










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkUniversitas Scientiarum
versão impressa ISSN 0122-7483
Univ. Sci. v.14 n.1 Bogotá jan./abr. 2009
Efecto del 3-nitropropionato sobre el metabolismo del lactato y del acetato en neuronas y astrocitos crecidos in vitro en concentraciones perinatales
3-Nitropropionate effect on the lactate and acetate metabolism of neurons and astrocytes grown in vitro with perinatal concentrations
Efeito do 3-nitropropionato sobre o metabolismo do lactato e do acetato em neuronios e astrocitos crescidos in vitro em concentrações perinatales
2Departamento de Biología, Universidad del Tolima, Ibagué, Tolima
Recibido: 08-04-2008; Aceptado: 05-06-2009
Resumen
Las reacciones anapleróticas son un mecanismo metabólico esencial para la continuidad postnatal del desarrollo cerebral, contribuyendo en procesos que requieren sustratos sintetizados a partir de intermediarios del ciclo de Krebs; sin embargo, se desconoce su papel en el neonato durante la prelactancia. Objetivo. Estimar la capacidad anaplerótica de neuronas y astrocitos crecidos in vitro. Materiales y métodos. Se midió el efecto del 3-nitropropionato (3-NPA)(2 mM), un inhibidor de Succinato Deshidrogenasa (SDH) sobre el metabolismo oxidativo y lipogénico de 14C derivados de acetato y lactato en concentraciones perinatales. Resultados. A pesar de la presencia del 3-NPA, se mantuvieron las velocidades de oxidación del lactato en neuronas y astrocitos en un 40 y 73% respectivamente y la lipogénesis en un 53 y 52% respectivamente. Con el acetato, la oxidación en neuronas y astrocitos se mantuvo en un 15 y 63% respectivamente, en tanto que la lipogénesis se mantuvo en astrocitos y aumentó en neuronas en un 174% (p<0,05). Todo esto comparado con sus respectivos controles sin inhibidor. Conclusiones. Estos resultados demuestran que a pesar de la depleción de oxalacetato generada por 3-NPA, las neuronas como los astrocitos son capaces de mantener en la prelactancia el metabolismo energético y la síntesis de lípidos empleando lactato o acetato gracias a la actividad anaplerótica. Adicionalmente, los astrocitos demostraron tener mayor capacidad de amortiguar los efectos del 3-NPA sobre la oxidación que las neuronas. Las neuronas y los astrocitos demostraron una mayor capacidad de dirigir el acetato para la síntesis de lípidos, activando la vía Acetil-CoA Sintetasa citosólica.
Palabras clave: acetato, astrocito, neurona, 3-nitropropionato, lactato.
Abstract
Anaplerotic reactions are an essential metabolic mechanism for the postnatal continuity of the brain development, contributing in processes that require substrates synthesized from Krebs cycle intermediates; however, their role during the presuckling period in the neonate is unknown. Objective. To estimate the anaplerotic capacity of neurons and astrocytes grown in vitro under perinatal conditions. Materials and methods. The effect of 3-nitropropionate (3-NPA)(2 mM) an inhibitor of the succinate dehydrogenase (SDH) on the oxidative and lipogenic metabolism of 14C-derived from acetate and lactate in perinatal concentrations. The results were compared with its respective controls without inhibitor. Results. In spite of the presence of 3-NPA, respiratory activity with lactate was 40% in neurons and 73% in astrocytes, the lipogenesis was 53% in neurons and 52% in astrocytes. With acetate, the oxidation in neurons was 15% and 63% in astrocytes, lipogenesis was maintained in astrocytes but in neurons it increased up to 174% (p<0.05). Conclusions. These results demonstrate that in spite the oxalacetate depletion generated by 3-NPA, neurons as well as astrocytes are able to maintain the energetic metabolism and the lipid synthesis using lactate or acetate thanks to the anaplerotic activity in the presuckling period. Additionally, astrocytes showed a capacity of buffering the effects of 3-NPA on the oxidation process greater than the neuron capacity. Neurons and astrocytes revealed a better capacity of directing acetate for lipid synthesis, activating the cytosolic acetyl-CoA synthetase pathway.
Key words: acetate, astrocyte, neuron, 3-nitropropionate, lactate.
Resumo
As reações anapleróticas são um mecanismo metabólico essencial para a continuidade pos-natal do desenvolvimento cerebral, contribuindo nos processos que precisam substratos sintetizados a partir de intermediários do ciclo de Krebs. Embora, seu papel é desconhecido no neonato durante a prelactancia. Objetivo. Estimar a capacidade anaplerótica de neuronios e astrocitos crescidos in vitro. Materiais e métodos. Quantificou-se o efeito do 3-nitropropionato (3-NPA)(2 mM), um inibidor do Succinato Deshidrogenasa (SDH), sobre o metabolismo oxidativo e lipogénico de 14C derivados de acetato e lactato em concentrações perinatais. Resultados. A pesar da presença do 3-NPA, mantiveram-se as velocidades de oxidação do lactato em neuronas e astrocitos num 40 e 73% respectivamente e a lipogenesis em 53 e 52% respectivamente. Com o acetato, a oxidação em neuronas e astrocitos mantive-se num 15 a 63% respectivamente, no em tanto, a lipogénesis mantive-se em astrocitos e aumento em neuronas num 174% (p<0,05). Tudo o anterior comparado com seus respectivos controles sem inibidor. Conclusões. Estes resultados demonstram que a pesar da depleção do oxalacetato gerada pelo 3-NPA, as neuronas como os astrocitos são capazes de manter na prelactancia o metabolismo energético e a síntese de lípidos empregando lactato ou acetato devido à atividade anaplerótica. Adicionalmente, os astrocitos demonstraram ter maior capacidade de amortecer os efeitos do 3-NPA sobre a oxidação que as neuronas. As neuronas e os astrocitos apresentaram uma maior capacidade de dirigir o acetato para a síntese de lipídeos, ativando a via Acetil-CoA sintetasa citosólica.
Palavras-Chave: acetato, astrocito, neurona, 3-nitropropionato, lactato.
Introducción
La prelactancia constituye un período crítico para el neonato en la transición de la vida intrauterina a la extrauterina ya que debe subsistir a expensas de los sustratos acumulados durante los últimos días de la gestación, cuando paralelamente los sistemas enzimáticos que se encargan de la oxidación de cuerpos cetónicos y lactato presentan su máximo pico de actividad (1), las concentraciones de glucosa plasmática disminuyen inmediatamente después del nacimiento presentándose una hipoglucemia. Dado que hay altos requerimientos energéticos en este período y al no ser suficiente para mantener el metabolismo energético del cerebro, se hace necesario que este órgano emplee sustratos energéticos alternativos, puesto que la etapa de desarrollo postnatal en rata y el humano requiere de un continuo suministro de carbonos no sólo para obtener energía sino también para construir estructuras cerebrales. Estos sustratos alternativos como los cuerpos cetónicos y el lactato incrementan sus concentraciones durante este período, garantizando la homeostasis energética y por tanto la continuidad de procesos vitales como el desarrollo de las sinapsis, crecimiento de axones, dendritas y la mielinización activa de vías neurales básicas.
Se ha demostrado que el cerebro en el período perinatal es capaz de utilizar el lactato y el acetato, calculándose la capacidad máxima de estos sustratos como precursores oxidativos y lipogénicos (2-5). Estos trabajos han expuesto que debe existir una actividad anaplerótica importante para el mantenimiento de la homeostasis energética de neuronas y astrocitos. Por lo tanto, se ha propuesto que una inhibición del ciclo de Krebs permitiría estimar la contribución de las reacciones anapleróticas en condiciones perinatales. Por esta razón, se ha determinado el efecto de un inhibidor de la succinato deshidrogenasa (SDH), el 3-nitropropionato (3-NPA) sobre las velocidades de oxidaci ón y lipogénesis del lactato y del acetato. El efecto neto del 3-NPA es inhibir el ciclo de Krebs, induciendo una disminución de la concentración de oxalacetato (OAA), favoreciendo a que el ciclo sea mantenido por activación de las reacciones anapleróticas de las cuales las más importantes son las catalizadas por la malato deshidrogenasa-NADP dependiente citosólica (cME) y la piruvato carboxilasa (PC) exclusivas de astrocitos; malato deshidrogenasa-NADP dependiente mitocondrial (mME) neuronal y astrocítica (6). Este trabajo ha demostrado que tanto las neuronas como los astrocitos en condiciones perinatales son capaces de mantener el metabolismo energético y la síntesis de lípidos a pesar de la depleción de OAA, generada por la presencia del inhibidor de la SDH cuando se emplean sustratos como lactato o acetato, corroborando la hipótesis de una actividad importante de las reacciones anapleróticas para el mantenimiento de la homeostasis cerebral durante la prelactancia.
Materiales y métodos
Cultivos primarios de neuronas fueron preparados a partir de cerebros de fetos de 17,5 días y los de astrocitos a partir de cerebros de neonatos de 1 día de ratas albinas Wistar (3, 7, 8). Las células fueron sembradas en frascos Roux, con una densidad de 1,35 x 106 células/ml para el cultivo de neuronas y 9 x 106 células/ml para el cultivo de astrocitos. Las células fueron mantenidas en medio Eagle modificado por Dulbecco (DMEM), al cual se le adicionó bicarbonato de sodio anhidro (3,7 g/l) y cloruro de potasio (1,86 g/l)(316 mOsm/kg H2O) y fue suplementado con suero fetal bovino (FBS) (10%), ampicilina, estreptomicina, anfotericina y penicilina a 37°C en incubadora con 5% de CO2. Una vez las células formaron una capa confluente y quiescente (a 7 días para las neuronas y 13 días para los astrocitos), fueron utilizadas para los experimentos y para hacer el contaje celular (9, 10).
Composición del medio Elliott de incubación
Esta solución tiene una composición aproximada a la del fluido cerebro espinal. Tiene más baja concentración de calcio que el medio Krebs y Henseleit, pero aproximadamente la misma cantidad de calcio ionizado en el plasma. El calcio tiende a precipitar con el fosfato; para asegurarse que no precipita, se adiciona una solución que tenga como máximo una concentración de 11 mM en fosfato de sodio. La composición definitiva fue: NaCl (122 mM), KCl (4,8 mM), KH2PO4 (0,4 mM), MgSO4 (1,2 mM), CaCl2 (1,3 mM), preparado en tampón fosfato sódico 10.8 mM a pH=7.6. Extemporáneamente, el pH se ajustó a 7,4, se filtró (0,22 µm poro) y se gaseó con oxígeno durante dos horas (292,4 mOsm/kg H2O).
Preparación de las soluciones para las incubaciones
Se ha elegido, de modo general, concentraciones saturantes de cada uno de los sustratos fríos según resultados previos obtenidos en el laboratorio (3, 4), L-lactato (10,5 mM) y acetato sódico (5 mM). Se hicieron determinaciones cuantitativas de cada uno de los sustratos fríos en el medio previamente a la incubación así como, para calcular la actividad específica (dpm/nmol). Se tuvo en cuenta las concentraciones para ajustar la osmolaridad a valores cercanos a 315 mOsm/kg H2O.
A 1,5 ml de medio de incubación Elliott, con los respectivos sustratos fríos por frasco Roux, se adicionaron, de acuerdo al experimento, los trazadores radiactivos [U-14C]-lactato (1 µCi) y [U-14C]-acetato, [1-14C]-acetato y [2-14C]-acetato (1 µCi). De cada solución se tomaron 100 µL para determinar la radiactividad inicial necesaria para calcular la actividad específica (dpm/nmol).
Incubación con cultivos primarios de neuronas y astrocitos
Se emplearon neuronas de 7 días y astrocitos de 13 días en cultivo, crecidos en frascos tipo Roux. Después de retirar el medio de cultivo se lavaron los frascos 2 veces con PBS y se añadió el medio de incubación, con los sustratos fríos y radiactivos deseados. Seguidamente se gasearon los frascos con O2 durante 30 segundos, se cerraron herméticamente con un tapón de goma y se incubaron a 37°C. En paralelo se llevaron frascos sin células pero con medio de incubación y sustratos fríos y radiactivos. Estos frascos sirvieron de blanco. Para finalizar la incubación los frascos se enfriaron a 4°C (3, 4).
Cuantificación de CO2
Para capturar el 14CO2 producido durante la incubación se empleó el método descrito por Sykes, con algunas modificaciones (11, 12). Para capturar el CO2 se utilizó un matraz erlermeyer con un pocillo central que sostenía un eppendorf con 500 µl de hidróxido de hiamina [cloruro-p-(diisobutilcreoxietoxietil) dimetilbencil amonio], que se utiliza para atrapar el CO2 en los experimentos de respiración. La hiamina emite quimioluminiscencia de baja energía por lo que se hizo curva de apantallamiento para hacer las correcciones a los datos finales. En el pocillo principal se adicionó 100 µl de KOH (10 M) después de lo cual se selló herméticamente con un tapón de goma.
Sin destapar el frasco de cultivo, se extrajo con una jeringuilla el medio de incubación y se inyectó en el correspondiente matraz erlenmeyer previamente preparado. Se lavó la monocapa de células con PBS que, posteriormente, fue recuperado en su respectivo matraz. Se inyectaron 2 ml de KOH (0,3 M), colocando el frasco de manera que el KOH no contacte las células pero capture el CO2 remanente en el frasco de cultivo. Una hora después, el KOH fue retirado e inyectado en su respectivo matraz erlenmeyer. Se realizó otro lavado con PBS y, por último, una vez reunidos todos los volúmenes se adicionó 100 µl de HClO4 (5 M) en el matraz Erlenmeyer con objeto de acidificar el medio y volatilizar el 14CO2 que luego es capturado por la hiamina, este proceso dura 1 hora.
A continuación se recogieron todos los tubos eppendorf que contienen el hidróxido de hiamina, se colocaron en viales y se les adicionó 5 ml de líquido de centelleo, se agitaron por 30 segundos y se dejaron en reposo 16 horas para medir las desintegraciones por minuto (dpm) utilizando la técnica de espectroscopia de centelleo líquido. La velocidad de respiración se reporta como nmol de CO2 producidos por hora y por millón de células.
Cuantificación de lípidos
Para hacer la determinación de la incorporación de los sustratos en lípidos totales, se siguió el método recomendado para el aislamiento y purificación de lípidos con ligeras modificaciones (3). Para extraer los lípidos, la monocapa de células se separó del frasco de cultivo con ayuda de un raspador en 1 ml de metanol, y se añadió a un tubo que contiene 2 ml de cloroformo (bidestilado). El tubo se agitó por 30 segundos y se almacenó durante 16 horas en el congelador. Posteriormente se centrifugó (1500 x g, 15 min, 4°C) y se lavó con NaCl (0,3%) saturado con cloroformo. Se realiz ó una centrifugación en las condiciones anteriores, se retiró la fase acuosa y se recogió la fase clorofórmica. Esta última fase se evaporó y el residuo lipídico se disolvió en líquido de centelleo, se agitó mecánicamente por 30 segundos y después de 24 horas, se midió la radiactividad incorporada en lípidos utilizando la técnica de espectroscopia de centelleo líquido. La velocidad de lipogénesis se reporta como nmol de carbonos incorporados a lípidos por hora por millón de células.
Cálculo de la velocidad de utilización de sustratos
La velocidad de utilización de los sustratos se calcula dividiendo la radiactividad incorporada en CO2 o lípidos, por la radiactividad específica de los sustratos en el medio de incubación (dpm/nmol), por el tiempo de incubación (1 h) y por el número de células (x 106) (3, 4). A las dpm de oxidación se les resta el valor del blanco. De acuerdo con este cálculo, los resultados se expresan como nmol de sustrato radiactivo transformados en CO2 o en lípidos, por hora y por 106 células.
Tratamiento estadístico
Los ensayos se condujeron según un diseño completamente aleatorizado, con triplicado, como mínimo, de cada determinación. El análisis de la significatividad entre las réplicas de un mismo experimento se ha efectuado utilizando el test "t" de Student. Los resultados obtenidos fueron evaluados estadísticamente usando el programa para Macintosh, STAT.VIEW. Se hicieron comparaciones múltiples utilizando el análisis de varianza (ANOVA), para determinar los efectos de los diferentes factores considerados, este análisis incluye el test F de significatividad. Las comparaciones que presentaron diferencias significativas entre tratamientos se expresaron con letras diferentes. Las letras mayúsculas se utilizaron para las neuronas y las letras minúsculas para astrocitos. Los resultados se expresan como promedios ± la desviación estándar de la media (SEM).
Resultados y discusión
Diferencias metabólicas en la utilización del lactato y del acetato
La utilización de estos dos monocarboxilatos obedece a su disponibilidad en el medio (concentración); a su afinidad por transportadores; al acople con enzimas cuya actividad o expresión depende del estado de desarrollo celular; al compartimento (citosol o mitocondria) donde están expresadas las enzimas, además de diversas vías reguladoras y/o moduladoras. Adicionalmente, también puede depender de la cantidad y madurez mitocondrial.
En los astrocitos, el piruvato uniformemente marcado ([U-14C]-piruvato) proveniente del lactato uniformemente marcado ([U-14C]-lactato) vía lactato deshidrogenasa (LDH) entra al ciclo de Krebs en la primera vuelta vía complejo piruvato deshidrogenasa (CPDH) generándose 14CO2 y [U-14C]-acetil-CoA, o vía PC originando [1,2,3-14C]-OAA; el citrato puede ser sintetizado desde [U-14C]-acetil-CoA y OAA frío o [1,2,3-14C]-OAA, así, existe la posibilidad de que se produzcan dos isotopómeros del isocitrato, pero sólo a través de la oxidación del [2,3,4,5-14C]-isocitrato y [2,3-14C]-isocitrato se puede generar 14CO2 desde el primer turno vía Isocitrato deshidrogenasa. La oxidación del α-cetoglutarato (α-KG) no produce 14CO2; más adelante, la hidrólisis del enlace tioéster de la succinil-CoA por la succinato tioquinasa produce succinato, una molécula simétrica.
Consecuentemente, el reservorio mitocondrial de malato presenta todos los carbonos marcados; así, en la reacción de la cME y fosfoenolpiruvato carboxiquinasa (PEPCK) (más activa en neuronas que astrocitos en la prelactancia) se generaría nuevamente 14CO2. Ya en la segunda vuelta a través del ciclo de Krebs se produce [1,2,3-14C]-OAA, que junto con [U-14C]-acetil-CoA y acetil-CoA frío, generan [U-14C]-isocitrato y [1,2,3-14C]-isocitrato, desde los cuales se puede producir 14CO2 vía isocitrato deshidrogenasa y posteriormente a través de los sustratos originados desde el a-cetoglutarato (α-KG) uniformemente marcado α-[U-14C]-KG) y α-[1,2,3-14C]-KG vía complejo α-cetoglutarato deshidrogenasa (α-CKGDH), nuevamente el reservorio de malato presenta todos los carbonos marcados.
En neuronas, el [U-14C]-piruvato proveniente del [U-14C]-lactato, entra al ciclo de Krebs en la primera vuelta vía CPDH y mME, a través de la cual se genera [1,2,3-14C]-malato y posteriormente [1,2,3-14C]-OAA, de igual manera se produce [2,3,4,5-14C]-isocitrato y [2,3-14C]-isocitrato como en astrocitos, sin embargo, no en la misma proporción, puesto que la actividad anaplerótica de la PC no existe en estas células.
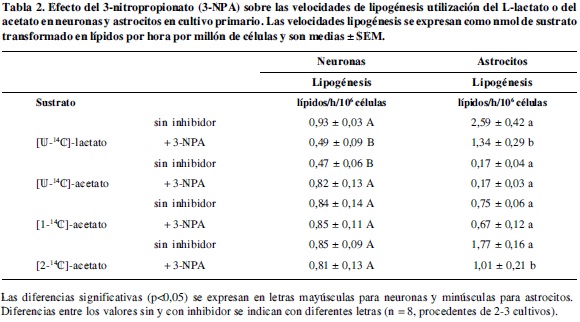
Las velocidades de oxidación y lipogénesis medidas en condiciones de control (Tablas 1 y 2), indican que in vitro, tanto las neuronas como los astrocitos son capaces de metabolizar lactato y acetato, confirmando inicialmente que estos sustratos utilizaron mecanismos de transporte al interior celular.
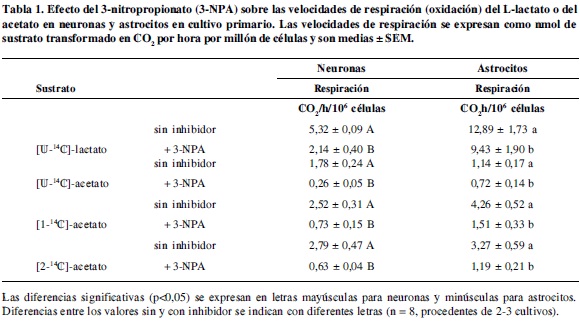
De acuerdo con los resultados, en primer lugar se puede decir que ambos tipos de células metabolizan en mayor proporción el lactato como sustrato oxidativo y lipogénico, lo cual en parte se puede deber a la menor concentración del acetato (5 mM), y en segundo lugar el acetato fue preferiblemente metabolizado por las neuronas, en tanto que el lactato por los astrocitos, probablemente por las siguientes razones:
El lactato extracelular es captado vía transportador de monocarboxilatos (MCT); las neuronas expresan la isoforma MCT-2 en tanto que los astrocitos las isoformas MCT-1 y MCT2, lo cual podría favorecer una mayor velocidad de transporte (13). Adicionalmente, en cuanto a la enzima relacionada con el metabolismo del lactato, dado que en este período es el sustrato energético y lipogénico más importante, es posible que la isoforma predominante sea la lactato deshidrogenasa tipo 1 (LDH-1) tanto en neuronas como astrocitos, favoreciendo su oxidación ya que durante el período perinatal los requerimientos energéticos son elevados.
Es posible que la actividad de la enzima LDH-1 sea mayor en astrocitos y de esta manera favorece una mayor disponibilidad de piruvato para ser oxidado o incorporado en lípidos; sin embargo, se requiere conocer con mayor precisión las diferencias en el nivel de actividad y patrón isoenzimático de la LDH expresado por neuronas fetales y astrocitos de neonatos crecidos in vitro, dado que a diferencia de la prelactancia, en la adultez se conoce bien que las neuronas expresan LDH-1 y los astrocitos principalmente LDH-5 que favorece la producción de lactato que es exportado (14). Por otra parte la actividad de la PC incrementa por el aumento de las concentraciones de piruvato (15), por lo tanto, los astrocitos pueden incorporar piruvato generado desde lactato en el ciclo de Krebs en una mayor proporción que las neuronas, dado que la capacidad anaplerótica de éstas se encuentra restringida a la mME cuya actividad es menor (16).
Por otro lado, se ha observado que el [U-13C]-lactato o [U-14C]-lactato se incorpora más en aminoácidos en neuronas que astrocitos, los cuales pueden ser el glutamato, aspartato, alanina, N-acetil-L-aspartato (NAA) y el neuromodulador N-acetil-L-aspartil-L-glutamato (NAAG) (14, 15) estos dos últimos se sintetizan en altas proporciones durante el desarrollo, constituyendo rutas importantes de escapes del marcaje que no se incorporaría en 14CO2 o lípidos, y por lo tanto, no cuantificables por la metodología empleada en el presente trabajo, lo cual podría explicar en parte el menor metabolismo oxidativo del lactato en neuronas a favor de la síntesis de neurotransmisores, mientras que en los astrocitos el lactato sería utilizado principalmente para la oxidación y en menor proporción para síntesis de lípidos como el ácido oleico o aminoácidos como glutamina.
En cuanto al metabolismo del acetato, este es un monocarboxilato que entra en ambos tipos de células vía MCT-2; sin embargo, en trabajos previos empleando el ácianocinnamato, un inhibidor de transportador de monocarboxilatos (3), se ha encontrado que el metabolismo oxidativo y lipogénico de neuronas no se altera, en tanto que el de los astrocitos disminuye (4), esto ha llevado a sugerir la participación de un mecanismo de transporte adicional posiblemente más activo en neuronas que podría ser el sistema intercambiador de aniones (mAE2) ya reportado, por lo tanto, la expresión de dos formas alternativas de transporte durante la prelactancia en neuronas favorecer ía la mayor velocidad de entrada de este sustrato.
Por otro lado, el metabolismo preferencial del acetato por neuronas se puede explicar por el nivel de actividad de las enzimas relacionadas con su metabolismo. En trabajos previos se ha demostrado que el acetato en condiciones perinatales, puede ser utilizado por neuronas y astrocitos por acción de la acetil-CoA sintetasa (AceCS) sugiriendo la actividad de dos isoformas una mitocondrial (AceCS2) y otra citosólica (AceCS1) en ambos tipos de células (3-5, 16). El mayor metabolismo oxidativo y lipogénico neuronal pueden estar indicando que la acetil-CoA sintetasa tipo 2 (AceCS2) parece ser mucho más activa en este tipo de células que en astrocitos favoreciendo la disponibilidad de Acetil-CoA mitocondrial para la síntesis de citrato. El hecho que las neuronas sinteticen altas cantidades de NAA y NAAG (18) puede indicar la presencia de un reservorio de Acetil-CoA mitocondrial más alto que en astrocitos y por tanto indica la mayor actividad de la AceCS2 neuronal. El nivel de actividad de la AceCS1 puede también estar contribuyendo a la lipogénesis neuronal donde su actividad puede ser mayor que en astrocitos.
Capacidad anaplerótica en la utilización del lactato y del acetato
Para determinar la contribución y la importancia de las reacciones anapleróticas en el mantenimiento de la homeostasis energética durante el período perinatal se utiliz ó un inhibidor de la succinato deshidrogenasa, el 3-nitropropionato, a través de la evaluación de su efecto sobre el metabolismo del lactato y del acetato en neuronas y astrocitos crecidos in vitro en condiciones perinatales. Con este fin, se utilizó una concentración 2 mM de 3-NPA de manera que no se indujera un bloqueo total del ciclo de Krebs en los dos tipos de células y se viera un efecto que reflejara su capacidad anaplerótica. Esta concentración no afecta la viabilidad celular dado que concentraciones de 3 mM y 10 mM bloquean completamente el ciclo de Krebs en cultivos de neuronas y astrocitos respectivamente (19).
Los resultados obtenidos (Tabla 1) muestran que se presentó una disminución significativa en las velocidades de oxidación del lactato en presencia del 3-NPA. Los resultados con este sustrato demuestran que las neuronas fueron mucho más sensibles al inhibidor que los astrocitos en cuanto a que se disminuyó la producción de CO2 en un 59%, mientras que en los astrocitos se disminuyó en un 26%. Lo anterior evidencia que un bloqueo parcial del ciclo de Krebs en ambos tipos de células que puede alterar la homeostasis energética en la prelactancia. Sin embargo, el hecho que las neuronas hayan sido capaces de mantener la respiración desde lactato en un 41% y los astrocitos en un 74%, es un reflejo de la actividad anaplerótica de estas células, y se evidencia que es mayor en astrocitos como ya se ha reportado (20).
Se ha discutido que la actividad anaplerótica de las neuronas vía mME es muy importante para la síntesis de glutamato y su actividad carboxilante o descarboxilante depende de los requerimientos celulares de ATP o neurotransmisores, lo cual se debe reflejar en los niveles de malato o piruvato; es así como una carboxilación neta del piruvato a malato puede ocurrir durante la activación neuronal, cuando se requiere ATP extra para restaurar los potenciales de membrana y cuando α-KG es necesario para restituir el reservorio de glutamato (21). Sin embargo, aun no hay consenso en cuanto a lo anterior, pues también se ha sugerido (15) que las neuronas no tienen una actividad anaplerótica cuantitativamente importante como para mantener los niveles de los intermediarios del ciclo de Krebs; nótese que en este artículo (15) se trabajó en concentraciones de lactato de cerebro adulto (1 mM) y a una concentración de 3-NPA (3 mM) que inhibe completamente el ciclo de Krebs en neuronas. Teniendo en cuenta lo anterior, se puede decir que la concentración de 3-NPA y de lactato, influyen en la actividad de la mME en la carboxilación del piruvato en las neuronas permitiendo la continuidad del ciclo de Krebs, lo cual no sería posible cuando la síntesis de OAA está bloqueada. Adicionalmente, en experimentos de fijación de NaH14CO3 en cultivos primarios de neuronas se ha reportado que hay incorporación de 14C en glutamato, aspartato y prolina, adicionalmente, la incorporación de 13C desde NaH13CO3 en aspartato, glutamato, GABA y glutamina (22), lo que demuestra que las neuronas tienen activa la ruta de la mME.
Los resultados del presente trabajo con [U-14C]-lactato sugieren que los astrocitos presentan una mejor capacidad de amortiguar los efectos del 3-NPA, en la concentración empleada, con relación a las neuronas, probablemente por el efecto anaplerótico de la PC, también relacionado con la baja actividad ciclo de Krebs (5-10% menor que en las neuronas) y la inhabilidad del 3-NPA de causar un incremento de calcio (23). Esta resistencia también se corrobora a nivel del metabolismo energético, como se demuestra por los resultados obtenidos en el presente trabajo, donde las velocidades de oxidación fueron mayores en astrocitos que en neuronas en presencia del inhibidor, lo cual no ha sido reportado.
La lipogénesis a partir del lactato disminuyó en neuronas y astrocitos en un 47 y 48% respectivamente, lo cual se explica porque decrece la generación de citrato mitocondrial a partir del [U-14C]-piruvato como consecuencia de la disminución de oxalacetato y el incremento dramático de los niveles de succinil-CoA que disminuyen la actividad de la citrato sintetasa. Por estas dos razones, se esperaba que la lipogénesis se viera afectada.
No obstante, la lipogénesis se mantiene a pesar de los efectos del inhibidor en ambos tipos de células, puede ser debida a la activación de la ruta citosólica desde lactato, reportado en trabajos previos empleando un inhibidor del transporte de citrato mitocondrial, el 1,2,3-bencenotricarboxilato, donde se encontró que las neuronas incrementaban su lipogénesis (p<0,05), mientras que en los astrocitos no se observó variación en la velocidad de incorporación del sustrato a lípidos (2); esto permitió postular que debía existir una ruta alternativa para la síntesis de lípidos, que debía ser muy activa en el período perinatal.
Estos resultados son coherentes con las conclusiones obtenidas a partir de trabajos en neuronas y astrocitos para evaluar el metabolismo del lactato (10,5 mM), empleando aminooxiacetato (AOA) (5 mM), un inhibidor de la aspartato aminotransferasa que incrementa las concentraciones de OAA intramitocondrial, que a diferencia de lo observado con el 3-NPA, la oxidación y lipogénesis se incrementó significativamente en neuronas y en astrocitos, concluyendo que la lanzadera del citrato sería la principal fuente de acetil-CoA en el citosol para la síntesis de lípidos a través de la ATP-citrato liasa (4).
En cuanto al metabolismo oxidativo del acetato, el 3-NPA causó una disminución de 85% en las neuronas y de un 36% en los astrocitos debido al descenso en la velocidad del ciclo de Krebs en ambos tipos de células. Sin embargo, a diferencia del lactato, el acetato no es precursor de OAA, por lo tanto, al disminuir las concentraciones de este intermediario ambos tipos de células deben recurrir a sustratos endógenos. En este sentido, dado que los astrocitos tienen la capacidad de almacenar lactato y glucógeno y una mayor capacidad anaplerótica, les permite mantener la oxidaci ón a partir de acetato en un 64%. El mantenimiento de la oxidación en neuronas tan sólo un 15% sugiere que las concentraciones de OAA son más limitantes en ellas y consecuentemente la utilización de acetil-CoA en mitocondria disminuye en la vía de síntesis de citrato, beneficiando la ruta de síntesis de NAA y NAAG que constituyen una ruta de escape de carbonos que pudieron ser utilizados en el ciclo de Krebs o para la síntesis de lípidos.
En cuanto a la lipogénesis (Tabla 2), se puede decir que fue activa en ambos tipos de células a pesar de la disminución de la utilización de acetato en la mitocondria para la síntesis de citrato. Sin embargo, el hecho de que no se afecte en astrocitos y por el contrario ocurriera un aumento de la lipogénesis en neuronas, está sugiriendo que la AceCS1 en éstas es más activa cuando los niveles de citrato citosólico no son suficientes corroborando los resultados obtenidos empleando a-ciano-4-cinnamato (α-CN) en trabajos previos (3-5).
Otras investigaciones con aminoxiacetato (OAA) y butilmalonato (BM), han demostrado que la lipogénesis y oxidación en ambos tipos de células aumenta, dado que el efecto de estos inhibidores es el incremento de las concentraciones de OAA mitocondrial. Al comparar estos resultados con los obtenidos con 3-NPA, el cual presenta efecto contrario, se comprueba que la variación en las concentraciones de este intermediario regula el ciclo de Krebs en ambos tipos de células (3-5). Una disminución mayor en la velocidad de oxidación en neuronas en presencia del inhibidor puede obedecer al escape de acetato para la síntesis de glutamato, aspartato, alanina, y especialmente NAA, NAAG, que se generan en altas concentraciones durante el período perinatal.
Nuevamente los resultados con acetato y 3-NPA sugieren que los astrocitos presentan una mayor capacidad de tolerar los efectos del inhibidor ya sea porque presenta mayor capacidad anaplerótica o porque almacenan sustratos precursores de OAA.
Metabolismo diferencial del acetato
El carbono 1 (1-14C) de los tres sustratos empleados [U-14C]-acetato, [1-14C]-acetato y el [2-14C]-acetato puede transformarse en CO2 rápidamente en la primera vuelta del ciclo de Krebs a través de la cME en astrocitos o mME en neuronas y a través del CPDH al inicio de la segunda vuelta del ciclo de Krebs. Comparativamente, el carbono 2 (2-14C) se descarboxila en la segunda vuelta a través del ciclo de Krebs por las mismas vías y por efecto de la isocitrato deshidrogenasa y el complejo a-cetoglutarato deshidrogenasa (α-CKGDH), indicando que puede permanecer más tiempo en el ciclo de Krebs, con mayor probabilidad de incorporarse en metabolitos sintetizados a partir de intermediarios como citrato o α-KG. Lo anterior implica que podrían obtenerse velocidades de oxidación menores que las observadas con [1-14C]-acetato.
Por otro lado, el [U-14C]-acetato se incorpora menos en 14CO2, que los anteriores sustratos ya que proporcionalmente se pueden escapar más carbonos para la síntesis de aminoácidos (glutamato, glutamina y aspartato) o lípidos durante la primera vuelta a través del ciclo de Krebs (3, 4).
El [1-14C]-acetato, fue un 40% más oxidativo en astrocitos que en neuronas, lo que probablemente está reflejando el efecto de la cME, PC y PEPCK, más activas en este tipo de células, así aportando una mayor proporción de descarboxilaciones. La menor velocidad de oxidación del [2-14C]-acetato en astrocitos refleja la fuga de carbonos en el ciclo de Krebs principalmente para la síntesis de citrato que serviría para síntesis de lípidos o glutamina en el citosol, constituyendo rutas de escape del marcaje, lo que no se observa en neuronas, donde las velocidades de oxidación de ambos sustratos ([1-14C]-acetato y [2-14C]-acetato) permanecen iguales, indicando que el acetato en neuronas permanece por más tiempo en el ciclo de Krebs porque es un sustrato más oxidativo que en los astrocitos, coincidiendo con la mayor velocidad de oxidación del [U-14C]-acetato en neuronas (3-5).
En el presente estudio, los astrocitos y neuronas en cultivo en condiciones perinatales y en presencia del 3-NPA disminuyeron significativamente las velocidades de oxidación de los sustratos [1-14C]-acetato y [2-14C]-acetato como consecuencia de la disminución de la velocidad del ciclo de Krebs; nuevamente las neuronas resultaron significativamente más afectadas (Tabla 1).
En los controles, la incorporación del [1-14C]-acetato en lípidos es similar en ambos tipos de células, mientras que el [2-14C]-acetato es incorporado significativamente en lípidos por astrocitos que en neuronas, dado que en éstas últimas, la lipogénesis permanece igual que con [1-14C]-acetato. Lo anterior indica que los astrocitos desvían más citrato para síntesis de lípidos que las neuronas, por lo que el [2-14C]-acetato puede marcar más citrato que [1-14C]-acetato. De cualquier manera la síntesis de lípidos aportado por la AceCS1 es relevante cuando los niveles de citrato citosólico no son apropiados (Tabla 2).
A pesar de la presencia del 3-NPA, la lipogénesis en neuronas se mantiene con [1-14C]-acetato y [2-14C]-acetato, indicando que el descenso en los niveles de OAA, que afectan la síntesis de citrato no influyen en la síntesis de lípidos neuronal que puede estar sustentada por la AceCS1; mientras que en astrocitos se mantiene con [1-14C]-acetato, pero la disminución observada con [2-14C]-acetato puede significar la dependencia de formación de citrato mitocondrial para la síntesis de lípidos que de igual manera puede estar dada por la AceCS1. Estos resultados confirman una mejor actividad de la AceCS1 en las neuronas que en los astrocitos. Adicionalmente, señalan una mayor dependencia, en los astrocitos de actividad anaplerótica para el mantenimiento del citrato mitocondrial, cuya disminución afectara la síntesis de lípidos, en especial el ácido oleico o la exportación de citrato, indicando que durante el período perinatal una alteración de estos dos mecanismos tendría un efecto negativo en el metabolismo y modulación de la señalización neuronal.
Los resultados en su conjunto indican que en condiciones de ayuno (prelactancia), las neuronas requieren y emplean todos los recursos energéticos posibles para mantener su metabolismo, incrementado por los requerimientos de ATP para la síntesis de neurotransmisores y el mantenimiento de la bomba Na+/K+. Lo anterior está de acuerdo con el hecho de que el ciclo de Krebs neuronal funciona principalmente para el metabolismo oxidativo y en astrocitos para la síntesis de intermediarios que son exportados hacia las neuronas.
Conclusiones
Las neuronas y los astrocitos en condiciones perinatales utilizan preferentemente el lactato que el acetato como sustrato oxidativo y lipogénico, no obstante, el acetato demostró ser más lipogénico en las neuronas, mientras que el lactato lo es para los astrocitos. Los resultados indican una mayor actividad de las Acetil-CoA Sintetasas neuronales y de la Lactato Deshidrogenasa astrocítica. Tanto las neuronas como los astrocitos son capaces de mantener en la prelactancia, el metabolismo energético y la síntesis de lípidos a pesar de la depleción de oxalacetato (OAA) generada por la presencia del inhibidor de la succinato deshidrogenasa (SDH) cuando se emplean sustratos como lactato o acetato; lo cual puede ser atribuido a enzimas anapleróticas cuantitativamente más importantes en el cerebro, la piruvato carboxilasa y las isoformas citosólica y mitocondrial de la malato deshidrogenasa NADPH-dependiente (enzima málica). Sería interesante ampliar este estudio a sustratos como la glucosa y el 3-hidroxibutirato. Se logró demostrar que la Acetil-CoA Sintetasa 1 (AceCS1) puede activarse cuando las concentraciones mitocondriales de citrato no son suficientes para mantener la lipogénesis en el citosol y presentando una mayor actividad en neuronas que astrocitos. El uso del inhibidor demostró que la concentración de oxalacetato es limitante en el metabolismo intermediario de las neuronas y los astrocitos, sin embargo, la capacidad anaplerótica astrocítica, debido a la actividad de la piruvato carboxilasa (PC) contribuye a mantener los niveles de este intermediario. Los astrocitos demostraron tener mayor capacidad de amortiguar los efectos del 3-NPA sobre la respiración que las neuronas, pudiendo ser atribuido a un conjunto de factores como compartimiento de expresión específica de la piruvato carboxilasa y malato deshidrogenasa-NADP dependiente citosólica (cME) y por otro lado a las diferentes tasas de recambio del ciclo de Krebs neuronal y astrocítico. La disminución de la respiración neuronal inducida por el 3-NPA en presencia de lactato y acetato, puede tener efectos negativos en el periodo perinatal, pues afecta significativamente la producción de lípidos y de ATP, debido la disminución de su capacidad anaplerótica.
Agradecimientos
Los autores agradecen a Angélica Pinzón por su colaboración en el mantenimiento de los animales de experimentación en el Bioterio de la Facultad de Ciencias (PUJ).
Financiación
La Vicerrectoría Académica de la Pontificia Universidad Javeriana dio su apoyo financiero para la elaboración de este trabajo (proyecto 1587).
Conflicto de intereses
Los autores declaran que no existen conflictos de intereses con este trabajo.
Referencias
1. Siesjo B. Brain energy metabolism. Wiley-Interscience Publication. New York, NY. 1978; 607 p. [ Links ]
2. Medina JM, Tabernero A, Tovar J, Martin-Barrientos J. Metabolic fuel utilization and pyruvate oxidation during the postnatal period. Journal of Inherited & Metabolic Diseases 1996; 19: 432-442. [ Links ]
3. Tovar J, Barrios B. Compartimentación intracelular del acetato en neuronas durante la prelactancia. Universitas Scientiarum 2006; 11: 73-86. [ Links ]
4. Tovar J, Saavedra F. Compartimentación intercelular del acetato en neuronas y astrocitos durante la prelactancia. Universitas Scientiarum 2005; 10: 21-38. [ Links ]
5. Tovar J. Salazar A. Compartimentación intracelular del acetato en astrocitos durante la prelactancia. Universitas Scientiarum 2005; 10: 39-50. [ Links ]
6. Zwingmann C, Leibfritz D. Regulation of glial metabolism studied by 13C-NMR. NMR in Biomedicine 2003; 16: 370-399. [ Links ]
7. Cohen, J, WiIlkin G. Neural cell culture. A practical approach. IRL press at Oxford University Press. London, England. 1995; 248 p. [ Links ]
8. Saneto R, De Vellis J., Neuronal and glial cells: Cell culture of the central nervous system. In: Turner AJ, Bachelard HS. Neurochemistry a Practical Approach. IRL Press. 1987; 27-64. [ Links ]
9. Kimelberg, H. Primary astrocyte cultures - A key to astrocite function. Cellular and Molecular Neurobiology 1983; 3 (1): 1-16. [ Links ]
10. Rose S, Sinha A. Some properties of isolated neuronal cell fractions. Journal of Neurochemistry 1969; 16: 1319-1328. [ Links ]
11. Edmond E, Robbins R, Bergstrom J, Cole R, De Vellis J. Capacity for substrates utilization in oxidative metabolism by neurons, astrocytes and oligodendrocytes from developing brain in primary culture. Journal of Neuroscience Research 1987; 18 (4): 551-561. [ Links ]
12. Sykes J., López-Cardoso M, Van Den Bergh S. Substrate utilization for energy production and lipid synthesis in oligodendrocyte-enriched cultures prepared from rat brain. Neurochemistry International. 1986; 8: 67-75. [ Links ]
13. Waniewski R, Martin D. Preferential utilization of acetate by astrocytes is attributable to transport. Journal of Neuroscience 1998; 14: 5225-5233. [ Links ]
14. Tovar J, Saavedra F, Bryón A. Metabolismo cerebral. En: Niño, M. y Ferrer, L. (eds.) Neuroanestesia. Enfoque perioperatorio en el paciente neurológico. Distribuna Editorial Médica. Bogotá, Colombia. 2005; 33-88. [ Links ]
15. Gamberino C, Berkich, D, Lynch C, Xu B, Lanoue K. Role of pyruvate carboxylase in facilitation o synthesis of glutamate and glutamine in cultured astrocytes. Journal of Neurochemistry 1997; 69(6): 2312-2325. [ Links ]
16. Hassel B, Brathe A. Neuronal piruvate carboxilation supports formation of transmitter glutamate. Neuroscience. 2000; 20: 1342-1347. [ Links ]
17. Albarracín S, Tovar J. Metabolismo del [U-14C]-acetato y del [2-14C]-acetato en astrocitos y astrocitoma C6. Biomédica 2002; 22 (Suplemento 1): 72. [ Links ]
18. Baslow M. N-Acetylaspartate in vertebrate brain: Metabolism and function. Neurochemical Research. 2003; 28 (6): 941-953. [ Links ]
19. Bakken I, Johnsen S, White L, Unsgard G, Aasly J, Sonnewald U. NMR spectroscopy study of the effect of 3-nitropropionic acid on glutamate metabolism in cultured astrocytes. Journal of Neuroscience Research 1997; 47 (6): 642-649. [ Links ]
20. Kaufman E, Driscoll B. Carbon dioxide fixation in neuronal and astroglial cells in culture. Journal of Neurochemistry. 1992; 58 (1): 258-269. [ Links ]
21. Hassel B. Pyruvate carboxylation in neurons. Neuroscience. 2001; 66: 755-762. [ Links ]
22. Hassel, B. Carboxylation and anaplerosis in neurons and glia. Neurobiology 2000. 22: 21-40. [ Links ]
23. Olsen C, Fonnum F, Paulsen, R, Hassel B. 3-Nitropropionic acid: an astrocyte-sparging neurotoxin in vitro. Brain Research. 1999; 850: 144-149. [ Links ]

