










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkUniversitas Scientiarum
versão impressa ISSN 0122-7483
Univ. Sci. vol.20 no.1 Bogotá jan./abr. 2015
https://doi.org/10.11144/Javeriana.SC20-1.baha
Bacterias autótrofas y heterótrofas asociadas a nieve marina lodosa en arrecifes con escorrentía continental
Autotrophic and heterotrophic bacteria associated to muddy marine snow in reefs with terrestrial runoff
Bacterias autótrofas e heterotróficas associadas a neve marinila lodosa em recifes com escoamento continental
Alejandro Henao-Castro1, Natalia Comba González2, Elvira M. Alvarado Ch3, Johanna Santamaría4
Edited by Alberto Acosta
1 Universidad Jorge Tadeo Lozano, Bogotá, Colombia.
2 Universidad Nacional de Colombia, Bogotá, Colombia.
3 Universidad Jorge Tadeo Lozano, Programa de Biología Marina, Bogotá, Colombia.
4 Universidad Jorge Tadeo Lozano, Facultad de Ciencias Naturales e Ingenierías, Bogotá, Colombia.
Funding: Universidad Jorge Tadeo Lozano; Fundación Diving Planet; Parque Nacional Corales del Rosario y San Bernardo.
Electronic supplementary material: N/A
Received: 20-03-2014 Accepted: 25-05-2014 Published on line: 10-07-2014
Para citar este artículo / To cite this article
Henao-Castro A, Comba N, Alvarado-Chacón E, Santamaría J (2015) Bacterias autótrofas y heterótrofas asociadas a nieve marina lodosa en arrecifes con escorrentía continental. Universitas Scientiarum 20(1): 9-16 doi: http://dx.doi.org/10.11144/Javeriana.SC20-1.baha
Resumen
En agregados de nieve marina influenciada por escorrentía continental, la densidad de bacterias heterótrofas es mayor que la de autótrofas. Esta premisa fue puesta a prueba en cuatro zonas arrecifales localizadas a diferente distancia del Canal del Dique; principal fuente de aportes continentales a los arrecifes de coral del Archipiélago Nuestra Señora del Rosario, Cartagena, Caribe colombiano. Mediante epifluorescencia se determinó densidad promedio de microorganismos presentes en agregados de nieve marina. Los resultados mostraron mayor densidad de bacterias heterótrofas (3.63 x 104 ± 1.6 x 104 SE células mL-1) que de autótrofas (6 x 103± 1.3 x 103 SE células mL-1), principalmente en arrecifes cercanos a descarga de escorrentía continental (bacterias heterótrofas con densidad 8.9 x 104 células mL-1 y 3 x 104 células mL-1 para Isla Arena y Tesoro, respectivamente). La densidad de microorganismos encontrada es típica de zonas con alto contenido de materia orgánica particulada; por lo cual podría servir como potencial indicador de escorrentía continental. Estudios futuros deben enfocarse en determinar la composición de comunidad bacteriana asociada a nieve marina y posible virulencia sobre organismos arrecifales.
Palabras clave: arrecifes coralinos; bacterias; epifluorescencia; escorrentía continental; nieve marina lodosa.
Resumo
Em aglomerados de neve marinha influenciados por escoamento continental, a densidade de bacterias heterotróficas é maior que a de autótrofas. Esta premissa foi posta a prova em quatro zonas de recifes localizadas em diferentes distancias do Canal de Dique; principal fonte de contribuic.ao continental aos recifes do coral do Arquipélago Nossa Senhora do Rosàrio, Cartagena, Caribe colombiano. Mediante epifluorescencia determinou-se densidade de micro-organismos presentes em aglomerados de neve marinha. Os resultados mostraram maior densidade de bacterias heterotróficas (3.63 x 104 ± 1.6 x 104 SE células mL-1) que de autótrofas (6 x 103 ± 1.3 x 103 SE células mL-1), principalmente em recifes próximos a descarga de escoamento continental (bacterias heterotróficas com densidade 8.9 x 104 células mL-1 y 3 x 104 células mL-1 para a Ilha Arena e Tesoro, respetivamente). A densidade de micro-organismos é típica de zonas com elevado conteúdo de matéria orgànica particulada; pela qual pode servir como potencial indicador de escoamento continental. Estudos futuros devem enfocar-se em determinar a composicäo da comunidade bacteriana associada a neve marinha e possível virulencia sobre organismos recifais.
Palavras-chave: recifes coralinos; bacterias; epifluorescencia; escoamento continental; neve marinha lodosa.
Abstract
The density of heterotrophic bacteria is greater than autotrophic in marine snow aggregates influenced by continental runoff. Four coral reef areas at different distances from the Canal del Dique served to evaluate this premise; this canal is the main source of inland resources for the coral reefs of the Nuestra Señora del Rosario archipelago in Cartagena in the Colombian Caribbean. The average density of microorganisms in marine snow aggregates was determined using epifluorescence. The results showed higher density of heterotrophic bacteria (3.63 x 104 ± 1.6 x 104 SE cells mL-1) than autotrophic (6 x 103± 1.3 x 103 SE cells mL-1), mainly in reefs near continental runoff discharges (heterotrophic bacteria density of 8.9 x 104 cells mL-1 and 3 x 104 cells mL-1 Isla Arena and Tesoro, respectively). The density of microorganisms found is typical of high-particulate organic matter areas and, therefore, could be a potential indicator of continental runoff. Future studies should focus on determining the composition of the bacterial community associated with marine snow and its potential virulence on reef organisms.
Keywords: Coral Reefs; bacteria; epifluorescence; muddy marine snow; terrestrial runoff.
Introducción
Los arrecifes de coral generalmente se asocian con aguas claras y oligotróficas (Stoddart 1969). No obstante, en la columna de agua se encuentran partículas de exopolímeros transparentes, conocidos como nieve marina, que son liberadas a través de procesos biológicos de los organismos planctónicos (Riebesell 1992) y bentónicos (Wolanski et al. 2003). La nieve marina se presenta en todos los océanos y su dinámica y distribución es regulada por factores ambientales (e.g. temperatura, salinidad, nutrientes) y biológicos como producción de mucus, heces y afloramiento fitoplanctónico (Alldredge & Silver 1988, Turley 2002). Los agregados de nieve marina albergan poblaciones bacterianas que presentan mayor actividad enzimática que la registrada por bacterias de vida libre, siendo responsables de procesos biológicos como la producción de materia orgánica realizada por bacterias autótrofas, así como la descomposición y remineralización de nutrientes en la columna de agua realizados por grupos de bacterias y protozoos heterótrofos, que habitan densamente los agregados de nieve marina (Alldredge et al. 1986, Kaltenbóck & Herndl 1992, Ziervogel et al. 2010). Sin embargo, cuando los ríos entran al mar, los nutrientes que son adsorbidos por los granos de sedimento fino (< 63 um) se vuelven lábiles y las partículas son rápidamente colonizadas por microorganismos productores de mucopolisacáridos (Wolanski et al. 2003). Este tipo de mucosidad aumenta la viscosidad de las partículas, lo que resulta en mayor agregación y finalmente en la formación de nieve marina lodosa (Fabricius & Wolanski 2000). Debido a sus altas tasas de sedimentación, los agregados aceleran el transporte vertical de la materia orgánica particulada desde la superficie del océano hacia el fondo (Asaeda & Wolanski 2002, Wolanski et al. 2003).
La nieve marina lodosa, generada a partir de la interacción entre la escorrentía continental y el agua marina, tiene efectos perjudiciales e incluso letales sobre los arrecifes de coral al servir como sustrato para el crecimiento de bacterias, en su mayoría heterótrofas (Luna et al. 2002), muchas de las cuales son responsables del surgimiento de enfermedades coralinas (Alldredge et al. 1990, Fabricius et al. 2003, Wolanski et al. 2003). Así, es probable que la reducción en el reclutamiento coralino, junto con la promoción del crecimiento de macroalgas y el incremento de sustrato físico y químico para especies microbianas patógenas, reflejen los efectos negativos más significativos de la escorrentía terrestre en los arrecifes de coral (Fabricius 2005, Lyons et al. 2007, Wolanski et al. 2008, Otero 2009, Lyons et al. 2010).
Al igual que en todo el Caribe, durante las últimas décadas se ha reportado un detrimento coralino en los arrecifes del Archipiélago Nuestra Señora del Rosario, evidenciado por la pérdida de cobertura de coral y el aumento de cobertura macroalgal (Alvarado et al. 2011, Rodríguez et al. 2010, Vega et al. 2011). Lo anterior ha sido atribuido a la dinámica existente entre escorrentía y zonas arrecifales, la cual parece estar afectada directamente por aportes del Canal del Dique (Garzón & Díaz 2003, Restrepo et al. 2006). Aunque no existe una evidencia sólida de la relación directa entre el estado arrecifal y los aportes continentales, se sugiere que la escorrentía del Canal del Dique, junto con partículas en suspensión y altas concentraciones de nutrientes alteran los procesos biológicos en los arrecifes. Adicionalmente, es probable que en esta zona se presenten procesos físicos y químicos entre las partículas, los nutrientes y microorganismos, que junto con las tasas de sedimentación y resuspensión de sedimentos finos, resultan en la formación de nieve marina lodosa (Restrepo & Alvarado 2011, Henao 2013). Por lo anterior y sumado a la falta de conocimiento en este tema para el Caribe colombiano, se estudió la presencia de nieve marina lodosa en los arrecifes coralinos del Archipiélago, y se determinó la abundancia de bacterias autótrofas y heterótrofas asociadas a esta clase de agregados. La información generada servirá de punto de partida para futuras investigaciones que comprueben los efectos deletéreos de la nieve marina lodosa en los arrecifes del Caribe colombiano, teniendo en cuenta no sólo los efectos directos como ahogamiento debido a la alta tasa de sedimentación y adherencia; sino también los indirectos como vector de enfermedades del coral, las cuales son cada vez más comunes (Navas et al. 2008, Navas et al. 2010).
Materiales y métodos
Se obtuvieron muestras de nieve marina de arrecifes de coral en cuatro puntos de muestreo (Figura 1): Isla Fiesta (N 10°11'9; 4'' W 75° 43' 41,5") e Isla Arena (N10° 8' 24; 6'' W 75° 43' 42,2") que corresponden a sitios más cercanos a la escorrentía continental e, Isla Tesoro (N 10° 13' 50; 9'' W 75° 44' 30") e Isla San Martín (N 10° 10' 18; 8'' W 75° 45' 56,2"), que son los sitios más alejados del foco de disturbio. En cada estación, la identificación de las partículas de nieve marina se realizó por observación directa y la extracción de la muestra se llevó a cabo utilizando jeringas de 50 ml (sin aguja) mediante buceo autónomo (suficiente para proceder a realizar análisis de epifluorescencia). Las muestras extraídas se consideran nieve marina lodosa por la presencia de aglomerados de materia orgánica y bacterias que conforman largas cadenas de filamentos, así como la presencia de partículas finas de sedimentos (< 63 um) corroborado con observación en microscopio-estereoscopio (Asaeda & Wolanski 2002, Wolanski et al. 2003). Por medio de observación en campo, los agregados recolectados comprendieron en su mayoría un diámetro mayor a 1 cm, lo cual es una clara evidencia de la influencia de la escorrentía continental sobre la nieve marina en ambientes arrecifales (Fabricius et al. 2003). Las muestras fueron colectadas en zonas de arrecifes de corales mixtos entre 8 y 12 m de profundidad. Éstas se fijaron con gluteraldehído al 15% y se almacenaron en frío (4 °C aproximadamente) para su posterior análisis.
La determinación de la abundancia microbiana se midió mediante epifluorescencia para lo cual las muestras se pasaron por filtros Nucleopore de 0,2 um de diámetro de poro y 25 mm de diámetro. Como fluorocromo se empleó primulina, que tiñe el citoplasma y las membranas de los organismos en estudio sin enmascarar la auto fluorescencia de la clorofila a, lo cual facilita la diferenciación de microorganismos autótrofos, de heterótrofos. Si bien existen microorganismos que contienen clorofila y presentan también metabolismo heterótrofo, la metodología utilizada permitió conocer el metabolismo bacteriano, en cuanto a la fuente de carbono utilizada, en el momento y las condiciones de muestreo.
El conteo de células se realizó utilizando un microscopio de epifluorescencia Olympus B-Max- 60 con filtros de excitación para luz azul y ultravioleta. La fracción total de microorganismos se determinó con luz ultravioleta, bajo la cual la primulina tiene su máximo de excitación y las células coloreadas se observan con un intenso brillo de color azul blancuzco. Al cambiar el filtro de excitación hacia la luz azul, se identificaron las células autótrofas por la observación de una coloración roja típica de la clorofila a, mientras que la abundancia de heterótrofas se obtuvo a partir de la resta de autótrofas del total de células observadas (Caron 1983). Para el conteo de células se eligieron al azar treinta campos en cada filtro de manera que se observaran más de 200 microorganismos. Se tuvieron en cuenta las células del picoplancton y del nanoplancton con tamaños entre 0,2 y 20 um, para lo que se utilizó una retícula ocular, en la cual para el objetivo de 100 X, cada cuadro equivale a 10 um. La densidad celular (células mL-1) se determinó teniendo en cuenta el número promedio de células por campo, las veces que cabe el área de la retícula en el área de filtración, el inverso del factor de dilución y el volumen filtrado de muestra (Caron 1983). Teniendo en cuenta que no se extrajeron réplicas de las muestras y, que el presente trabajo es un estudio inicial (primer reporte), no fue posible realizar comparaciones estadísticas entre los sitios. Por lo anterior, a partir de los datos obtenidos se analizó gráficamente la densidad de microorganismos discriminándolos según fueran autótrofos o heterótrofos.
Resultados y discusión
Se registró la presencia de distintos tipos de microorganismos asociados a la nieve marina lodosa (euglenas, diatomeas, bacterias, hongos, cianobacterias, entre otros). Por una parte los microorganismos encontrados correspondieron a bacterias de vida libre, euglenas y, en algunos casos hongos y cianobacterias que tienden a no adherirse a los agregados. Estudios han demostrado que la nieve marina está habitada por una comunidad de organismos fitoplanctónicos, con densidades en algunos casos superiores a las de microorganismos de vida libre, como consecuencia de la mayor disponibilidad de nutrientes orgánicos e inorgánicos en comparación con el agua circundante (Strauss 2005, Azam & Malfatti 2007, Yam 2007).
Asociadas a los agregados de nieve marina también se observaron "bacterias especialistas en partículas", término atribuido a bacterias, que por sus características metabólicas cuentan con la maquinaria enzimática requerida para asimilar la materia orgánica y los nutrientes disponibles en estos agregados (Grossart et al. 2003). Estas bacterias se presentaron agrupadas formando largas cadenas. Por último se registró la presencia de bacterias generalistas, caracterizadas por estar tanto asociadas a los agregados de nieve marina como en el agua circundante, lo cual al parecer depende de la disponibilidad de nutrientes, así como de las condiciones particulares del ambiente marino (Grossart et al. 2006).
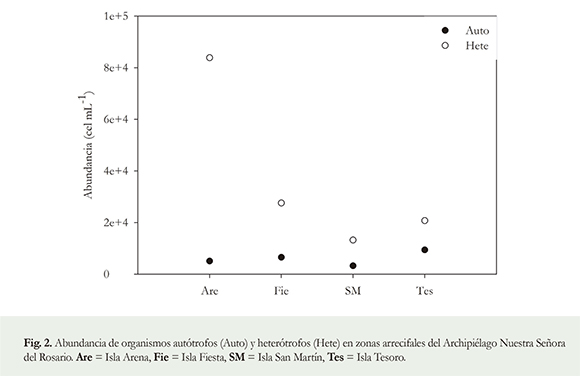
Las bacterias heterótrofas exhibieron mayores abundancias promedio (± SE) que las autótrofas, i.e. 3.63 x 104 ± 1.6 x 104 y 6 x 103± 1.3 x 103 células mL-1, respectivamente. La mayor densidad de heterótrofos, favorece la agregación de partículas de nieve marina y, de este modo, se obtienen partículas de mayor tamaño (Bidle & Fletcher 1995). El mayor tamaño de los agregados, incrementa la velocidad de hundimiento (Asaeda & Wolanski 2002), depositando la nieve marina lodosa con una alta carga de bacterias heterótrofas en los arrecifes (Fabricius & Wolanski 2000, Fabricius et al. 2003). Esto resulta perjudicial para los organismos sésiles como los corales ya que su remoción se dificulta aún más, generando mortalidad parcial en colonias adultas y total en reclutas (Fabricius et al. 2003, Wolanski et al. 2003). También se infiere que dentro del grupo de bacterias pueden existir patógenos potenciales que se constituyen en vectores de enfermedades coralinas, estas bacterias alóctonas crean condiciones anóxicas y producen metabolitos tóxicos; que generan efectos letales para los arrecifes de coral (Wolanski et al. 2003, Fabricius et al. 2005, Otero 2009).
La abundancia de microorganismos presentó variaciones entre las estaciones de muestreo. Isla Arena evidenció el mayor valor (8.9 x 104 células mL-1), mientras que Isla San Martín e Isla Tesoro registraron las menores abundancias con valores de 1.6 x 104 y 3 x 104 células mL-1, respectivamente. Las densidades obtenidas son análogas a otras reportadas en arrecifes del Caribe (Tabla 1) en zonas influenciadas por aguas continentales con abundante carga de material particulado (Bidle & Fletcher 1995, Jürgens et al. 2000, Kiorboe 2001, Garneau et al. 2009). En este estudio, Isla Arena es el sitio más cercano a la fuente de escorrentía, mientras que San Martín y Tesoro, corresponden a zonas más retiradas. Los sitios cercanos registraron mayores abundancias de microorganismos heterótrofos, mientras que aquellos más distantes presentaron un mayor número de células autótrofas (Figura 2). Lo anterior puede indicar la presencia de materia orgánica que al agregarse a la nieve marina lodosa, estaría promoviendo el establecimiento y crecimiento de bacterias heterótrofas que asimilan los nutrientes (Alldredge et al. 1998, Strauss 2005, Garneau et al. 2009), por lo que la densidad de este tipo de microorganismos podría ser empleada como bioindicador de la influencia de la escorrentía continental que aporta material particulado a los arrecifes. Con lo anterior, además de inferir que en las zonas arrecifales estudiadas existe alta disponibilidad de materia orgánica por aportes de escorrentía, también se cree que puede presentarse un gradiente ambiental que refleja una mayor influencia de la nieve marina lodosa con alta carga bacteriana heterótrofa en zonas arrecifales más cercanas a los aportes del Canal del Dique desde la Bahía de Barbacoas.

Conclusión
Con este estudio se reporta la presencia de nieve marina lodosa en arrecifes de la zona de Cartagena, Caribe colombiano, con una alta carga de microorganismos, especialmente heterótrofos. Se requieren investigaciones dirigidas a determinar las concentraciones de nieve marina lodosa en los arrecifes, así como a la composición de la comunidad de bacterias, y la posible virulencia de las bacterias heterótrofas de la nieve marina lodosa hacia los corales hermatípicos de los arrecifes. Por otra parte son fundamentales estudios relacionados con la determinación de la fracción mineral y la posible relación entre la composición mineralógica de los agregados de nieve marina y las bacterias asociadas.
Agradecimientos
Los autores agradecen a la Universidad Jorge Tadeo Lozano, a Beatriz Ossa de la Fundación Diving Planet y al Parque Nacional Corales del Rosario y San Bernardo por su apoyo económico y logístico en las diferentes fases de la investigación. También a los funcionarios del Parque Esteban Zarza, Diego Duque y Ameth Vargas, por su ayuda en la extracción de las muestras.
Conflicto de intereses
Los autores declaramos no tener conflicto de intereses.
Referencias
Alldredge AL, Cole JJ, Caron DA (1986) Production of heterotrophic bacteria inhabiting macroscopic organic aggregates (marine snow) from surfaces waters. Limnology and Oceanography 31(1): 68-78 [ Links ]
Alldredge AL, Silver MW (1988) Characteristics, dynamics and significance of marine snow. Progress in Oceanography 20(1):41-82 doi: 10.1016/j.bbr.2011.03.031 [ Links ]
Alldredge AL, Granata TC, Gotschalk CC, Dickey TD (1990) The physical strength of marine snow and its implications for particle disaggregation in the ocean. Limnology and Oceanography 35(7):1415-1428 [ Links ]
Alldredge AL, Passow U, Haddock HD (1998) The characteristics and transparent exopolymer particle (TEP) content of marine snow formed from thecate dinoflagellates. Journal of Plankton Research 20(3):393-406 doi: 10.1093/plankt/20.3.393 [ Links ]
Alvarado EM, Pizarro V, Sarmiento A (2011) Formaciones arrecifales. En: Zarza E (ed) El entorno ambiental del Parque Nacional Natural Corales del Rosario y de San Bernardo. Parques Nacionales Naturales de Colombia. Dirección Territorial Caribe, Cartagena de Indias, Colombia, pp 109-123 [ Links ]
Asaeda T, Wolanski E (2002) Settling of muddy marine snow. Wetlands Ecology and Management 10(4): 283-287 doi: 10.1023/A:1020348918829 [ Links ]
Azam F, Malfatti F (2007) Microbial structuring of marine ecosystems. Nature Reviews Microbiology. 5: 782-791 doi: 10.1038/nrmicro1747 [ Links ]
Bidle KD, Fletcher M (1995) Comparison of free-living and particle-associated bacterial communities in the Chesapeake Bay by stable low-molecular- weight RNA analysis. Applied and Environmental Microbiology 61(3):944-952 [ Links ]
Caron DA (1983) Technique for enumeration of heterotrophic and phototrophic nanoplankton, using epifluorescence microscopy, and comparison with other procedures. Applied and Environmental Microbiology 46(2):491-498 [ Links ]
Fabricius KE, Wolanski E (2000) Rapid smothering of coral reef organisms by muddy marine snow. Estuarine, Coastal and Shelf Science 50(1):115-120 doi: 10.1016/j.bbr.2011.03.031 [ Links ]
Fabricius KE, Wild C, Wolanski E, Abele D (2003) Effects of transparent exopolymer particles and muddy terrigenous sediments on the survival of hard coral recruits. Estuarine, Coastal and Shelf Science 56:1-9 doi: 10.1016/j.bbr.2011.03.031 [ Links ]
Fabricius KE (2005) Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Marine Pollution Bulletin 50(2):125-146 doi: 10.1016/j.bbr.2011.03.031 [ Links ]
Garneau ME, Vicent WF, Terrado R, Lovejoy C (2009) Importance of particle-associated bacterial heterotrophy in a coastal Arctic ecosystem. Journal of Marine Systems 75(1-2):185-197 doi: 10.1016/j.bbr.2011.03.031 [ Links ]
Garzón J, Díaz JM (2003) The Caribbean coral reefs of Colombia. In: Jorge Cortés (ed) Latin American coral reefs. Elsevier Science B.V, Amsterdam, The Netherlands, pp 275-301 [ Links ]
Grossart HP, Kiorbe T, Tang KW, Ploug H (2003) Bacterial colonization of particles: growth and interactions. Applied and Environmental Microbiology. 69(6):3500-3509 doi: 10.1128/AEM.69.6.3500-3509.200 [ Links ]
Grossart HP, Kiorbe T, Tang KW, Allgaier M, Yam EM, Ploug H (2006) Interactions between marine snow and heterotrophic bacteria: aggregate formation and microbial dynamics. Aquatic Microbial Ecology 42:19-26 [ Links ]
Henao-Castro HA (2013) Efectos de los aportes del Canal del Dique sobre el reclutamiento de especies de coral en los arrecifes del Archipiélago Nuestra Señora del Rosario, área marina protegida. Tesis de Maestría. Facultad de Ciencias Naturales e Ingeniería, Universidad Jorge Tadeo Lozano, Colombia [ Links ]
Jürgens K, Gasol JM, Vaqué D (2000) Bacteria- flagellate coupling in microcosm experiments in the Central Atlantic Ocean. Journal of Experimental Marine Biology and Ecology 245(1):127-147 doi:10.1016/j.bbr.2011.03.031 [ Links ]
Kaltenbock E, Herndl GJ (1992) Ecology of amorphous aggregations (marine snow) in the Northern Adriatic Sea. IV. Dissolved nutrients and the autotrophic community associated with marine snow. Marine Ecology Progress Series 87:147-159 [ Links ]
Kiorbe T (2001) Formation and fate of marine snow: small-scale processes with large-scale implications. Scientia Marina 65(2):57-71 doi: 10.3989/scimar.2001.65s257 [ Links ]
Luna GM, Manini E, Danovaro R (2002) Large fraction of dead and inactive bacteria in coastal marine sediments: comparison of protocols for determination and ecological significance. Applied and Environmental Microbiology 68(7):3509-3513 doi: 10.1128/AEM.68.7.3509-3513.2002 [ Links ]
Lyons MM, Ward JE, Gaff H, Hicks RE, Drake JM, Dobbs FC (2010) Theory of island biogeography on a microscopic scale: organic aggregates as islands for aquatic pathogens. Aquatic Microbial Ecology 60:1-13 doi: 10.3354/ame01417 [ Links ]
Lyons MM, Lau YT, Carden WE, Ward JE, Roberts SB, Smolowitz R, et al. (2007) Characteristics of marine aggregates in shallow-water ecosystems: implications for disease ecology. EcoHealth 4(4):406-420 doi: 10.1007/s10393-007-0134-0 [ Links ]
Navas R Rodríguez Y (2008) Estado de los arrecifes coralinos. En. INVEMAR (ed) Informe del Estado de los Ambientes y Recursos Marinos y Costeros en Colombia año 2007. Serie de Publicaciones Periódicas, Santa Marta, Colombia, pp 53-57 [ Links ]
Navas R, Agudelo DL, Rodríguez A, Reyes MC, Garzón J (2010) Coral diseases and bleaching on Colombian Caribbean coral reefs. Revista de Biología Tropical 58(1):95-106 [ Links ]
Otero E (2009) Spatial and temporal patterns of water quality indicators in reef systems of southwestern Puerto Rico. Caribbean Journal of Science 45(2-3):168-180 [ Links ]
Restrepo JD, Kjerfve B, Hermelin M, Restrepo JC (2006) Factors controlling sediment yield in a major South American drainage basin: the Magdalena River, Colombia. Journal of Hydrology 316(1-4):213-232 doi: 10.1016/j.bbr.2011.03.031 [ Links ]
Restrepo JD, Alvarado EM (2011) Assessing major environmental issues in the Caribbean and Pacific coasts of Colombia, South America: an overview of fluvial fluxes, coral reef degradation, and mangrove ecosystems impacted by river diversion. In: Wolanski E, McLusky D (ed) Treatise on Estuarine and Coastal Science. Waltham: Academic Press, USA, pp 289-314 doi: 10.1016/B978-0-12-374711-2.01117-7 [ Links ]
Riebesell U (1992) The formation of large marine snows and its sustained residence in surface waters. Limnology and Oceanography 37(1):63-76 [ Links ]
Rodríguez A, Reyes MC, Zea S, Navas R, Garzón J, et al. (2010) Recent dynamics and condition of coral reefs in the Colombian Caribbean. Revista de Biología Tropical 58(1):107-131 [ Links ]
Simon M, Grossart HP, Schweitzer B, Ploug H (2002) Microbial ecology of organic aggregates in aquatic ecosystems. Aquatic Microbial Ecology 28(2):175-211 [ Links ]
Stoddart DR (1969) Ecology and morphology of recent coral reefs. Biological Reviews 44(4):433-498 doi: 10.1111/j.1469-185X.1969.tb00609.x [ Links ]
Strauss EA (2005) Microbiology of lotic aggregates and biofilms. Water Encyclopedia, pp 305-309 doi:10.1002/047147844X.sw1611 [ Links ]
Turley C (2002) The importance of "marine snow". Microbiology Today 29:177-179 [ Links ]
Vega J, Navas R, Gómez K, López T, Duque D (2011) Estado de los ecosistemas marinos y costeros. En: INVEMAR (ed) Informe del estado de los ambientes y recursos marinos y costeros en Colombia: Año 2010. Serie de Publicaciones Periódicas No. 8, Santa Marta, Colombia, pp 88-115 [ Links ]
Wolanski E, Richmond R, McCook L, Sweatman H (2003) Mud Marine Snow and Coral Reefs. American Scientist 91(1):44-51 [ Links ]
Wolanski E, Fabricius KE, Cooper TF, Humphrey C (2008) Wet season fine sediment dynamics on the inner shelf of the Great Barrier Reef. Estuarine, Coastal and Shelf Science 77(4):755-762 doi: 10.1016/j.bbr.2011.03.031 [ Links ]
Yam E (2007) The role of bacteria- particle interactions in marine snow dynamics. Thesis of Master. School of Marine Science, Faculty of Science, The College of William and Mary, Williamsburg, Virginia [ Links ]
Ziervogel K, Steen AD, Arnosti C (2010) Changes in the spectrum and rates of extracellular enzyme activities in seawater following aggregate formation. Biogeosciences 7:1007-1015 doi: 10.5194/bg-7-1007-2010 [ Links ]
