










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkUniversitas Scientiarum
versión impresa ISSN 0122-7483
Univ. Sci. v.13 n.2 Bogotá mayo/ago. 2008
In vitro regeneration of three chrysanthemum (Dendrathema grandiflora) varieties "via" organogenesis and somatic embryogenesis
Regeneración in vitro de tres variedades de crisantemo (Dendrathema grandiflora) a través de orgagénesis y embriogénesis somática
Luz Estela Monsalve3, Wilmer Acero3
Pontificia Universidad Javeriana, Cra. 7 No. 40-62, Bogotá, Colombia
2 Universidad de Pamplona, Pamplona, Colombia
3 Estudiantes pasantía, Unidad de Biotecnología Vegetal, Departamento de Biología,
Facultad de Ciencias, Pontificia Universidad Javeriana
ehodson@javeriana.edu.co, andrea.forero@javeriana.edu.co
Recibido: 12-03-2007: Aceptado: 14-10-2008:
Abstract
Chrysanthemum (Dendrathema grandiflora) has a high demand in the Colombian and international cut flower markets. Since commercial production of this ornamental species is strongly affected by fungal diseases such as chrysanthemum white rust (Puccinia horiana), high doses of fungicides are being used posing increased environmental and commercial costs. Assessment of in vitro regeneration systems from leaf discs was a first step in developing a plant genetic transformation protocol to obtain fungi-resistant plants. Leaf discs of White Albatross, Yellow Albatross, and Escapade varieties were established in vitro on MS medium supplemented with NAA (0 - 4.83 µM) and BAP (0 - 13.32 µM) alone and in combination. Leaf discs were also cultured on MumB medium containing 2,4-D (0 - 4.52 µM) for 7, 14, and 21 days prior to their transferral to a 2,4-D free MumB medium. Regenerated shoots were individualized, rooted, and hardened. Results show that MS with 4.83 µM NAA + 4.44 µM BAP and 4.83 µM NAA + 13.32 µM BAP induce organogenesis, and MumB with 2.26 µM 2,4-D induces somatic embryogenesis on all three varieties, with exposition periods to 2,4-D of 14 days for White Albatross and 21 days for Yellow Albatross and Escapade. Shoot development from somatic embryos was observed in the three varieties when cultured on a 2,4-D free MumB medium. Spontaneous rooting was recorded in 85% of the shoots thus facilitating hardening and successful transfer to soil.
Key words: Dendrathema grandiflora, in vitro regeneration, organogenesis, somatic embryogenesis.
Resumen
El crisantemo (Dendrathema grandiflora) presenta alta demanda en los mercados de flor cortada, tanto colombianos como internacionales. La producción de esta especie se ve seriamente afectada por enfermedades fúngicas como la roya Blanca (Puccinia horiana), lo que ocasiona que se empleen grandes cantidades de fungicidas aumentando los costos de producción a nivel económico y ecológico. La evaluación de sistemas de regeneración in vitro de crisantemo a partir de discos de hoja constituyó un primer paso hacia el empleo de la transformación genética, como apoyo a las técnicas de mejoramiento convencional para la obtención de plantas resistentes al hongo. Se establecieron discos de hoja de D. grandiflora var. Escapade, var. White albatross y var. Yellow albatross sobre medio MS en presencia ANA (0 - 4.83 mM) y BAP (0 - 13.32 mM) solos y en combinación. Así mismo, se establecieron discos foliares de las tres variedades en estudio sobre el medio MumB en presencia 2,4-D (0 - 4.52 mM) durante 7, 14 y 21 días, tiempo en el cual los explantes fueron transferidos a medio a medio Mum B sin 2-4D. Los brotes regenerados, fueron individualizados, enraizados y endurecidos. Los resultados obtenidos indican que el medio MS suplementado con: ANA 4.83 mM + BAP 4.44 mM ó ANA 4.83 mM y BAP 13.32 mM permite la regeneración de plantas vía organogénesis para las tres variedades y que es posible obtener embriones somáticos de las tres variedades, sobre medio Mum B en presencia de 2,4-D 2.26 mM, con periodos de incubación de 14 días para White Albatross y 21 días para Yellow Albatros y Escapade. El medio Mum B sin 2,4-D, permite el desarrollo de los brotes, a partir de embriones somáticos en los tres casos. El 85% de los brotes obtenidos presentaron enraizamiento espontáneo, lo que facilitó el endurecimiento y transferencia exitosa a suelo.
Palabras clave: Dendrathema grandiflora, regeneración in vitro, organogénesis, embriogénesis somática.
INTRODUCTION
Colombia is one of the largest flower producers in the world. With 35 years of activities in the flower industry, it is now ranked second exporter of fresh cut flowers after The Netherlands, having a total participation of 14% in the international commerce. Chrysanthemum (Dendrathema grandiflora) flower production is seriously affected by pathogenic fungi that are difficult to control by small growers due mainly to a lack tolerant plant material. As a consequence, fungal diseases are spread, thus increasing production costs and ecological impacts.
An approach to solve this problem is the introduction of resistance or tolerance traits via plant transformation. Studies on chrysanthemum genetic transformation aimed to obtain fungi-resistant plants have been conducted with around 200 varieties grown in Japan, Netherlands, Italy and United States (Texeira, 2003; Shirasawa et al., 2000; Kim et al., 1998, Rout & Das, 1997). In vitro regeneration of whole plants is a necessary step in the implementation of genetic transformation systems, therefore the assessment of different regeneration and micropropagation alternatives is required prior to any genetic transformation attempt. This study was aimed to find an appropriate system to regenerate plants via organogenesis and somatic embryogenesis from three chrysanthemum varieties: Escapade, White Albatross, and Yellow Albatross.
MATERIALS AND METHODS
Plant material
A constant supply of plant material for conducting all the experiments was obtained after having established and multiplied in vitro 450 nodal segments from 2-month old donor plants of D. grandiflora varieties Escapade, White Albatross, and Yellow Albatross, provided by the flower company Americaflor Ltd.
Surface disinfection was done placing the explants in a 1 g/l Benlate® solution for 20 minutes followed by immersion in a 1,25% sodium hypochlorite solution with 1% Tween 20® in constant agitation for 5 minutes. Disinfecting solution was rinsed three times with deionised distilled water during 1, 3, and 5 minutes respectively. Nodal segments were then established on Murashige & Skoog culture medium (MS) with full-strength salts, 3% sucrose, pH 5.8, and 0.6% agar, without growth regulators. Cultures were incubated at 21 ± 2°C with 16 hours of photoperiod provided by daylight 39W Sylvania® fluorescent tubes.
In vitro plant regeneration via organogenesis
Leaf segments of 0.5 cm2 were dissected from shoots developed under the culture conditions described before and cultivated using 16 different combinations of naphthaleneacetic acid (NAA) and 6-benzylaminopurine (BAP) to induce adventitious shoot formation. NAA concentrations were: 0, 1.61, 3.22, and 4.83 µM, and BAP concentrations were: 0, 4.44, 8.87, and 13.32 µM.
Leaf discs were placed on MS culture medium with fullstrength salts, 3% sucrose, pH 5.8, 0.6% agar and supplemented with growth regulators combined as described before. Cultures were incubated at 21 ± 2°C with 16 hours of photoperiod provided by daylight 39W Sylvania® fluorescent tubes during 60 days.
Data were recorded every 2 weeks for the following variables: shoot formation (BR), number of shoots produced per explant (Br/E), relative shoot regeneration rate (TRB) defined as Br2 – Br1/ t2 – t1 (number of shoots produced per day). The experimental unit was each leaf disc. Each treatment had 40 repetitions and the experiment was replicated three times. Shoots obtained were dissected and rooted on MS medium with full-strength salts, 3% sucrose, pH 5.8, 0.6% agar, without growth regulators.
In vitro plant regeneration via somatic embryogenesis
The effect of 2,4-dichlorophenoxyacetic acid (2,4-D) on somatic embryogenesis induction from leaf discs was evaluated after 7, 14 and 21 days of culture using the following concentrations: 0, 1.13, 2.26, 3.39, and 4.52 µM (Texeira, 2003; Tanaka et al., 2000).
The somatic embryogenesis induction protocol developed by Sherman et al. (1998) was followed based on the favourable results obtained with other chrysanthemum varieties. Leaf discs were cultivated on Mum B culture medium (MS medium supplemented with 2 mg/l indole-3- acetic acid (IAA) and 0.23 mg/l BAP. After 7, 14 and 21 days of culture, explants were transferred to Mum B culture medium free of 2, 4-D to allow embryo maturation.
Embryos showing signs of advanced development, even if still retaining masses of proembryogenic tissue (2-4 weeks depending upon the variety), were isolated from the initial explant and grown on a germination medium, which contained 2 mg/l kinetin (KIN), 0.02 mg/l NAA, and 10 mg/l gibberellic acid (GA3) on MS medium. After 4 weeks of culture, the number of plants obtained from each variety was determined. Explant incubation was carried out at 21 ± 2°C with 16 hours of photoperiod provided by daylight 39W Sylvania® fluorescent tubes during 60 days.
Every 2 weeks data were recorded for the following variables on induction of proembryonary masses: explant viability (V), presence of proembryonary masses (PEM), oxidation (OX), chlorosis (CL); and on embryo germination: shoot formation (BR), and number of plants obtained per explant (Br/E). The experimental unit was each leaf disc. Each treatment had 40 repetitions and the experiment was replicated three times.
Plant acclimatization
A sample of 40 plants of each variety was obtained out of the total plants regenerated in vitro via somatic embryogenesis and organogenesis. Plants were transferred to a sealed chamber and grown on peat substrate. Relative humidity was gradually reduced inside the chamber to allow progressive acclimatization of plants.
Data analysis
Anovas and Duncan comparison tests for each evaluated factor were carried out using the Statistical Analysis System package (SAS) 8.0 for each of the three varieties studied.
RESULTS AND DISCUSSION
In chrysanthemum, as in many other species, there is a clear relationship between varieties and successful regeneration. Studies on adventitious shoot regeneration from petal explants, nodal segments; protoplasts and somatic embryos report a wide range of responses depending upon varieties and types of explant (Shirasawa et al., 2000; Kim et al., 1998; Lowe et al., 1993). The regeneration capacity of several varieties of chrysanthemum, in order to select the genotype with the highest response, as well as the plant material on which the subsequent genetic transformation experiments were going to be based on, has been evaluated by several researchers (Texeira, 2003; Sherman et al., 1998). Specific requirements of varieties in achieving in vitro regeneration are of paramount relevance when additional manipulations that usually reduce the tissue morphogenetic capacity are required, such as inoculating Agrobacterium strains or the use of antibiotics.
In vitro plant regeneration via organogenesis
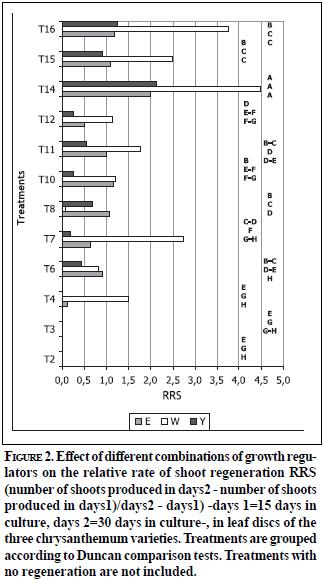
Regeneration via organogenesis was achieved on the three varieties. Presence of BAP in the culture medium was necessary for shoot regeneration, although concentrations higher than 4.44 µM reduced the shoot regeneration frequency. This inhibitory effect has been reported with other chrysanthemum varieties, and it has been related to a negative effect of BAP on protein synthesis (Staden & Crounch, 1996). Additionally, the requirement of an exogenous stimulus with auxins (NAA) to obtain a successful regeneration was evident, since shoot formation in absence of this plant regulator (T1-T4) was very low or was not observed (Figure 1). These results support previous findings in other varieties, where a given BAP/NAA balance was required to obtain adventitious shoot induction (Nhut et al., 2003; Texeira, 2003). Despite several studies having reported the requirement of a high cytokinin/auxin ratio (Chakrabarty et al., 2000), the present study reports higher organogenic responses when cytokinin/auxin ratios are near to 1.0 (Figures 1 and 2). This result can be related to manipulations of exogenous cytokinin concentrations that may cause an increase of endogenous auxin concentrations, which is probably induced by an inhibition of free IAA conjugation due to the presence of exogenous cytokinins (Haberer & Kieber, 2002). A proportional stimulus provided by plant growth regulators allows the cell cycle regulation required to induce cell division and specialization during plant development initiated from isolated cells of a differentiated tissue, such as leaf discs. Similar previous studies show that auxins control endogenous levels of active cytokinins by inhibiting their synthesis or promoting the N-glucoside formation or the cytokinin-oxidase activation (Coenen & Lomas, 1997). Chaudhury & Rongda (2000) have shown that requirements of exogenous needed to produce an organogenic response depend upon specific concentrations of endogenous regulators. Several physiological effects of cytokinins can be explained by their biochemical interaction with auxins, considering that both regulators are mutually dependent. The manipulation of cytokinin levels causes an increase of levels of endogenous auxins thus stimulating undifferentiated tissue production and favouring root formation on explants and further rooting on a hormonefree medium (Haberer & Kieber, 2002).
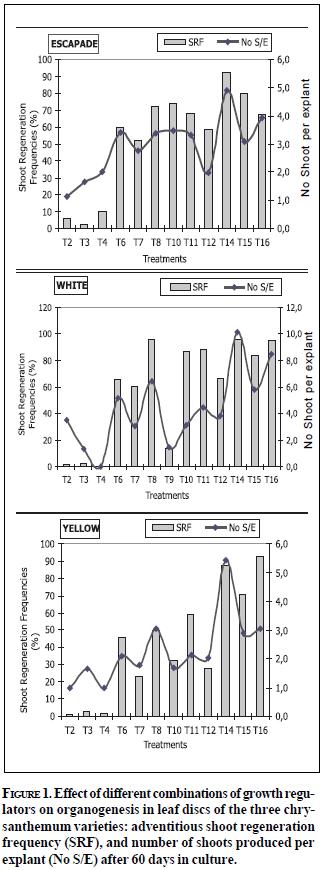
The highest shoot production was observed on the explants cut edge as a consequence of mechanical injury caused on tissues, which provides an effective stimulus in inducing cell competence needed to regenerate adventitious organs (Figure 3). Response to tissue damage is the biological basis of somatic cell proliferation and regeneration as injuries on the explants cut edge provoke a rapid cell division and eventually provide an adequate target tissue for further manipulations such as genetic transformation. High numbers of shoots were also developed along the leaf mid rib, a common phenomenon reported with other species.

Carnier & Carneiro (1994) observed in Passiflora that the high organogenic potential of this leaf area is due to a high concentration of plant growth regulators present in its tissues.
Presence of undifferentiated tissue was a common feature to all the three varieties regardless the treatment employed. This response has been also reported with other chrysanthemum varieties where BAP has stimulated callus production (Texeira, 2003; Rout et al., 1997; Khan et al., 1994). However, there was a significant percentage of explants with no callus or shoot regeneration, especially in Yellow Albatross variety (Table 1). Inhibition of regeneration in some species has been explained by excessive accumulation of polyamines (putrescine diamine and spermidine) resulting from stress provoked during in vitro tissue culture (Bais & Ravishankar, 2002). Metabolism and polyamines effect are strongly influenced by other plant growth regulators and the application of hormones (auxins, cytokinins and gibberellins) generally increase the polyamines content. This response has been described with some varieties of chrysanthemum (Martín et al., 1997) and could therefore be correlated to the low shoot production on Yellow Albatross leaf explants.
Explant oxidation affected the induction of organogenesis, particularly with Yellow Albatross and Escapade varieties. Phenol production by leaf tissue is induced by the activation of a chain of enzymatic reactions due to hypersensitivity to stress factors such as desiccation or mechanical wounding, which are frequent during the excision of leaf discs from in vitro material, and to pH alterations and changes in osmotic and water potentials occurring during the explant incubation stage. These reactions are initiated when cell membranes are injured, which increases respiration rates. Lipoxigenase enzyme is then activated promoting the production of free radicals of oxygen that provoke a higher activity of the peroxidase enzyme. Peroxidase catalyzes the production of H2O2 (an oxidizing agent causing tissue damage) and polymerization of monophenols, which produce dehydrogenated polymers such as condensed tannins, ending with the typical browning of in vitro tissues that have undergone the phenolization process (Goodman & Novacky, 1994). Oxidation products are phytotoxic and therefore alter the morphogenetic process.
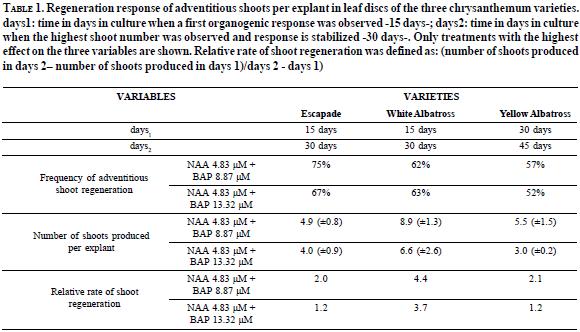
In relation to shoot individualization, rooting and acclimatization, shoot elongation was observed in all the shoots obtained, and rooting occurred in most of them without any exogenous growth regulator in the culture medium. High rooting percentages were obtained with all the three varieties (92% with Escapade, 91% with White Albatross and 87% with Yellow Albatross) as opposed to studies with other chrysanthemum varieties, which reported the requirement of gibberellic acid for shoot elongation and IAA or NAA for rooting induction (Texeira, 2003; Tanaka et al., 2000; Sherman et al., 1998). Acclimatization of regenerated whole plants from all the three varieties was successfully achieved, facilitating transfer to soil (Escapade 97%, White Albatross 100%, and Yellow Albatross 97%).
Plant regeneration from leaf discs via somatic embryogenesis
Leaf tissues of the varieties employed in this study exhibit a high capacity of response to phytoregulation stimuli that facilitate competence acquisition, determinism and induction of embryo formation under the culture conditions evaluated (Figure 3). A favourable effect of 2, 4-D on the induction of somatic embryogenesis was evident, similarly to previous reports on chrysanthemum and other species (Fiore et al., 2002).
Significant differences were found between treatments evaluated with Escapade (E) and Yellow Albatross (Y) varieties. There was a favourable effect of 2.26 µM 2, 4-D after 21 days of culture of leaf discs, on proembrionic structure formation (E: 76. 4% and Y: 72.03%), unipolar germination (E: 51,14% and Y: 5,3%) and average number of plants produced per explant (E: 2 and Y: 1). White Albatross variety also developed proembrionic masses with 2.26 µM 2, 4-D although a reduced time of exposure (14 days) was more favourable for further shoot development -5,7% germination frequency and 1.4 regenerated plants per leaf disc- (Figure 4).
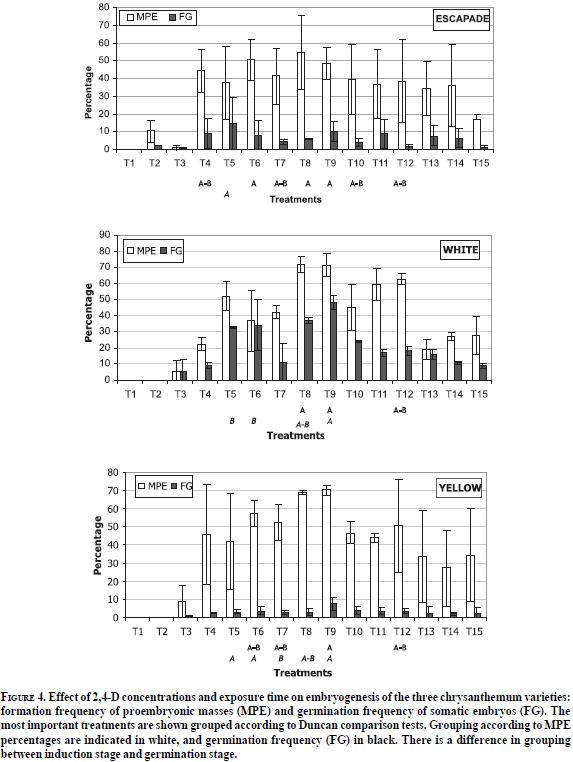
Results obtained in the present study can be explained by three different reasons. Firstly, initial high concentrations of 2, 4-D may cause a stress condition giving way to DNA methylation, which has been correlated to low levels of genetic expression and the interruption of the initial expression pattern resulting in the activation of the embryogenic programme. Although the induction of somatic embryogenesis activates genetic control pathways similar to those of zygotic embryogenesis, which can be regarded as a universal pattern in plants, the variable response can be accounted by genotypic differences responsible of a differential activation of the embryo development pathway (Freire, 2004). Secondly, the combined application of low concentrations of cytokinin (BAP) with high concentrations of auxins (IAA + 2, 4-D) in the culture medium Mum B favoured the embryogenic response, since BAP increases tissue sensitivity to auxin stimulus thus facilitating cell dedifferentiation and subsequent differentiation required to shift from a somatic cell state to an embryogenic one. Finally, the central role played by endogenous levels of IAA during the early stages of embryogenesis must be considered. Addition of exogenous auxins as 2, 4-D has proved to substantially increase the endogenous concentrations of IAA, which are in turn associated to high embryogenic responses in several plant species (Feher et al., 2003). Studies have concluded that when 2, 4-D is present in the tissues, phosphatidilinositol (Ptdlns) hydrolysis is stimulated, cytoplasm and cell wall pHs are altered, and asymmetrical cell divisions are repeated as a result of altered cell polarity, leading to the formation of globular embryos.
Germination frequencies obtained with each treatment were significantly different. A 21-day exposure period favoured the formation of proembrionic structures (76,4%) and germination (51%) of Escapade leaf discs, in contrast with Yellow and White Albatross varieties, in which low germination frequencies (Y: 5.3% and W: 5,7%) and reduced numbers of plants obtained per explant (Y: 1 and W: 1.4) were recorded (Figure 4). Low germination frequencies can be due to an overexposure to 2, 4-D, considering that transfer of embryogenic calli to a 2, 4-D-free medium was necessary to facilitate embryo germination, as reported by Sherman et al. (1998). Shoot and root elongation did not require any exogenous hormone supplement as was evident from the regeneration of whole plants achieved after transferring embryos of the three varieties to MS basal medium.
The development sequence of somatic embryos was determined with the three studied varieties. Initial formation of proembrionic masses was observed 7-10 days after having established the leaf disc cultures on Mum B medium (stage 0). Proembryos shifted to a globular stage 14-21 days after sowing (stage 1) and, with a reduction of auxin concentrations, began to develop into heartshaped structures (stage 2, bilateral symmetry and cotyledon formation). The torpedo structure was reached in 22-30 days. Torpedo embryos initiated a precocious unipolar germination process, ending with early shoot development and late root formation (Figure 3). Germination during this morphogenesis stage is undesirable because embryos are unable to store enough compounds necessary to enter into a quiescent state, which is a fundamental requisite to achieve a synchronic development (Feher et al., 2003). Additionally, the opaque white coloration expected to be present during the stage of embryo maturation was not easily identified and this may be an indication of an anomalous accumulation of storage compounds inside the embryo delaying the development of the apical root meristem and subsequent bipolar germination (Datta et al., 2001).
Embryogenic processes are affected by a series of factors that in certain circumstances can either favour or difficult the in vitro manipulation; these factors include: the genotype, culture conditions, growth regulators and other medium components, and the type and physiological state of the explant (Fiore, et al., 2002; Bornhoff & Harst, 2000). Alterations of the maturation and germination processes are believed to be a consequence of an initial overexposure to 2, 4-D and its subsequent effect on the polyamine interconversion process. When initial concentrations of auxins are low or absent in the culture medium, the synthesis of spermine and spermidine polyamines from diamine putrescine is triggered. These reactions are regarded as basic to cell differentiation needed during the process of embryo maturation that can be affected by an auxin overstimulus. Additionally, the presence of ABA, the osmotic manipulation of the culture medium, or an aminoacid supplement may be required to achieve appropriate maturation and germination of somatic embryos of the three varieties.
Several authors have reported that the addition of ABA into the culture medium regulates embryo maturation, prevents precocious germination and facilitates the accumulation of carbohydrates and storage proteins. Besides ABA, osmotic agents such as polyethylene glycol (PEG), mannitol and sucrose have been demonstrated to inhibit precocious germination, the establishment of initial high osmotic pressures in the culture medium being more effective than an exogenous provision of ABA (Freire, 2004). Aminoacid incorporation into the culture medium in the form of organic nitrogen, hydrolyzed casein and glutamine, amongst others, showed beneficial effects on the maturation of avocado somatic embryos. Further experiments on the effect of ABA and aminoacids on controlling the precocious germination of chrysanthemum somatic embryos are required to be conducted. The assessment of exposure periods to 2, 4-D less than 14 days is also required. Future basic research on somatic embryogenesis of ornamental plants can be promising in helping to reduce production costs of laborious micropropagation and hand labour.
CONCLUSIONS
Reproducible and efficient regeneration systems —via organogenesis and somatic embryogenesis— from three chrysanthemum (D. grandiflora) varieties: Escapade, White Albatross, and Yellow Albatross were obtained. The morphogenetic capacity of the three varieties of D. grandiflora was shown. Acclimatization and transfer to soil of regenerated whole plants from all the three varieties was successfully achieved.
Regeneration via organogenesis was achieved in the presence of BAP in the culture medium. The highest shoot production was observed on the explants cut edge and along the leaf mid rib as a consequence of mechanical injury caused on tissues, which provides an effective stimulus in inducing cell competence needed to regenerate adventitious organs. Plant regeneration from leaf discs via somatic embryogenesis was also obtained in the study, observing a favourable effect of 2, 4-D on the induction of somatic embryogenesis.
The development of the regeneration systems allows genetic transformation studies in chrysanthemum (D. grandiflora)
ACKNOWLEDGMENTS
Financial support was provided by Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología 'Francisco José de Caldas' and Pontificia Universidad Javeriana (Colciencias Project 1203-12-123-86). Americaflor Ltda. provided the plant material required to carry out this study. Santiago Grillo-Astudillo (Universidad Nacional de Colombia) was of invaluable help in assisting with the statistical analyses and Sandra Constantino in translating the manuscript into English.
REFERENCES
BAIS, H.P. and RAVISHANKAR, G.A. Role of polyamines in the ontogeny of plants and their biotechnological applications. Plant Cell and Organ Culture, 2002, 69, 1-34. [ Links ]
CARNIER, M. and CARNEIRO, M. L. Tissue culture studies on species of Passiflora. Plant Cell Tissue and Organ Culture, 1994, 36, 211-217. [ Links ]
CHAKRABARTY, D.; MANDAL, A.K. and DATTA, S.K. SEM and light microscopic studies on direct shoot regeneration from ray florets of Chrysanthemum. Israel Journal of Plant Sciences, 2000, 48, 105-107. [ Links ]
CHAUDHURY, A. and RONGDA, QU. Somatic embryogenesis and plant regeneration of turf-type Bermuda grass: effect of 6-benzyadenine in callus induction medium. Plant Cell Tissue and Organ Culture, 2000, 60 (2), 113-120. [ Links ]
COENEN, C. and LOMAX, T.L. Auxin-citoquinin interactions in higher plants: olds problems and new tools. Trends Plant Science, 1997, 2, 351-356. [ Links ]
DATTA, K.B.; KANJILAL, B. and DE SARKER, D. Artificial seed technology: Development of a protocol in Geodorum densiflorum (Lam) Schltr. An endangered orchid. Current Science, 2001, 76 (8), 1142-1145. [ Links ]
FEHER, A.; PASTERNACK, T.P. and DUDITS, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tissue and Organ Culture, 2003, 74, 201-228. [ Links ]
FIORE, S.; DE PASQUALE, F.; CARIMI, F. and SAJEVA, M. Effect of 2, 4-D and 4-CPPU on somatic embryogenesis from stigma and style transverse thin cell layers of citrus. Plant Cell Tissue and Organ Culture, 2002, 68, 57-63. [ Links ]
FREIRE, S.M. Aspectos básicos de la embriogénesis somática. Revista del Ministerio de Educación Superior (MES) de la República de Cuba. Biotecnología Vegetal, 2004, 3 (4), 195-211. [ Links ]
GOODMAN, R.N. and NOVACKY, A.J. The hypersensitive reaction in plants to pathogens: a resistance phenomenon. The American Phytopathological Society, APS Press. St. Paul., 1994. 244 p. [ Links ]
HABERER, G. and KIEBER, J. Cytokinins. New Insights into a classic Phytohormone. Plant Physiology, 2002, 128, 354-362. [ Links ]
KIM, J.; PARK, Y.; JUNG, S.; CHUNG, H.; SHIN, S. and SHEOP, J. Transformation of chrysanthemum by Agrobacterium tumefaciens with three vectors. Journal Korea Society for Horticultural Science, 1998, 39, 360–6. [ Links ]
LOWE, J.M.; DAVEY, M.R.; POWER, J.B. and BLUNDY, K.S. A study of some factors affecting Agrobacteriumtransformation and plant regeneration of Dendranthema grandiflora Tzvelev (syn Chrysanthemum morifolium Ramat.). Plant Cell Tissue and Organ Culture, 1993, 33, 171-180. [ Links ]
MARTIN, T.J.; ARIBAUD, M.; CARRE, M. and GASPAR, T. ODCmediated biosyntheis and DAO-mediated catabolism of putrescine involved in rooting of chrysanthemum explants in vitro. Plant Physiology and Biochemistry, 1997, 35, 595-602. [ Links ]
NHUT, D.T.; TEIXEIRA, D.A.; SILVA, J.A. and ASWATH, C.R. The importance of the explant on regeneration in thin cell layer technology. In Vitro Cellular and Developmental Biology, 2003, 11, 266-276. [ Links ]
ROUT, G.R. and DAS, P. Recent trends in the biotechnology of Chrysanthemum: a critical review. Scientific Horticulture, 1997, 69, 239-256. [ Links ]
ROUT, G.R.; PALAI, S.K.; PANDEY, P. and DAS, P. Direct plant regeneration of Chrysanthemum morifolium Ramat Deep Pink: influence of explant source, culture environment, carbohydrates, nutritional factors and hormone regime. Proceedings National Academy of Science India, Sect B Biology Science, 1997, 67, 57-66. [ Links ]
SHERMAN, J.; MOYER, J. and DAUB, M. A Regeneration and Agrobacterium-mediated transformacion system for genetically diverse Chrysanthemum cultivars. Journal of the American Society for Horticultural Science, 1998, 123, 189-194. [ Links ]
SHIRASAWA, N.; IWAI, T.; NAKAMURA, S. and HONKURA, R. Transformation and transgene expression of chrysanthemum (Dendranthema grandiflora (Ramat.) Kitamura). Bulletin of Miyagi Prefecture Agriculture Research Center, 2000, 67, 15-20. [ Links ]
STADEN, J. and CROUNCH, N. Benzyladenine and derivatestheir significance and interconversion in plants. Plant Growth Regulation, 1996, 19, 153-175. [ Links ]
TANAKA, K.; KANNO, Y.; KUDO, S. and SUZUKI, M. Somatic embryogenesis and plant regeneration in Chrysanthemum (Dendranthema grandiflorum (Ramat) Kitamura. Plant Cell Report 19: 2000, 946-953. [ Links ]
TEIXEIRA, J.A. Carbon Source affects the outcome in vitro organogenesis of Chrysanthemum thin cell layers. New Zealand Journal of Crop and Horticultural Science, 2003. [ Links ]

