AN)n + (H*MN)O + (H*MN*AN)P + (H*G)q + eijklmnopq
Where:
Yijklmnopq = age at first service, age at first birth, age at first postpartum service or age at second birth, of individual X, carrier of genotype i, located in herd j, with year of birth k and month of birth l
µ= mean for the characteristic
Gi= fixed effect of the gentoype for BGH (i= 1...3)
Hj= fixed effect of the herd (j = 1â¦8)
ANk= fixed effect of the year of birth (k=1â¦15)
MNl= fixed effect of the month of birth (l=1â¦12)
(MN*AN)m= fixed effect of the interaction between the month of birth and the year of birth (m=1â¦86)
(H*AN)n= fixed effect of the interaction between the herd and the year of birth (n=1â¦46)
(H*MN)o= fixed effect of the interaction between the herd and the month of birth (o=1â¦68)
(H*MN*AN)P = fixed effect of the interaction between the herd, the month of birth and the year of birth (p=1â¦41)
(H*G)q= fixed effect of the interaction between the herd and the genotype (q=1â¦.13)
eijklmnopq = experimental error.
An analysis of simple linear regression was conducted to determine the effect of allelic substitution on each of the study variables. For this, the genotype was converted to a quantitative scale 0, 1 and 2 for (negative/negative), (positive/negative) and (positive/positive), respectively.
The linear regression model used was the following:
Yi = β0 + β1 Xi + ei
Where:
Yi= value of the dependent variable (age at first service, age at first birth, age at first postpartum service and age at second birth) depending on the number of + alleles
β0= intercept
β1= coefficient of linear regression estimated for allele replacement (+)
Xi= number of + alleles in individual i. (0, 1, 2)
ei= experimental error.
RESULTS
Determination of genotype and allele frequencies of the gene for the growth hormone (BGH). A 329pb fragment was amplified from the DNA of 408 animals. The analysis of the restriction fragments using the MspI enzyme originated two restriction patterns; 329 pb, corresponding to the (negative) allele and 224 and 105 pb, corresponding to the (positive) allele. Allele frequencies (positive) and (negative) were 0.91 and 0.09, respectively. The genotype frequencies were 0.77, 0.2 and 0.03 for (positive/positive), (positive/negative) and (negative/negative), respectively.
Descriptive analysis. The average age at first service was 589±91.17 days. For the age at first birth an average of 911±126.2 days was obtained, for the first postpartum service 1013±142.57 days, and for the second birth 1314±138.01 days was obtained. All characteristics showed a relatively low Coefficient of Variation (CV), with the age at first service having the highest value (15.4%) (Table 1). However, these values ââare too high with respect to the reproductive goals that efficient milk production systems should have (1). Descriptive statistics for all characteristics are summarized in table 1.
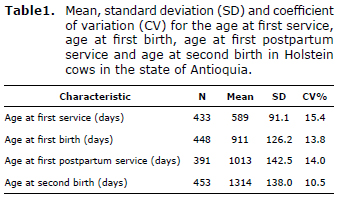
Effect of genotype on the age of reproductive events. The genotype had a highly significant effect (p≤0.01) on age at first service; the model showed a coefficient of determination of 0.88; indicating that the herd, year of birth, month of birth, interaction between the month and year of birth, between the herd and year of birth, herd and month of birth, and interaction between the herd, month and year of birth, explained an 88% variation of the characteristic. The Tukey analysis, which was performed subsequently, showed that heifers with the (negative/negative) genotype took 69 days more for their first service than heifers with the (positive/positive) genotype. Likewise, (positive/negative) individuals were delayed 13 days more than (positive/positive) individuals. Animals with the (negative/negative) genotype had a lower performance because they had their first service at later ages. These results are presented in table 2.

The genotype had a highly significant effect (p≤0.01) on age at first birth and first postpartum service; the model showed a coefficient of determination of 0.81 and 0.83 respectively, indicating that the effects included in the model explained the 81% variation in age at first birth and 83% variation in age at first postpartum service. The Tukey analysis, subsequently carried out, showed significant differences (p≤0.05), denoting the (negative/negative) genotype as the least favorable, as it showed the later age for both characteristics. The analysis of means for the (positive/positive) and (positive/negative) genotypes showed no significant differences (p>0.05, Table 2).
The genotype had a highly significant effect (p≤0.01) on age at second birth; the model showed a coefficient of determination of 0.79, indicating that the effects included in the model explained the 79% variation of this characteristic. The Tukey analysis showed significant differences (p≤0.05) among the means, denoting the (negative/negative) genotype as the least favorable for age at second birth, with an average of more days. The analysis of means of the (positive/positive) and (negative/positive) genotypes showed no significant differences (p>0.05, Table 2).
Effect of allelic replacement of BGH gene polymorphism on the ages at which some reproductive events occur. For the age at first service, a regression coefficient (β) of -20.3 was found, indicating that for each (positive) allele that the individual bears, the age at first service decreases by 20 days; age at first birth had a regression coefficient (β) of -11.67, ie, for each (positive) allele, the age at first birth decreases by 11 days. For the age at first postpartum service the β coefficient was -8.5, indicating that for each (positive) allele, the age decreases by 8 days, and the β for the age at second birth was -15.83 indicating that for each (positive) allele, the age at second birth decreases by 15 days. Regression coefficients for each of the characteristics are shown in table 3.
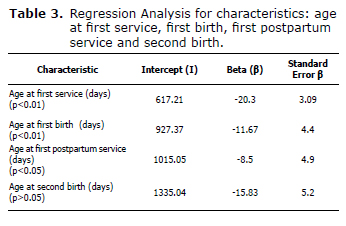
The estimated regression coefficients for age at first and second birth were highly significant (p<0.01). The regression coefficient for age at first birth was significant (p<0.05). For age at first postpartum service, the regression coefficient was not statistically significant (p>0.05).
DISCUSSION
Genotypic and allelic frequencies obtained in this study agree with those reported by Gorbani et al (15), who, in a population of 183 Holsteins, found frequencies of 0.787, 0.191 and 0.022 for (positive/positive), (positive/negative) and (negative/negative) genotypes, respectively, and 0.883 and 0.117 for (positive) and (negative) alleles respectively.
Mullen et al (16) found an association between three of six polymorphisms located in the first exon (5' region) of the GH gene. These newly found SNP were associated with pregnancy rate and first service; however, no association with the age at which the event occurs were performed. Reports generated by Katalin et al (17) mentioned that polymorphism recognized by the AluI enzyme in the exon 5 region position 127, and designated L and V alleles, is not associated with the age at first birth in Holstein cows.
The age at first breeding in heifers is an important parameter of reproductive efficiency; reducing the rearing period and rapid entry to the breeding program generates a higher economic return; a later age, the result obtained in this research, reflects low reproductive efficiency in the herd (1.2).
The age at first birth is another important factor in the productive life of the animal. Advanced ages at which this event occurs impact lesser milk production and increased feed intake, resulting, therefore, in a higher initial cost of breeding (18). Establishing the age around which heifers are giving birth evaluates the growth rate from birth to the time they contribute milk or calves to the system and return the rearing investment. The age at first birth in this study was also high and therefore should be the subject of work by both farmers and researchers.
The (negative/negative) genotype was less favorable, as it delays the age at first service, as well as the age at first birth, postpartum service and second birth. Although the low frequency of the (negative/negative) genotype may indicate some limitation when performing association analysis, the intron 3 polymorphism of the BGH showed significant results for all characteristics.
The greatest effect of genotype polymorphism of the BGH was on age at first service. For each (positive) allele, the first service occurred 20 days before. This effect decreased age at first birth by 11 days and for age at second birth by 15 days. The decrease in each of these parameters is important for the herd's breeding performance, resulting in a longer productive life (19, 20). Thus, association of BHG polymorphism can be used to help select animals that tend to enter reproductive stages earlier, decreasing economic losses generated in the breeding of animals to maturity (21).
The finding that the positive/positive genotype influences the age at first service, reducing the rearing period, may indicate a lower age at second birth and increased productive life of these carrier animals. Early entry to breeding programs generates greater economic performance and lower feed intake which results in lower breeding costs. This information, in productive and reproductive terms, is an important contribution to breeding programs (1).
In conclusion this is a pioneering study on the association of intron 3 polymorphisms of the BGH with reproductive characteristics of dairy cows. The results suggest that the intron 3 polymorphism of the BGH is associated with age at first service, first birth, first postpartum service and second birth by facilitating the selection of individuals with favorable genotypes for use in breeding programs.
We found that the positive/positive genotype is optimal for reducing age at first service, first birth, first postpartum service and second birth; the genotype's greatest effect being on age at first service.
Due to the low frequency of the negative/negative genotype; increasing the sample size is recommended for future studies and to achieve greater reliability in the results.
REFERENCES
1. Faure R, Morales C. La pubertad de la hembra bovina: I. aspectos fisiológicos. Rev Salud Anim 2003; 25(1):13-19. [ Links ]
2. Zhou G, Jin H, Liu C, Guo S, Zhu Q, Wu Y. Association of genetic polymorphism in GH gene with milk production traits in Beijing Holstein cows. J Biosci 2005; 30(5):595-598. [ Links ]
3. Abolfazl G, Rasoul T, Mortaza B, Cyrus A. Restriction fragment length polymorphism of bovine growth hormone gene intron 3 and its association with testis biometry traits in Iranian Holstein bull. Afr J Microbiol Res 2009;3(11):809-814. [ Links ]
4. Kanoth K, Kouno S, Okazaki A, Suzuki K, Obara Y. Interaction of GH polymorphism with body weight and endocrine functions in Japanese black calves. Domest Anim Endocrinol 2008;34(1):25-30. [ Links ]
5. Pereira A, Maurício M, Henrique N, Luciana C. Association of GH and IGF-1 polymorphisms with growth traits in a synthetic beef cattle breed. Genet Mol Biol 2005; 28(2):230-236. [ Links ]
6. Curi A, Oliveira N, Silveira C, Lopes R. Association between IGF-I, IGF-IR and GHRH gene polymorphisms and growth and carcass traits in beef cattle. Livest Prod Sci 2005; 94(3):159-167. [ Links ]
7. Kanoth K, Kouno S, Okazaki A, Suzuki K, Obara Y. Interaction of GH polymorphism with body weight and endocrine functions in Japanese black calves. Domest Anim Endocrinol 2008; 34(1):25-30. [ Links ]
8. An P, Hou J, Wang l, Li G, Wang J , Song Y. Polymorphisms of the growth hormone gene and their effect on growth traits in Chinese goats. Meat Sci 2010; 86(3):758-763. [ Links ]
9. Patton J, Kenny D, Mcnamara S, Mee J, O'mara F, Diskin M, et al. Relationships among milk production, energy balance, plasma analytes, and reproduction in Holstein-Friesian cows. J Dairy Sci 2007; 90(2):649-658. [ Links ]
10. Lucy M. Functional differences in the growth hormone and insulin-like growth factor axis in cattle and pigs: implications for post-partum nutrition and reproduction. Reprod Domest Anim 2008; 43(2):31-39. [ Links ]
11. Rodríguez N, López A, Echeverri J. Estructura genética poblacional del gen lactoferrina bovino en vacas Holstein del departamento de Antioquia. Rev MVZ Córdoba 2013; 18(1):3355-3361. [ Links ]
12. Dybus A. Associations of growth (GH) and Prolactin (PRL) genes polymorphisms with milk production traits in Polish Black and White cattle. Anim Sci P 2002; 20(4):203-212. [ Links ]
13. Hartl DL, and Clark AG. Principles of population genetics. 4th edition Sunderland (MA): Sinauer Associates Inc Publishers 2007. [ Links ]
14. SAS 9.2 SQL. Statistical Analysis Systems Institute., Inc., Cary, N.C, USA, 2009. [ Links ]
15. Gorbani A, Vaez T, Bonyadi M, Amirinia C. A MspI PCR-RFLP within bovin growth hormone gene and its association with sperm quality traits in Iranian Holstein bulls. Afr J Biotechnol 2009; 8(19);4811-4816. [ Links ]
16. Mullen P, Lynch C, Waters S, Howard D, O'boyle P, Kenny D, et al. Single nucleotide polymorphisms in the growth hormone and insulin-like growth factor-1 genes are associated with milk production, body condition score and fertility traits in dairy cows. Genet Mol Res 2011; 10(3):1819-1830. [ Links ]
17. Katalin K, József V, Attila Z, István G, László F. Associations between the AluI polymorphism of growth hormone gene and production and reproduction traits in a Hungarian Holstein-Friesian bull dam population. Arch Tierz 2006; 49(3):236-249. [ Links ]
18. Orrego J, Alfredo D, Echavarría L. Vida productiva y principales causas de descartes de vacas Holstein en la cuenca de lima. Rev int vet Perú 2003; 14(1):68-73. [ Links ]
19. Haworth G, Tranter W, Chuck J, Cheng Z, Wathes V. Relatioships between age at first calving and first lactation milk yield, and lifetime productivity and longevity in dairy cows. Vet Rec 2008; 162(20):643-647. [ Links ]
20. Marini P. Comportamiento de vacas de diferentes edades al primer parto. Arch Zootec 2004; 53(202):205-208. [ Links ]
21. Arauna P, Chakravarty A, Bhattacharya T, Joshi B, Arjava S. Detection of Polymorphism of Growth Hormone Gene for the Analysis of Relationship between Allele Type and Growth Traits in Karan Fries Cattle. J Anim Sci 2004; 17(10):1334-1337. [ Links ]
