










Servicios Personalizados
Revista
Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkUniversitas Scientiarum
versión impresa ISSN 0122-7483
Univ. Sci. v.15 n.3 Bogotá sep./dic. 2010
Arvores prestadoras de serviços ecológicos na Amazônia Brasileira
Trees of ecological services in the Brazilian Amazon
Árboles de servicios ecológicos en Amazonía Brasileña
2Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA) - Centro de Pesquisa Agropecuária do Trópico Úmido - CPATU, Belém, Pará, Brasil.
Recebido: 28-10-2010: Aprovado:14-12-2010
Resumo
Objetivo. O metabolismo das rizosferas de recursos arbóreos potencialmente acumuladores de fósforo (P) e, portanto, prestadores de serviços ecológicos como Neea macrophylla e Cecropia palmata pode exercer certa influência sobre a dinâmica do solo próximo às mesmas como forma de tolerância a ambientes de estresse, sobretudo quanto às concentrações de fósforo (P), alumínio (Al) e Matéria Orgânica (MO). O objetivo deste trabalho foi caracterizar os perfis das rizosferas das espécies consideradas e determinar a biomassa lenhosa, foliar e total como alternativa de incremento de carbono produzida pelas mesmas. Metodologia. Foram selecionadas árvores (indivíduos) em duas florestas secundárias (capoeiras), uma enriquecida e outra somente com regeneração natural no Município de Igarapé-Açú, estado do Pará para monitoramento das concentrações de fósforo (P), alumínio (Al) e Matéria Orgânica (MO). Resultados. Os solos próximos às rizosferas de Neea se mostraram com maior teor de fósforo e Matéria Orgânica do que os de Cecropia, contudo as menores concentrações de alumínio foram observadas nos solos sob influência da rizosfera de Cecropia. Conclusões. Tendências de redução do Teor de fósforo e Matéria Orgânica em escala potencial com o aumento das concentrações de alumínio foram observadas para todas as espécies. A biomassa foliar não diferiu entre as espécies, apenas a lenhosa.
Palavras-chave: serviço ecológico, alumínio, floresta.
Abstract
The metabolism of rhizospheres of plants that accumulate phosphorus (P) as Neea macrophylla and Cecropia palmata can exert certain influence on soil dynamics as a tolerance against environmental stresses, mostly in relation to phosphorus (P), aluminum (Al) and organic matter (MO) concentrations. The objective of this work was to characterize the profiles of the rhizospheres of selected species and to determine wood, leaf and total biomass as an alternative to the increment of the carbon produced. Materials and methods. We selected individual trees in two secondary forests (capoeiras), one enriched and another one only with natural vegetation in the Igarapé-Açú City, state of Pará, for measuring the concentrations of phosphorus (P), aluminium (Al) and organic matter (MO). Results. Soil adjacent to the rhizosphere of Neea showed higher concentrations of phosphorus and organic matter than Cecropia, however low concentrations of aluminium had been observed in soil under the influence of Cecropia. Conclusions. Trends of a reduction on phosphorus and organic matter concentrations in a potential scale with the increase of aluminium concentrations had been observed in all the rhizospheres. Leaf biomass did not differ between species, while wood biomass did differ between species.
Key words: ecological service, aluminium, forest.
Resumen
El metabolismo de rizóforas de las plantas que acumulan el fósforo (P) como Neea macrophylla y Cecropia palmata pueden ejercer cierta influencia en la dinámica de suelo como forma de la tolerancia para suportar ambientes com estresse, sobre todo en lo referente a concentraciones del fósforo de (P), de aluminio (Al) y de la materia orgánica (MO). El objetivo de este trabajo fuecaracterizar los perfiles de los rizosferas de las espécies consideradas y determinar biomassa en la madera, la hoja y la biomasa total como alternativa del incremento del carbón producido. Materiales y métodos. Se seleccionaron árboles individuales em dos bosques secundários (Capoeiras), uno enriquecido y outro solamente com La vegetación natural en la ciudad de Igarapé-Açú, estado de Pará habían sido seleccionados, para supervisar las concentraciones de fósforo (P), de aluminio (Al) y de la materia orgánica (MO). Resultados. El suelo adyacente del rizosfera de Neea mostro concentraciones más altas de fósforo y de Materia Orgánica que Cecropia, no obstante pocas concentraciones de Al habían sido observadas en suelo bajo influencia del Cecropia. Conclusiones. Las tendencias de la reducción del fósforo y de las concentraciones de la Materia Orgánica en escala potencial con el aumento de las concentraciones del Al habían sido observadas para todo el rizosferas. No se presentaron diferencias entre lãs esp écies para La biomasa foliar, pero si entre ellas respecto a La biomasa de La madera.
Palabras clave: servicio ecológico, alumínio, bosque.
Introdução
As relações de interação solo-planta podem ou não favorecer as rizoferas das espécies, geralmente quando existe algum tipo de tolerância das espécies a determinado estresse é provável que o metabolismo, sobretudo radicular das plantas torne-se defensivo às condições do entorno alterando concentrações de determinados minerais do solo a fim de que sua sustentabilidade esteja garantida (Oliveira, 2007). Sendo assim, é de se esperar que recursos vegetais potencialmente acumuladores de fósforo (P) como Neea macrophylla e Cecropia palmata e, portanto prestadoras de serviços ecológicos encontradas em ambientes deficientes em P no Nordeste Paraense devam exercer trocas peculiares com o solo sob influência de suas rizosferas. Tais interações contextualizarão produções de biomassa aérea específicas, as quais segundo Davidson et al (2004) são constrangidas com a deficiência de P nos solos.
De acordo com EBERHARDT et al (2008) estudando os teores totais de P, assim como sua disponibilidade e retenção em Latossolos sob pastagens do Cerrado e relação destes dados com suas características mineralógicas, granulométricas e químicas, observou que houve uma correlação negativa entre o P remanescente e o C total, explicada pelo fato de que o estoque de C aumenta linearmente com o conteúdo de argila e silte no solo. Além disso, o P disponível resina mostrou-se correlacionado negativamente com as características mineralógicas e granulométricas e positivamente com os teores de matéria orgânica do solo. Estes dados sugerem, portanto, que um aumento na matéria orgânica do solo leva a um aumento do fósforo, o que é fundamental para a nutrição de plantas e ambientes, quer no cerrado, quer na floresta. O mesmo autor ainda observou que os óxidos de Fe e Al foram os componentes mineralógicos que exerceram a maior influência sobre o P remanescente.
Em florestas secundárias amazônicas, condições de elevados teores de ferro e alumínio nos solos são comuns, o que também poderia levar à restrição de fósforo para as plantas e para o ambiente produtivo ou natural no Bioma Amazônia além de perdas de nitrogênio segundo Neill et al (2006).
Outros fatores, no entanto, também podem exercer influencia sobre a nutrição de vegetações quanto às restrições de fósforo. De acordo com Bonfim (2004) estudando a influência de características físicas e químicas relacionadas com o fator capacidade de P (FCP) na determinação dos níveis críticos de P pelos extratores Mehlich-1, Mehlich-3 e Bray-1 e na parte aérea de plantas, encontrou resultados que demonstraram que as características que melhor refletiram o FCP foram a capacidade máxima de adsorção de P (CMAP) e o P remanescente. Foi observado que os níveis críticos de P no solo e em plantas de B. brizantha foram menores em solos com maior CMAP e com menor P remanescente.
Tais resultados sugerem que quanto menor for a capacidade dos solos de adsorverem fósforo, menor será a CMAP. Daí a importância das concentrações de matéria orgânica dos solos, como componentes doadores de fósforo orgânico.
Dessa forma, espécies prestadoras de serviços ecológicos, como as acumuladoras de fósforo em situações de estresse deste mineral nos solos, podem contribuir positivamente na manutenção dos níveis adequados de fósforo em Eco(agro)sistemas e por conseguinte da CMAP.
Ecossistemas com baixos teores de fósforo como os agroecossistemas amazônicos, sobretudo no contexto da agricultura familiar, onde não existe adição de adubação fosfatada acabam impondo uma maior tolerância às vegetações em processos de regeneração ao estresse por fósforo, haja vista, que a única forma de reposição de fósforo é o fósforo orgânico oriundo da decomposição de folhas e galhos. Este fato é comprovado em outros trabalhos como o de Gatiboni et al (2007), os quais avaliando a biodisponibilidade das formas de P acumuladas em um solo cultivado sob sistema de plantio direto com diferentes quantidades de P adicionado, observaram que em solos com baixa ou nenhuma adição de fertilizantes fosfatados, as formas orgânicas de P são as principais mantenedoras do P absorvido pelas plantas.
Neste contexto, espécies acumuladoras de fósforo em tecidos foliares e lenhosos pode ser fundamental na manutenção das concentrações de fósforo nos eco (agro) sistemas através do processo de decomposição orgânico da serrapilheira. Atrelado a isto, de acordo com Nziguheba (2002) em sistemas de agricultura familiar o aporte de adubações fosfatadas em solos cultivados com milho provocam ao longo do tempo uma reposição deste mineral extraido pelas culturas em solos deficientes.
Espécies prestadoras de serviços ecológicos, como as acumuladoras de fósforo, conseguem tal nível nutricional em condições normais de acidez do solo (ph = 4); porém de acordo com Souza et al (2006), o mesmo ressalta a importância da correção da acidez e elevação dos teores de matéria orgânica dos solos para a otimização do uso do P pelas culturas. É de se esperar, portanto altos padrões de eficiência no uso do fósforo por vegetações em cenários de regeneração de floresta secundária na Amazônia brasileira, haja vista, elevada acidez.
Processos de interferência em florestas secundárias locais como o Enriquecimento de capoeiras, uma tentativa recente no Nordeste Paraense com fins ao acúmulo de biomassa aérea durante a fase de pousio agrícola conforme experimento desenvolvido no Município de Igarapé-Açú por Brienza Júnior et al (1999) deve ser olhado com cautela. Segundo os autores a introdução de espécies exóticas e comuns na Região como Acácia mangium, Clitoria racemosa, Inga edulis e Sclerolobium paniculatum aumentou a produção de biomassa em relação à biomassa produzida nos sistemas de pousio tradicional. Contudo, uma redução na regeneração natural da capoeira foi observada, sobretudo nos menores espaçamentos. Estes fatos sugerem que estudos acerca do comportamento ecofisiológico de tais espécies devem ser mais bem conhecidos antes de plantios sistemáticos como forma de evitar distúrbios ambientais locais no que tange à competição por recursos do solo e do ar bem como acerca da riqueza florística local. Neea macrophylla e Cecropia palmata caracterizaram-se como espécies pioneiras haja vista sua ocorrência logo após a derrubada destas árvores plantadas.
Desta forma, esta pesquisa teve por objetivo a caracterização da Biomassa Foliar, Lenhosa e Total de duas espécies regenerantes, N.macrophylla e C.palmata em capoeira enriquecida (situs II) e não enriquecida (situs I) bem como o delineamento do perfil do solo sob a influência das rizosferas das espécies estudadas quanto aos Teores de Fósforo (P), Alumínio (Al) e Matéria Orgânica (MO).
As espécies N.macrophylla e C.palmata são espécies que ocorrem ao longo do processo de regeneração das capoeiras no Bioma Amazônia, território Amazônia Legal, entre 2 a 8 anos aproximadamente. São espécies tolerantes a baixos níveis de fósforo e intensa radiação luminosa. N.macrophylla pertence à família das Nyctaginaceaes, enquanto C.palmata às Cecropiaceaes.
Material e métodos
A pesquisa foi desenvolvida no Município de Igarapé-Açú em duas capoeiras com idades distintas (8 anos, situs II e 6 anos, situs I) onde foram selecionadas 6 árvores por espécie, N.macrophylla e C. palmata. Foram então feitas coletas de solo próximo às rizosferas das espécies para determinação dos teores de P (mg.dm-3), Al (cmol.dm-3) e Matéria Orgânica (g.kg-1) em três profundidades, 0-5, 5-10 e 20-40cm ao longo do perfil do solo (Oxisol, latossolo amarelo, textura média) e em 3 posições à 40, 80 e à 300 cm da árvore.
Os solos da capoeira estudada se caracterizavam nos primeiros 5 cm de profundidade de solos por ph em torno de 5.2, C (1,5%), N (%) 0.12, P (2mg/kg), Al (12%), C/N (12) e P/N (0.04). As condições climáticas locais se caracterizaram por um total de chuva anual entre 2300 e 2.800 mm, temperatura media anual de 26°C, com fotoperiodo de doze horas.
A produção de biomassa aérea foliar, lenhosa e total (kg) foi obtida pela média de 6 indivíduos por espécie estudada nas duas capoeiras. As amostras foram secas em estufa à temperatura de 65oC até peso constante.
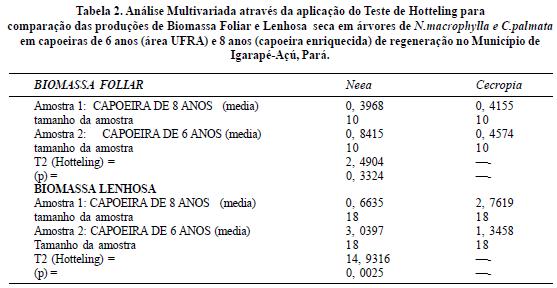
Embora, diferentes metodologias podem ser utilizadas no estudo da dinâmica de fósforo em agroeco-sistemas (Kolawole 2003, Davidson1999, Lehmann 2001,Lima 2009, Makoto 2010, Wood 2008, Duran 2010, Unger 2010, Giardina 2000,Smithson 2002, Atkinson 2010, Kennard 2001, Silva 2010 e Wu 2010), este trabalho aplicou inicialmente o Teste Tukey para comparação das médias de biomassa foliar e lenhosa entre as espécies estudadas na capoeira mais nova. A seguir foi feita a Análise Multivariada através da aplicação do Teste de Hotteling para comparação tanto da biomassa foliar quanto da lenhosa entre as duas capoeiras a fim de verificar possíveis efeitos da idade, embora a diferença tenha sido pouca entre as mesmas. O programa estatístico utilizado foi o BioEstat 4.0 (Ayres et al, 2005).
Resultados e discussões
Os resultados obtidos quando avaliamos o teor de P no solo demonstraram que o solo da rizosfera de N. macrophylla reteve mais P (mg.dm-3) nas camadas mais superficiais, 0-5 cm e 5-10 cm reduzindo gradativamente até 40 cm de profundidade tanto à 20 como 80 cm de distância das árvores em relação à C. palmata, (Figuras 1 e 2). Contudo, no entorno das árvores, i.e., à 300 cm de distância onde não haviam as 2 espécies estudadas, os teores de P não diferiram significativamente (Figura 3). Dessa forma, parece que tanto N. macrophylla quanto C. palmata são capazes de propiciar uma situação favorável de fertilidade do solo através do aumento do teor de P na projeção de suas copas, sobretudo N. macrophylla através de uma correlação positiva com a profundidade do solo.
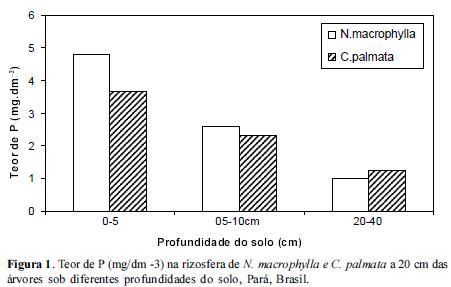
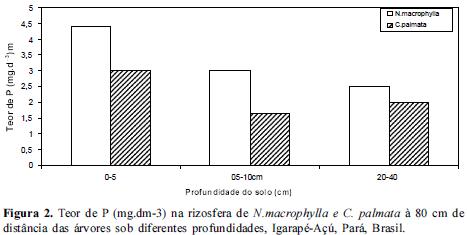
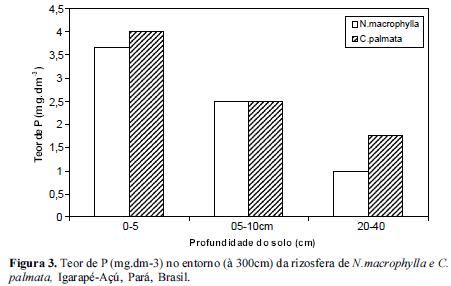
Segundo Oberson et al (2001), outras situações também podem oferecer uma maior disponibilidade de fósforo às plantas, como por exemplo o plantio de leguminosas consorciadas.
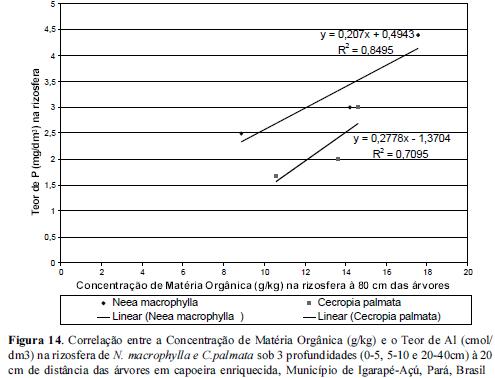
Quanto ao teor de Al no solo da rizosfera das espécies estudadas, observou-se um aumento do mesmo a partir da camada superior em direção a mais profunda (20-40) em todas as distâncias conforme figuras 4, 5 e 6. Os teores mais elevados de Al encontrados no extrato de 20-40 cm de profundidade poderão estar influenciando o teor de P disponível para as raízes localizadas neste perfil, normalmente raízes secundárias mais grossas provavelmente ainda relacionadas com o processo de difusão do P. De acordo com Silva (2009) a rizotoxidez ao alumínio pode ser atenuada pelo efeito protetor do Magnésio (Mg), dessa forma é de se esperar que entre 20 a 40 cm de profundidade nestas rizosferas etudadas provavelmente a rizotoxidez por aluminio deve estar ocorrendo.
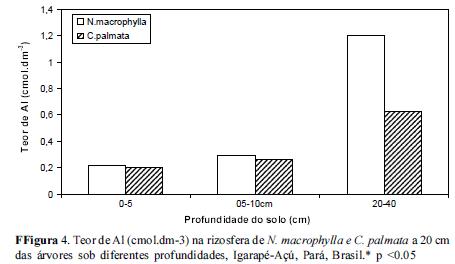
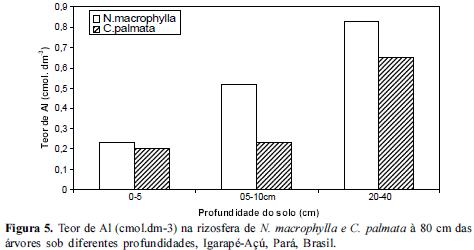
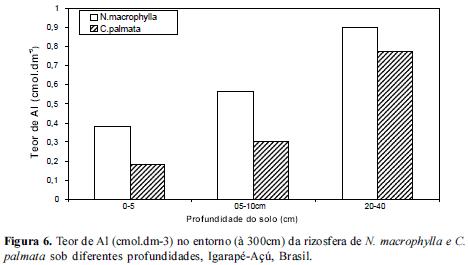
A rizosfera de Cecropia palmata apresentou uma menor concentração de Al em todas as profundidades em relação à rizosfera de Neea macrophylla e portanto criando um meio com um elemento a menos para fixar-se ao P, podendo torná-lo às vezes mais disponível para as plantas. Cecropia palmata uma vez selecionada para o enriquecimento de vegetações secundárias ou em Sistemas Agroflorestais como componente arbóreo atenuador das condições de toxidez de Al, pode vir a ser uma estratégia de ganho de sustentabilidade em sistemas agrícolas familiares em que a capoeira é fase imprescindível nos ciclos agrícolas locais.
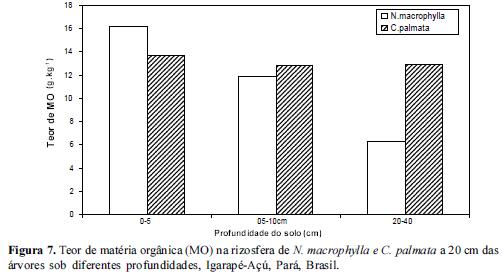
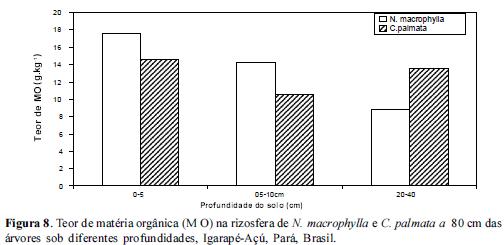
Quanto à concentração da matéria orgânica (MO), esta foi maior em N.macrophylla em todas as profundidades e nas 2 distâncias em relação a C. palmata conforme figuras 7, 8 e 9. Isto sugere que mecanismos de decomposição da liteira associados à qualidade da liteira devem exercer um papel mais acentuado na rizosfera de N.macrophylla do que na de C. palmata. A disponibilização do P orgânico para o solo tende então a ser mais rápida em rizosferas com maior teor de MO em condições naturais. Estes dados estão de acordo com Pandey (2009) o qual observou que um aumento no carbono lábil do solo associado a um aumento na intensidade de chuvas provocava um aumento no fósforo extraído por anaerobiose em florestas amazônicas.
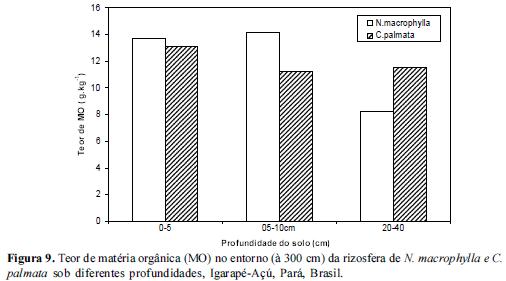
Da mesma forma, estão de acordo com Zaia et al (2008), onde os mesmos identificaram que o tipo de cobertura vegetal altera os niveis de fósforo nos solos através de resultados observados em plantios de Acacia auriculiformis (acácia) e Mimosa caesalpiniifolia (sabiá), em floresta secundária e em pastagem, onde o maior teor relativo de Po (total e lábil) foi observado em plantios de acácia do que nos solos sob pastagem e capoeira. Neste contexto, identificar espécies prestadoras de serviços ecológicos, especificamente com potencial para manter níveis estáveis de fósforo total é fundamental em processos de recuperação de área degradada.
O enriquecimento de vegetações secundárias ou de Sistemas Agroflorestais com espécies potencialmente acumuladoras de P como N.macrophylla, com o objetivo de colocar um componente arbóreo capaz de aumentar os níveis de MO, poderá exercer influência marcante no ciclo biogeoquímico do P através de um aumento na disponibilização do P total. A faixa de maior concentração de MO foi de 0-5 e 5-10 para ambas espécies estudadas, reduzindo significativamente no extrato de 20-40 cm. Estes resultados estão no mesmo sentido dos resultados acerca dos Teores de P e inversamente aos resultados obtidos para a concentração de Al.
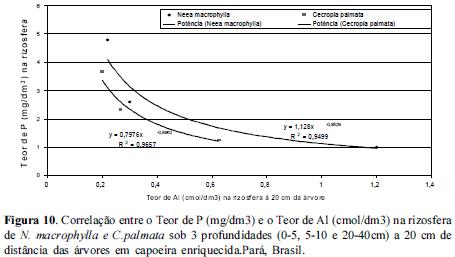
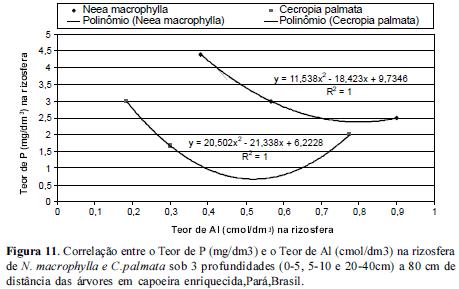
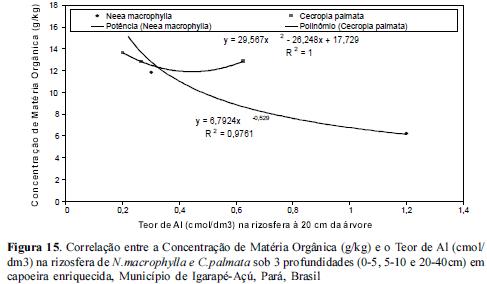
Quando observamos a correlação entre as concentrações de P e Al nas rizosferas das 3 espécies estudadas notamos que à medida que as concentrações de P diminuem com o aumento da profundidade, as de Al aumentam, com um coeficiente de correlação igual à 100% em um modelo de ajuste polinomial tanto para N. macrophylla quanto para C. palmata, conforme figuras 10 e 11.
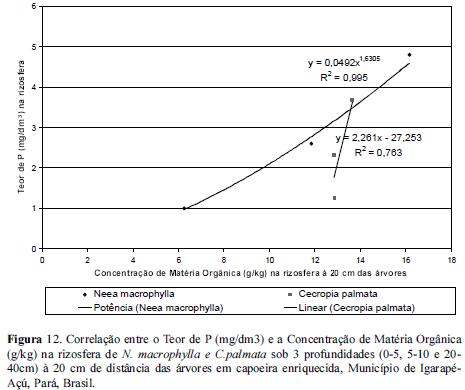
No entanto, quando correlacionamos as concentrações de P com as de Matéria Orgânica, tanto à 20 quanto à 80 cm de distância das árvores as concentrações de P tenderam a aumentar com o aumento da concentração de matéria orgânica sendo estas maiores nas rizosferas de N. macrophylla e menores nas de C.palmata com um ajuste do tipo linear entre as 2 variáveis, conforme figuras 12 e 13. Contudo, segundo Beck & Sanchez (1996) o conhecimento de outros fatores como o fósforo pouco lábil e resíduos de fertilizantes são necessários ao entendimento da dinâmica de fósforo nos solos.
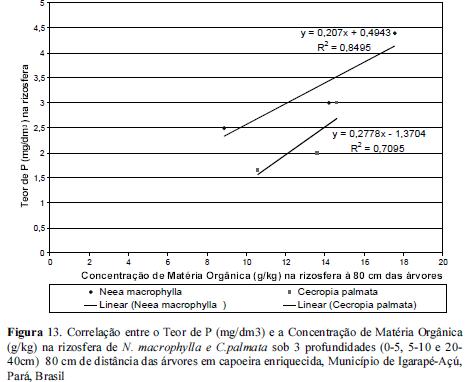
Os resultados acerca das correlações entre as concentrações de Matéria Orgânica e de aluminio demonstraram que à medida que se reduz as concentrações de matéria orgânica aumentam as de alumínio à 20 cm das árvores, onde nas rizosferas de C. palmata foi observada uma baixa amplitude de Al e matéria orgânica em todas as 3 profundidades estudadas em relação às rizosferas de N.macrophylla, conforme figuras 14 e 15.
Quanto aos resultados acerca da produção de biomassa em capoeira com 6 anos de regeneração (stio I) foi observada ausência de diferença significativa entre as médias da biomassa foliar de N.macrophylla e C. palmata com P=0,15 conforme tabela 1 (Análise de Variância a partir da aplicação do Teste Tukey).
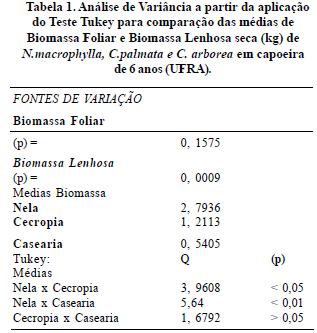
Estes dados sugerem que a escolha de uma ou outra espécie acumuladora de P, Neea ou Cecropia pode fornecer quantidades similares de tecido verde uma vez derrubada. Contudo o mesmo não aconteceu com a biomassa lenhosa que diferiu a nivel de 1% de probabilidade entre Neea, Cecropia e Casearia. N.macrophylla foi a espécie que apresentou maior biomassa lenhosa, logo se considerarmos este fator então Neea é mais viável como componente ecológico em Sistemas Agroflorestais do que C.palmata.
No entanto quando comparamos as biomassas médias foliares entre as duas capoeiras com idades distintas observamos que a biomassa foliar não diferiu significativamente embora a biomassa lenhosa tenha diferido de acordo com a tabela 2, o que nos leva a pensar que o efeito da idade da regeneração pode ser melhor expresso a partir da biomassa lenhosa do que foliar. A biomassa lenhosa de Neea em capoeira com 6 anos foi nitidamente superior a de 8 anos, sugerindo que na floresta de 6 anos as condições propícias ao início da regenereração desta espécie se deu mais cedo do que na floresta mais antiga, indicando um certo retardamento relativo do processo de regeneração decorrente talvez do enriquecimento realizado nesta área? C.palmata por sua vez apresentou comportamento oposto ou seja, obteve maior produção de biomassa lenhosa na floresta de 8 anos sugerindo melhor adaptabilidade nesta área do que na de 6 anos conforme figura 16.
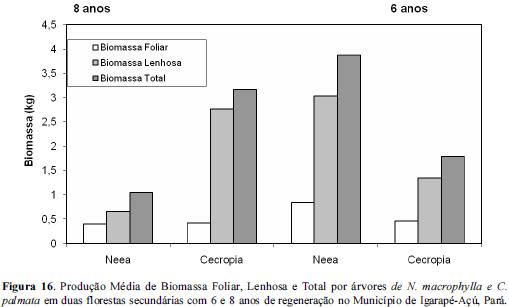
Conclusões
A rizosfera de N.macrophylla acumulou mais fósforo e Matéria orgânica do que a de C.palmata sugerindo melhores condições de disponibilidade do nutriente às plantas situadas sob influência dessa espécie. As rizosferas de C. palmata, no entanto, obtiveram as menores concentrações de Alumínio o que significa um fator a menos no envolvimento da retenção do P às plantas situadas em tais rizosferas. Foi observada uma tendência de redução do fósforo e da Matéria Orgânica com o aumento das concentrações de alumínio para Neea e Cecropia. A produção de Biomassa Foliar não diferiu entre as espécies e tão pouco com a diferença de idade de regeneração, porém foi a produção de Biomassa Lenhosa que mostrou ser o melhor parâmetro diferencial entre as espécies prestadoras de serviços ecológicos, como as acumuladoras de fósforo.
Referências
1. Ayres M, Ayres Junior, Ayres DL, dos Santos AS. BioEstat 4.0. Aplicações estatísticas nas áreas das Ciências Biológicas e Médicas. Sociedade Civil Mamirauá/MCT-CNPQ/Conservation International, Belém-Pará, 2005. [ Links ]
2. Beck M, Sanchez PA Soil phosphorus movement and budget after 13 years of fertilized cultivation in the Amazon basin, Plant and Soil, 1996, 184, 1, 23-31. [ Links ]
3. Bonfim EMS. Níveis críticos de fósforo para Brachiaria brizantha e suas relações com características físicas e químicas em solos de Pernambuco. Revista Brasileira de Ciência do Solo. 2004; 28 (2): 281-288. [ Links ]
4. Brienza Junior S, Costa VO, Santos WE, Pantoja R, Sá TDA, Vielhauer K, Denich M, Vlek P. Enriquecimento de capoeira com árvores leguminosas contribuindo para o acúmulo de biomassa na agricultura familiar do Nordeste do Estado do Pará, Brasil. In: seminário para o manejo da vegetação secundária para a sustentabilidade da agricultura familiar da amazônia oriental. Anais 2000, Belém, 83 p. [ Links ]
5. Davidson E A, Carvalho C.J.R, Vieira IC, Figueiredo R O, Moutinho P, Ishida F Y, Santos MTP. Nitrogen and phosphorus limitation of biomass growth in a tropical secondary forest. Ecological Applications, 2004, 14 (4): 150-163. [ Links ]
6. Davidson R, Gagnon D and Mauffette Y. Growth and mineral nutrition of the native trees Pollalesta discolor and the N-fixing Inga densiflora in relation to the soil properties of a degraded volcanic soil of the Ecuadorian Amazon, Plant and Soil, 1999, 208, 1, 135-147. [ Links ]
7. Durán J, Rodríguez A, Fernández-Palacios JM, Gallardo A. Changes in leaf nutrient traits in a wildfire chronosequence, Plant and Soil, 2010, 331, 1-2, 69-77. [ Links ]
8. Eberhardt, D N. Influência da granulometria e da mineralogia sobre a retenção do fósforo em latossolos sob pastagens no cerrado. Revista Brasileira de Ciência do Solo. 2008; 32 (3):1009-1016. [ Links ]
9. Gatiboni LC. Biodisponibilidade de formas de fósforo acumuladas em solo sob sistema plantio direto. Revista Brasileira de Ciência do Solo. 2007; 31 (4): 691-699. [ Links ]
10. Giardina CP, Sanford RL, Døckersmith IC, Jaramillo VJ. The effects of slash burning on ecosystem nutrients during the land preparation phase of shifting cultivation. Plant and Soil, 2000, 220, 1-2, 247-260. [ Links ]
11. Kennard DK, Gholz HL. Effects of high- and lowintensity fires on soil properties and plant growth in a Bolivian dry forest, Plant and Soil, 2001, 234, 1, 119-129. [ Links ]
12. Kolawole G O, Tian G, Tijani-Eniola. Dynamics of phosphorus fractions during fallow with natural vegetation and planted Pueraria phaseoloides in south-western Nigeria. Plant and Soil, 2003, 257, 1, 63-70. [ Links ]
13. Kolawole G O, Tijani-Eniola, Tian G. Phosphorus fractions in fallow systems of West Africa: Effect of residue management, Plant and Soil, 2004, 263, 1, 113-120. [ Links ]
14. Lehmann J, Silva JP, Steiner C, Nehls T, Zech W. Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: fertilizer, manure and charcoal amendments, Plant and Soil, 2003, 249, 2, 343-357. [ Links ]
15. Lehmann J, Cravo MS, Macêdo JLV, Moreira A, Schroth G. Phosphorus management for perennial crops in central Amazonian upland soils, Plant and Soil 2001, 237, 2, 309-319. [ Links ]
16. Lima AS, Nóbrega RSA, Barberi A, Silva K, Ferreira DF. Nitrogen-fixing bacteria communities occurring in soils under different uses in the Western Amazon Region as indicated by nodulation of siratro (Macroptilium atropurpureum), Plant and Soil, 2009, 319, 1-2, 127-145. [ Links ]
17. Makoto K, Tamai Y, Kim YS, Koike T. Buried charcoal layer and ectomycorrhizae cooperatively promote the growth of Larix gmelinii seedlings, Plant and Soil, 2010, 327, 1-2, 143-152. [ Links ]
18. Martinazzo R. Fósforo microbiano do solo sob sistema plantio direto em resposta à adição de fosfato solúvel. Revista Brasileira de Ciência do Solo. 2007; 31 (3): 563-570. [ Links ]
19. Neill C, Piccolo M, Cerri C, Steudler P, Melillo J Soil. Solution Nitrogen Losses During Clearing of Lowland Amazon Forest for Pasture, Plant and Soil, 2006, 281, 1-2, 233-245. [ Links ]
20. Nziguheba G, Merckx R, Cheryl A. Palm Soil phosphorus dynamics and maize response to different rates of phosphorus fertilizer applied to an Acrisol in western Kenya, Plant and Soil, 2002, 243, 1, 1-10. [ Links ]
21. Oberson A, Friesen D. K., Rao I.M., Bühler S., Frossard E. Phosphorus Transformations in an Oxisol under contrasting land-use systems: The role of the soil microbial biomass, Plant and Soil, 2001, 237, 2, 197-210. [ Links ]
22. Pandey C B, Srivastava R C. Plant available phosphorus in homegarden and native forest soils under high rainfall in an equatorial humid tropics, Plant and Soil, 2009, 316, 1-2, 71-80. [ Links ]
23. Silva I R. Timing, location and crop species influence the magnitude of amelioration of aluminum toxicity by magnesium. Revista Brasileira de Ciência do Solo. 2009; 33 (1): 65-76. [ Links ]
24. Silva LCR, Haridasan M, Sternberg LSL, Franco AC, Hoffmann WA. Not all forests are expanding over central Brazilian savannas. Plant and Soil, 2010, 333, 1-2, 431-442. [ Links ]
25. Smithson PC, Giller KE. Appropriate farm management practices for alleviating N and P deficiencies in lownutrient soils of the tropics. Plant and Soil, 2002, 245, 1, 169-180. [ Links ]
26. Souza RF. Calagem e adubação orgânica: influência na adsorção de fósforo em solos. Revista Brasileira de Ciência do Solo. 2006; 30 (6): 975-983. [ Links ]
27. Unger M, Leuschner C, Homeier J. Variability of indices of macronutrient availability in soils at different spatial scales along an elevation transect in tropical moist forests (NE Ecuador), Plant and Soil, 2010, 336, 1-2, 443-458. [ Links ]
28. Zaia FC, Gama-Rodrigues AC, Forestieri E. Formas de fósforo no solo sob leguminosas florestais, floresta secundária e pastagem no Norte Fluminense. Revista Brasileira de Ciência do Solo. 2008; 32 (3): 1191-1197. [ Links ]
29. Wood TE, Lawrence D. No short-term change in soil properties following four-fold litter addition in a Costa Rican rain forest, Plant and Soil, 2008, 307, 1-2, 113-122. [ Links ]
30. Wu GL, Liu ZH, Zhang L, Hu TM, Chen JM. Effects of artificial grassland establishment on soil nutrients and carbon properties in a black-soil-type degraded grassland, Plant and Soil, 2010, 333, 1-2, 469-479. [ Links ]

