











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Vertebrate craniofacial morphogenesis requires the interaction of the three germ layers (endoderm, ectoderm, and mesoderm) with the so-called fourth germ layer, the neural crest cell (NCC)-derived mesenchym. The latest begins with the differentiation of the neural crest at the boundary between the ectoderm and the neuroectoderm in the neural tube, from which NCCs from the cephalic region delaminate and migrate ventromedially to populate the branchial arches (BAs)1.
BAs are transitional segmented structures that form on the ventral region of the embryo and subsequently give rise to various structures of the face and neck, including the maxilla and the parathyroid glands 2. The first BA (BA1) is key in the development of the face, particularly during early stages of facial development, because it gives rise to the maxillary and mandibular prominences evident at late stages of facial development, which later form the maxilla and the mandible, respectively 3.
In parallel to the NCC migration toward the BAs, a fraction of these cells migrate anteriorly to form a fifth prominence called the frontonasal prominence, also called the frontonasal mass in chick embryos, which is responsible for nose, upper lip, and premaxilla formation 4. Several studies using mouse embryos have shown that the frontonasal prominence is composed of NCCs derived from the medial and anterior regions of the cephalic neural plate, while the BA1 is occupied by NCCs originating from the caudal-most region of the cephalic neural plate, which gives rise to the midbrain and the hindbrain 5.
Fusion of the five prominences, two maxillary, two mandibular (one at either side) and one frontonasal, gives rise to the mature form of the face. This is a highly conserved process in vertebrates, which makes its study possible in animal models, such as the chick embryo 6.
The complex configuration of the BAs and the facial prominences results from a strict combination of the expression domains of various molecules belonging to several cell signaling pathways 4. Previous studies have reported the involvement of signaling pathways, such as the Notch, Wnt, FGF, and BMP pathways, in the development of the precursor structures of the face through the regulation of cellular processes, such as proliferation, apoptosis, and differentiation 1,6.
The Notch signaling pathway is a mechanism of intercellular communication involved in various aspects of embryonic development as well as pathologies, such as CADASIL (cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy), cancer, and diabetes 7. Elements of this pathway include Notch receptors, Serrate and Delta family ligands, and target genes, such as Hes and Hey family genes 8.
The Hey gene family consists of three members, Hey1, Hey2 and HeyL, which are highly conserved between species 9. Hey1, a representative target gene of the Notch signaling pathway, encodes a transcription factor in the basic helix-loop-helix (bHLH) protein family. This transcription factor can bind to specific promoter sites to regulate the transcription of certain target genes, including Hey1 itself 9. The expression of this gene is linked to several events during embryogenesis that generally involve the regulation of cell fate and the formation of anatomical boundaries 10.
The Hey1 gene is involved in the embryonic development of various organs and systems, such as the heart, somites, central nervous system, and vascular system 11,12. Furthermore, this gene is a GGGnegative regulator of osteoblast mineralization in vitro and is expressed in craniofacial structures, such as the branchial clefts, BA1, nasal pit, otic vesicles, supporting cells of the auditory system, calvaria, maxilla, and mandible 10,13-15. Despite reports of Hey1 expression in structures such as the BAs, these descriptions are general and do not shed light about the specific distribution of Hey1 in the different germ layers. Thus, this work aimed to describe in detail the expression patterns of Hey1 in the precursor structures of face formation, the BAs and the facial prominences in chick embryos using in situ hybridization analysis.
MATERIALS AND METHODS
Chick embryo collection and fixation. Fertilized chicken (Gallus gallus) eggs obtained from a local egg farm were incubated at 37.5 -38.5ºC with 70% relative humidity until the embryos reached Hamilton-Hamburger stages HH14 through HH25 16. At least 3 replicates (n=3) were obtained for each stage.
Upon reaching an adequate stage, the embryos were extracted in PBS/DEPC (phosphate-buffered saline/diethylpyrocarbonate) to remove the extraembryonic membranes and were fixed in 4% paraformaldehyde (PFA) overnight at 4oC (6-12 hours). After fixation, the embryos were washed, dehydrated with PBT (PBS-Tween Buffer) and serial washes of methanol/PBT (25%, 50%, 75% and 100%) for 5 minutes each, and stored at -20ºC.
Riboprobe synthesis. Plasmids containing a DNA sequence encoding an antisense probe construct for chicken Hey1 were used for riboprobe synthesis and have been previously published 17. In vitro synthesis of the riboprobe was done with RNA polymerase T3 (Roche ® ) and digoxigenin UTP (DIG)-labeled ribonucleotides (Roche ® ).
Whole embryos ISH. In situ hybridization was performed using whole embryos following protocols used by the Developmental Biology group at Pompeu Fabra University-Barcelona Biomedical Research Park, with modifications as described below.
Embryos were rehydrated by serial washes in 75% to 25% methanol/PBT. Following one wash in PBT, the embryos were treated with 10 μg/ml of proteinase K for 3 to 15 minutes depending on the developmental stage of the embryo. Next, the embryos were washed in PBT and post-fixed in 4% PFA. Following post-fixation, the embryos were rinsed with PBT and hybridization buffer(SSC 5X,Formamide, EDTA, CHAPS 0.5%, Heparin and Yeast RNA). Hybridization was done overnight at 60ºC in hybridization buffer containing the probe. The embryos were then washed for 30 minutes with solutions I (formamide, SSC 20% and SDS 10%) and Solution III (formamide, 10% SDS) 3 times at 60ºC.
Subsequently, the embryos were washed in TBST and blocking solution to prepare them for anti-DIG addition and for DIG-labeled nucleotide visualization. After washing, anti-DIG was added to a fresh blocking solution in which the embryos were submerged overnight at 4ºC. The next day, the embryos were washed 3 times with TBST and incubated overnight (in TBST). On the fourth day, washes were done using TBST and NTMT. Visualization was done using BCIP/NBT solution for approximately 45 minutes while protected from light. Once visualization reaction was completed, the samples were washed with NTMT and PBT. The embryos were postfixed in 4% PFA and stored at 4ºC protected from light. Following ISH, the embryos were clarified by serial washing in methanol/PBT, PBT, and glycerol to be recorded. Images were obtained using a camera couple to stereoscope (Stemi 508 Zeiss).
Histological sectioning. To confirm the results observed in whole embryos, sagittal and coronal cryosectioning was performed using a thickness of 10 µm. The embryos used were those prepared for whole embryo in situ hybridization analysis. The embryos were over-developed, processed in different sucrose washes, and then soaked in OCT/sucrose for freezing. Frozen embryo sectioning was performed with a cryostat (Leica1850). The sections were mounted with 80% glycerol and were imaged in microscope Axio imager A2 (Zeiss).
Ethical aspects. The study samples were collected by the students of the Master in Dentistry at the Institute of Genetics of the National University of Colombia, which took into account the procedures for the collection of scientific, administrative and ethical samples contained in Law 84 of 1989 (National Congress of Colombia). The investigation was approved by the research committee of the Faculty of Dentistry of the National University of Colombia through act 03 of February 28, 2011. In addition, this research comprehensively covered the international principles of the 3 Rs y the Guide for the care and use of laboratory Animals (NIH Guide).
RESULTS
Hey1 expression during branchial arch development. Early stages of craniofacial morphogenesis. BA development in chick embryos occurs mainly between the Hamilton-Hamburger stages HH14 and HH1916.
In situ hybridization (ISH) analysis of whole embryos revealed that Hey1 gene expression was localized to the first three branchial clefts, the mesenchyme of the maxillary and mandibular prominence primordia, and the mesenchyme of BA2 during stage HH14 (Figure 1, A). Analysis of sagittal and coronal cryosections confirmed that Hey1 expression is especially present in the endoderm of the branchial pouches and the ectoderm that composes the branchial clefts, although at lower expression levels (Figure 1, B and C).
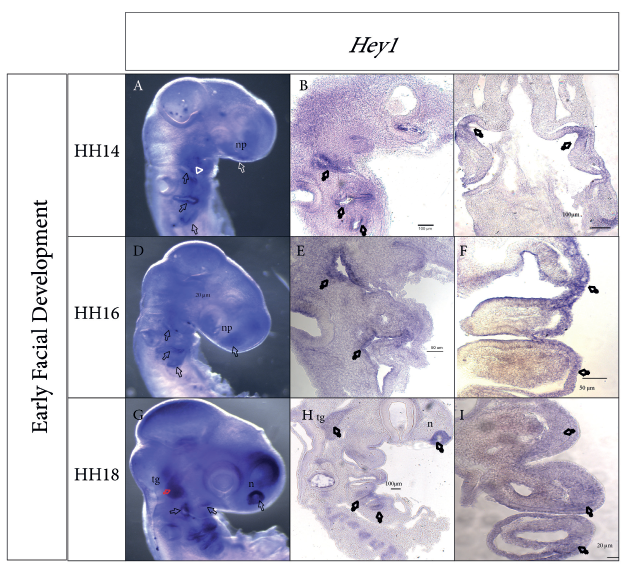
Figure 1 Hey1 gene expression patterns during branchial arch development. A, D, G Sagittal view of whole embryos during stages HH14, HH16, and HH18, respectively.B, E, H Sagittal cryosections. C, F, I Coronal cryosections. A, Expression in the three branchial clefts, first branchial arch, and nasal placode region. B, Expression in the ectoderm of the first, second, and third branchial clefts. C, Low expression in the branchial endoderm. D, Hey1 expression in the first three clefts and the nasal placode. E, Expression is maintained in the ectoderm of the clefts. F, Hey1 expression in the endoderm and mesenchyme of the first and second branchial arches. G, Expression is clearly visualized in the nasal pit, trigeminal ganglion region(red arrow), first three branchial clefts, maxillary and mandibular primordia, and second and third branchial arches. H, Sagittal cut confirms expression in the nasal epithelium, trigeminal ganglion region, ectoderm of the maxillary primordium, and ectodermic epithelium of the first and third branchial clefts. I, Coronal sections reveal Hey1 expression in the mesenchyme adjacent to the first and second branchial arches. f, frotonasal prominence; n, pit nasal; np, nasal placode; mx, maxillary prominence; md, mandibular prominence; tg, trigeminal ganglion.
During stage HH16, Hey1 gene expression was maintained in the first three branchial clefts, the maxillary prominence primordium, the dorsal side of the mandibular prominence primordium, the ventral region of BA2 and BA3, the margins of the otic vesicle, and in the nasal placode (Figure 1, D). Cryosection analysis confirmed its expression in the epithelium of the branchial clefts and pouches (Figure 1, E and F). Furthermore, Hey1 expression was detected in the BA mesenchyme (Figure 1, F).
During the HH18 stage, Hey1 expression exhibited an obvious, localized signal in the epithelium of the nasal pit (Figure 1, G and H). Expression was also detected in the anterior and posterior poles of the otic vesicle, the trigeminal ganglion, and in every cleft (Figure 1, G). Although in the dorsal region of the first cleft, Hey1 expression expanded toward the mesenchyme. The signal was also evident in the maxillary and mandibular prominences, although with lower intensity in the latter, in contrast to the signal observed in the ventral region of BA2 and BA3; furthermore, this signal extended from the second cleft toward the adjacent mesenchyme, invading the caudal-most region of BA2. Cryosection analysis confirmed extension of the expression toward the mesenchyme adjacent to the branchial clefts and pouches (Figure 1, H and I).
Hey1 expression during facial prominence development (late stages of craniofacial morphogenesis). Development of the facial prominences, which are precursors of the face, occurs mainly between the Hamilton-Hamburger stages HH21 and HH2516. During this phase, the prominences primarily undergo an increase in size and become morphologically established.
Analysis of stage HH21 showed that Hey1 expression remained localized to the epithelium of the nasal pit and maxillary and mandibular prominences and also the caudal-most region of BA2, trigeminal ganglion, and third and fourth branchial clefts, where a strong signal was observed (Figure 2, A). The signal, which was previously confined in the first branchial cleft, extended further toward the dorsal region of the cleft (Figure 2, A). Sagittal sections in the frontonasal mass region showed Hey1 expression in the nasal epithelium and the frontonasal mass ectoderm (Figure 2, C). Furthermore, coronal sectioning allowed detection of Hey1 expression in the epithelium, the lateral mesenchyme of the mandibular prominence, and the lateral epithelium of the maxillary prominence (Figure 2, B).
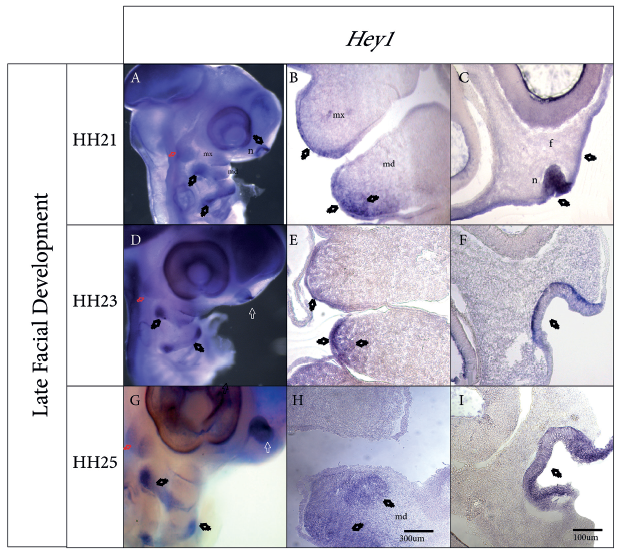
Figure 2 Hey1 gene expression patterns during facial prominence development. A, D, G Sagittal view of whole embryos during stages HH21, HH23, and HH25, respectively. B, E, H Coronal sections in the maxillary and mandibular prominence region. C, F, I Sagittal sections in the frontonasal prominence and nasal pit region. A, Expression is revealed in the nasal pit, trigeminal ganglion (red arrow), maxillary and mandibular prominences, and second branchial arch. B, Hey1 expression is observed in the mesenchyme adjacent to the epithelium of the mandibular prominence and the ectoderm that covers both prominences. C, Expression in the ectoderm that covers the frontonasal prominence and in the epithelium of the nasal pit. D, Expression in the nasal pit, trigeminal ganglion, maxillary prominence, mandibular prominence, and second branchial arch. E, Expression in the ectoderm that covers the maxillary and mandibular prominences, as well as the mandibular mesenchyme adjacent to the ectoderm. F, Clear expression in the epithelium of the nasal pit. G, Expression in the nasal pit, trigeminal ganglion, mandibular prominence, and second branchial arch. H, Coronal cut reveals Hey1 expression in the mandibular mesenchyme. I, Expression revealed in the epithelium of the nasal pit. f, frotonasal prominence; n, pit nasal; mx, maxillary prominence; md, mandibular prominence. Scale bar of 300 μm for B, E and H. Scale bar of 100 μm for C, F and I.
Subsequently, during the HH23 stage, Hey1 primary transcripts were detected in the nasal pit, the maxillary prominence, the ventral and dorsal regions of the mandibular prominence (more markedly on the dorsal side), and the region corresponding to the trigeminal ganglion (Figure 2, D and F). The analyzed sections revealed this expression to be in the epithelium and the lateral mesenchyme adjacent to the maxillary and mandibular prominences (Figure 2, E). Finally, the BA2 showed two expression domains, one ventral and the other dorsal, with a weaker signal in the first, third and fourth clefts (Figure 2, D and F).
In the latest stage analyzed, HH25, the expression patterns were similar to those observed for HH21 and HH23, although the signal was stronger. Namely, Hey1 expression was observed in the nasal pit and the dorsal region of the mandibular prominence (Figure 2, G). However, during this stage, Hey1 expression was not detected in the maxillary prominence. Sagittal cryosections showed that Hey1 expression was restricted to the epithelium of the nasal pit (Figure 2, I), while coronal sections in the mandibular prominence region showed expression specifically in the lateral mesenchyme of the mandibular prominence (Figure 2, H).
DISCUSSION
Various cell signaling pathways are associated with craniofacial morphogenesis1. The Notch signaling pathway target gene Hey1 is expressed in craniofacial structures, such as the BA1, suggesting a role for this pathway in face development10. Despite existing reports, the precise location of Hey1 gene expression in the germ layers that compose the BAs and the facial prominences, which are precursors of face development, is unclear. Using in situ hybridization analysis of chick embryos and histology sections, we present a detailed description of the expression patterns of this component of the Notch pathway.
Hey1 expression during branchial arch development. Early stage of face formation. The initial phase of craniofacial morphogenesis is characterized by the formation of the BAs, which arise during pharyngeal development when the lateral wall of the pharynx becomes invaginated, forming the structures known as the branchial pouches; in the outer embryo, the pharyngeal endoderm becomes depressed, forming the fissures known as the branchial clefts 2. The formation of these pouches represents the first sign of segmentation of the pharyngeal region, which is representative of all chordates and defines anterior-posterior polarity in the embryo2. Subsequently, the BAs increase in size due to the migration and proliferation of NCCs-derived cells, which form the BA mesenchyme along the paraxial mesoderm. Cells originating from each BA germ layer are responsible for the formation of a given specific structure or participate indirectly by signaling to the neighboring epithelium as a result of the mesenchyme-epithelium reciprocal interaction 18.
Using in situ hybridization analysis, it was found that between the HH14 and HH18 stages, Hey1 expression was primarily localized to the endoderm of the branchial pouches. Previous studies highlight the importance of the endoderm in BA organization not only through the formation of the branchial pouches as the first signal of pharyngeal segmentation but also as a source of signaling to the neighboring ectoderm and mesenchyme 19. Our findings in this study suggest that the Hey1 gene maybe involved in the initial segmentation of the pharyngeal region. In addition, the extension of Hey1 expression toward the mesenchyme may also suggest that a series of events occur at this stage, with the purpose of increasing cell proliferation to achieve an adequate arch size.
Hey1 expression during facial prominence development. Late stage of face formation. Establishment of the five facial prominences that form the face is the most important aspect of facial development. Facial prominences are mesenchyme buds covered by epithelium that emerge, grow and fuse to form the face 20. The maxillary and mandibular prominences arise from the growth and morphological differentiation of BA1. Meanwhile, the frontonasal prominence arises through the proliferation of NCCs originating from the anterior neural plate that gives rise to the forebrain and the midbrain5,6,21.
High levels of mesenchymal proliferation have been detected during the development of the maxillary and mandibular prominences between stages HH19 and HH21, mediated by epithelial-mesenchymal reciprocal interactions22. However, as development proceeds, these proliferation levels decrease, leading to cell differentiation processes that give rise to the structures derived from these prominences 22. In the present study, Hey1 gene expression was identified in the epithelium that envelops the maxillary and mandibular prominences during stages HH19 and HH21, which coincides with the highest levels of mesenchymal proliferation in the maxillary and mandibular prominences 22. This finding may suggest the involvement of the Notch pathway in this cellular process through the effector gene Hey1.
In addition, it was determined that between stages HH21 and HH26, mesenchymal cells condense and then differentiate into prechondrogenic cells that give rise to Meckel’s cartilage, which is the blueprint for mandibular development 23,24. Both this process of chondrogenesis and cell proliferation events depend on the interactions between the epithelium and the adjacent lateral mesenchyme 25. In this study, Hey1 gene expression was detected in the lateral mandibular mesenchyme between stages HH21 and HH25. These results agree with previous reports stating that chondrogenesis in the mandibular prominence begins during the same time period 23,24. Furthermore, other studies in mice have shown that Hey1 is required for the initiation of early chondrogenesis in human bone marrow cells and favors the formation of hypertrophic chondrocytes 14,26. Additionally, it has been shown that Hey1 activation through the Notch receptor favors the formation of chondrogenic cell condensates in human bone marrow cells 27. The expression patterns found in this study during the stage of mandibular chondrogenesis, specifically in the mandibular mesenchyme between stages HH21 and HH25, along with previous reports, suggest the involvement of the Notch pathway in the process of mandibular chondrogenesis via its effector gene Hey1.
Regarding the development of the frontonasal prominence, previous studies have shown that this process is regulated by a group of signals arising from two signaling nodes found in the nasal epithelium and the frontonasal ectodermal zone (FEZ)20,28,29. Several molecules, such as FGF8, BMP, and SHH, mediate proliferation and apoptosis in the frontonasal mesenchyme within these signaling nodes 20,29,30. In this study, Hey1 expression was detected in the nasal epithelium between stages HH19 and HH25. This expression pattern coincides with the point in time in which cell differentiation of the nasal epithelium occurs and with the stage at which the nasal epithelium signals for the development of the lateral nasal mesenchyme, suggesting the involvement of Hey1 in this process 20.
Hey1 transcripts were detected in the epithelium that covers the frontonasal prominence during stage HH21 (Figure 2, B). These expression patterns coincide with the stages in which the FEZ becomes established in birds, stages HH20-HH22 28. This signaling node is conserved between birds and mammals and is involved in controlling dorso-ventral polarity and the proximo-distal extension of the frontonasal prominence 28. The results obtained here suggest that Hey1 participates in the establishment of polarity of this prominence.
In conclusions this study revealed the specific expression patterns of the Hey1 gene during BAs and facial prominence establishment and development. These patterns of expression suggest a role for this component of the Notch pathway in craniofacial morphogenesis, possibly establishing pharyngeal segmentation patterns during early stages and/or regulating cell proliferation and differentiation during the late stages of face development. However, this role should be confirmed through studies exploring the correlation between gene expression patterns and cell proliferation patterns, such as gain and loss of function assays of Hey1.

