











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
In plants, the disequilibrium of the terrestrial carbon cycle (excess CO2) causes biological alterations such as increased biomass and increased production of carboxylic acids. These carboxylic acids are secreted through the roots, as a detoxification system, to the soil [1-4]. Oxalic acid and its calcium salt, which contain dicarboxylate ions, are widely distributed in animals, plants, and fungi and other microorganisms [5]. The accumulation of calcium oxalate has been reported in more than 215 families of angiosperms and gymnosperms, and occurs in approximately 1 000 genera [6], including the Oxalidaceae family [6-11].
The oxalate-carbonate pathway (OCP) is a biogeochemical natural process for trapping atmospheric carbon and includes interactions between plants and microorganisms [12]. Many taxonomic groups of bacteria can use oxalate as a sole carbon source via the degradation of calcium oxalate. Consequently, this degradation leads to the precipitation of CaCO3 [13]. Oxalotrophy is widespread and is exhibited by gram-negative (Alpha-, Beta- and Gammaproteobacteria) and gram-positive bacteria (Firmicutes and Actinobacteria). The role of oxalotrophic bacteria as major calcium oxalate oxidizers was recently demonstrated [12]. Conventional culture methods reveal a diverse assemblage of oxalotrophic bacteria living near the rhizospheres of Terminalia oblonga, Terminalia bellirica, and Milicia excelsa in tropical forests of Bolivia, India, and Cameroon, respectively [14]. In the intertropical zone, the OCP observed underneath certain tropical trees like iroko (Milicia excelsa) is surprisingly well developed. By photosynthesis, the trees accumulate large quantities of organic salts, mainly in the form of calcium oxalate, which is a sparingly soluble salt (solubility product constant (Ksp) of 1.6 x 10-9 at 25 °C for the monohydrate), forming crystals in the plant tissues. The thermodynamic solubility product of calcium oxalate monohydrate (COM) at 25 °C is 1.6 x 10-9, that of calcium oxalate dihydrate (COD) is 3.7 x 10-9 and that of calcium oxalate trihydrate (COT) is 5.0 x 10-9. These values are associated with relative solubility, which indicates that the solubility of calcium oxalate hydrates decreases in the order COM > COD > COT [15, 16].
Calcium oxalate is consumed by bacteria present in the soil, which convert calcium oxalate to calcium carbonate via a biomineralization process in combination with other factors (pH, transformation of oxalate into bicarbonate, and equilibrium reactions and constants governing the dissolution of CO2 in aqueous media (CO3 2-, HCO3-, and H2CO3)) [17-19] (fig. 1). This p henomenon is a n et c arbon t rap in w hich the i nitial CO2 is atmospheric. The mainly acidic, non-calcareous soils found in tropical countries represent a significant sink for atmospheric CO2: at the present rate of increase, one full-grown iroko tree may stabilize the concentration of CO2 in the air column over a 2 400 m2 surface [9,14].

These circumstances make the rhizosphere (not a region of definable size or shape, but instead, a gradient of chemical, biological and physical properties that change both radially and longitudinally along the root) a competitive environment where microorganisms must develop strategies and mechanisms to improve their establishment, proliferation, and survival. The accumulation of polyhydroxyalkanoates (PHAs) has been proposed as being one such strategy. This is because the accumulation and subsequent degradation of these compounds is used for carbon and energy storage, and these compounds are mainly synthesized in chains of four carbons (polyhydroxybutyrate - PHB) from a wide range of organic substances in conditions of carbon imbalance, compared to nitrogen or phosphorus [20-23]. Several investigations have reported that a high concentration of oxalate in the rhizosphere is metabolized via two known pathways that diverge as the glycolate pathway and a variant of the serine pathway to produce 3-P-glycerate [5], which subsequently produces acetyl-CoA, a metabolic molecule that yields PHA [22]. However, to our knowledge, no research has been focused on the study of this type of bacteria in the application of this metabolism in soil.
In this study, we screened the production of PHA in oxalotrophic bacterial isolates from the soil adhering to the root of plant species from the genus Oxalis growing in the high Andean zone of Colombia. The positive strains were identified by partial sequencing of the 16S rRNA gene. Additionally, we discuss the results of gas chromatography analysis for the biopolymeric synthesis.
Materials and methods
Description of sampling sites
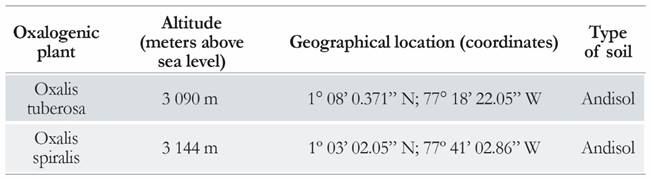
In Colombia, the species O. spiralis and O. tuberosa are found at 2700 and 3 800 m above sea level along the central and southern Andes, respectively. Considering this distribution, the plants were collected by taking into account the presence of seeds and the state of maturation of the individual [23]. The local temperature was between 12 °C and 18 °C, and the precipitation was between 1 000 mm and 4 000 mm per year. The samples (plant and soil near the roots) were labelled and transported to the PSO herbarium of the University of Narino to be dried and identified. The selected soil was used in previous studies, and the physicochemical parameters of this soil have been characterized extensively [24]. The geographical locations of sampling are given in Table 1.
Isolation of carbon-storing oxalotrophic bacteria
Enrichment was performed in Schlegel AB liquid medium [25] modified for the isolation of oxalotrophic bacteria with 16 g/L of insoluble calcium oxalate monohydrate, Ca(COO)2-H2O, as the sole carbon source (good energy efficiency [26]). The culture medium was adjusted to pH 7.0 and autoclaved for two hours. Ninety millilitres of medium in 250-mL flasks were inoculated with 10 g of dried soil and incubated at 30°C with constant shaking at 100 rpm for 12 hours. Serial dilutions up to 10-7 were prepared and 0.1 mL of the dilutions was spread on solid Schlegel AB medium with calcium oxalate. Fluorescence of the Nile red dye (0.01%, Sigma-Aldrich, USA) was measured as an indicator of PHA [27]. The plates were incubated at 30 °C for two weeks following the conditions proposed by Sahin and collaborators [28]. The bacterial colonies showing clear halos and fluorescence in Schlegel medium were considered positive for oxalate degradation and PHA lipid accumulation, respectively [29,30-32].
Oxalate consumption by oxalotrophic bacteria
Oxalate consumption by the bacteria was determined by the methodology proposed by Blackmore and Quayle [33]. Each bacterial isolate was inoculated into 250-mL flasks containing 60 mL of Schlegel culture broth adjusted to pH 5.0, at which the plant and the soil adhering to the root were collected. Cell suspensions were incubated at 30 °C with constant shaking at 100 rpm.
Every 24 hours the pH was measured, and the medium resulted, in general, slightly acidic [24].
The oxalate concentration of the medium after the incubation period was then determined by titration with 0.1 N standard sodium permanganate (KMnO4). Three millilitres of culture medium were extracted, and the calcium oxalate was solubilized by the addition of 1 mL of 2 N sulfuric acid (H2SO4) and titrated at 60 °C until a pink colour persisted for 30 s. Oxalate concentrations in the media (consumption) were calculated from the stoichiometry of reactants and products [33].
Evaluation of polyhydroxyalkanoate synthesis by bacterial isolates
The selected isolates were subjected to batch-type fermentation to produce PHA in liquid mineral medium (MM). The mineral medium, adapted from Rocha et al. [34], contained the following components (per litre): 1.5 g of KH2PO4; 3.54 g of Na2HPO4; 1.0 g of (NH4)2SO4; 0.2 g of MgSO4-7H2O; 0.01 g of CaCl22H2O; 0.06 g of (NH4)5[Fe(C6H4O7)2]-2H2O and 1.0 mL of trace elements solution. The trace elements solution contained the following components (g/L): H3BO3, 0.30; CoCl2-6H2O, 0.20; ZnSO4-7H2O, 0.1; MnCl2-4H2O, 0.03; NaMoO4-2H2O, 0.03; NiCl2-6H2O, 0.02 and CuSO4-5H20,0.01 [34]. The medium was supplemented with calcium oxalate as the sole carbon source; the source N:C ratio was 1:4. The fermentation experiments were conducted in triplicate for each bacterial isolate over 144 hours at 30 °C and 100 rpm. Then, 10 mL of a sample was centrifuged at 8 000 rpm for 10 minutes. The pellet was resuspended in 1 mL of 5% sodium hypochlorite and 10 mM EDTA. The suspension was incubated for 75 minutes at 60 °C, centrifuged at 8 000 rpm for 5 minutes, and washed repeatedly with 1 mL of distilled water, 1 mL of acetone and 1 mL of cold methanol; after each step, the sample was centrifuged at 8 000 rpm for 5 minutes [26,51]. Finally, the polymer was dried and weighed [35]. The values were reported as the means and the standard deviation (± SD) was calculated.
Characterization of the polymer by gas chromatography (GC)
The obtained polymer was dissolved in 1 mL of chloroform (HPLC grade) and 1 mL of derivatizing reagent (methanol/sulfuric acid; 85:15 v/v), and the reaction was performed at 90 °C for two hours. The mixture was cooled rapidly, and 1 mL of saline solution (0.1% NaCl) was added. Finally, the aqueous phase was removed, and the organic phase was treated with pure anhydrous sodium sulphate to remove any remaining water [36].
The gas chromatography (GC) analysis was carried out in the University of Narino laboratories in a Shimadzu GC-17A gas chromatograph equipped with a flame ionization detector (FID); helium AP was used as a carrier gas, and a capillary column of fused silica MDN-5 was used, which was 30 m in length, 0.25 mm in internal diameter and 0.25 microns in film thickness (Supelco). The injector and detector temperatures were set to an ionizing flame at 250 °C. The temperature program was 60 °C for 2 minutes, increasing 6 °C per minute to a maximum temperature of 220 °C [35,36]. The reference standard used was the copolymer poly-3-hydroxybutyrate-co-3-hydroxyvalerate.
Biochemical characterization of oxalotrophic bacteria producing neutral lipids.
Biochemical characterization was conducted as per Bergey’s manual [37]. These tests included the following: motility test, indole test, sulphide test, catalase activity test, oxidase activity test, the Voges-Proskauer test, and Simmons’ citrate test. The temperatures of incubation were 20 °C, 40 °C and 65 °C. The NaCl concentrations used were 2%, 4%, 7%, and 10%, and the pH values used were 4.0, 5.7, and 6.8. The fermentation process used glucose, sucrose, fructose, maltose, galactose, xylose, rhamnose, lactose, mannose, arabinose, and ribose [35].
DNA extraction
For DNA extraction, a protocol by Rivera and collaborators [38] was used with some modifications. Initially, bacterial isolates were inoculated into 25 mL of Luria-Bertani broth (LB) and incubated at 30 °C for 24 hours or until an approximate concentration of 108 CFU.mL-1 was reached. Subsequently, the cells were centrifuged at 7 800 rpm for 10 minutes; the cell pellet was dissolved in 2.4 mL of TE buffer (1 M) and then homogenized by vortexing for 5 seconds. Then, 62.5 μL of SDS 10% and 6.25 μL of proteinase K (20 mg/mL) were added to each tube prior to gentle homogenization. The mix was incubated in a water bath at 37 °C for 1 hour [38].
Subsequently, 300 μL of 5 M NaCl and 188 μL of CTAB (10%) were added. The solution was stirred in a water bath at 65 °C for 20 minutes. Then, an equal volume of chloroform:isoamyl alcohol (24:1) was added, and the solution was centrifuged at 8 000 rpm for 20 minutes. The supernatant was transferred to another tube and 3 μL of RNase (10 mg/mL) was added; the tube was incubated for 1 hour at 37 °C. Then, an equal volume of chloroform was added, and the mixture was homogenized, vortexed and centrifuged at 8 000 rpm for 15 minutes. The supernatant was transferred to another tube, and DNA was precipitated with 0.6 mL of isopropanol. The mix was centrifuged again at 8 000 rpm for 20 minutes, the pellet was washed with 100 mL of ethanol and finally centrifuged at 9 000 rpm for 5 minutes. The supernatant was discarded and the DNA was dried overnight at room temperature. The DNA was resuspended in TE (1 M) and stored at 4 °C for subsequent use. [35]
Quantification of DNA
The quality of the extracted DNA was checked on a 1% agarose gel run in a Fisher electrophoresis chamber at 60 volts for two hours. The molecular marker used was Lambda Hind III (Promega). The DNA was obtained by measuring the absorbance at 260 nm and 280 nm using a Genova Jenway spectrophotometer. Additionally, the A260/A280 ratios were calculated using the extinction coefficient for double stranded DNA (dsDNA), 0.0208 (μg/ml)-1 cm-1, and assuming that 1 OD at 260 nm equals 50 μg/mL DNA [39].
Amplification of the 16S rRNA gene
For the 16S rRNA gene amplification, universal primers 27F [AGAGTTTGATCMTGGCTCAG] [40].
And 1041R [CGGTGTGTACAAGACCC] [41] were used; the primers were synthesized by Integrated DNA Technologies.
The Polymerase Chain Reaction (PCR) was performed in a final volume of 50 μL. The master mix consisted of 10 μL of Green GoTaq® 5X buffer (Promega), 1 μL of 2.5 mM MgCl2 (Promega), 1 μL of a 2.5 mM dNTP mix, 1 μL of each primer (27F and 1041R) at a final concentration of 20 μM, 0.4 jL of DNA Polymerase GoTaq® (Promega) (5U/μL) and 2 μL of 100 ng/μL DNA target.
Amplification cycles were as follows: 2 minutes at 95 °C for initial denaturation followed by 30 cycles of 94 °C for 3 minutes, 5 °C for 1 minute, and 74 °C for 3 minutes, and a final elongation at 72 °C for 10 minutes. All reactions were performed in a thermocycler (MultiGene) in 0.2-mL Eppendorf tubes.
The DNA fragments amplified by PCR were visualized by agarose gel electrophoresis; the gel was run in 1X TBE buffer for 2 hours at 70 V and stained with 1% ethidium bromide (0.5 μg/mL). A 1-Kb molecular size marker was used (Promega). The amplified DNA fragments were visualized under UV light, and images were observed with a Gel Doc XR+ System controlled by Image LabTM software (Bio-Rad).
Amplicons were sent to Macrogen (Korea) for sequencing. Sequencing was performed using the method of Sanger et al. [42] using BigDye ABI3730XL with capillary electrophoresis. Using the ClustalW program and BLAST, the partial sequences of the 16S rRNA genes of the characterized wild bacterial isolates were aligned with further downstream nucleotide sequences obtained from the GenBank/Ribosomal Database Project/NCBI databases.
The 16S rRNA partial sequences were compared with the sequences available at the Ribosomal Database Project (RDP - http://rdp.cme.msu.edu/) [43] using the classifier tool to assign sequences to taxonomic levels. The Sequence Match tool was used to assign one representative of each cluster to the closest type-strain neighbour. Additionally, the same sequences were compared with 16S rRNA sequences deposited in the database of the National Centre for Biotechnology Information (NCBI GenBank) using the BLASTn tool.
Results
Identification and analysis of oxalotrophic strains
A total of 346 colonies with clear halos were obtained; no fungal microbiota was observed. Thirty oxalotrophic strains were isolated; colonies showing dissolution halos were selected as positive, and slants were prepared in fresh medium. Seven of the slants showed growth and fluorescence in Schlegel AB supplemented with calcium oxalate. This observation made possible the evaluation of the stained colonies that fluoresced under UV light. The isolates grew over 12 days, and all the isolates showed degradation halos around the colonies. The fluorescence was clear only on the fifteenth day. Six isolates were gram-positive and one was gram-negative. The selected colonies were transferred to plate count agar medium with Nile red and glucose. This medium had a greater imbalance in the carbon/nitrogen ratio; this imbalance was necessary for lipid accumulation [21,31].
The ideal growth temperatures were 30 °C to 42 °C. The ideal growth medium containing 4% and 7% of NaCl was used to test growth in the presence of different carbohydrates (Table 2).
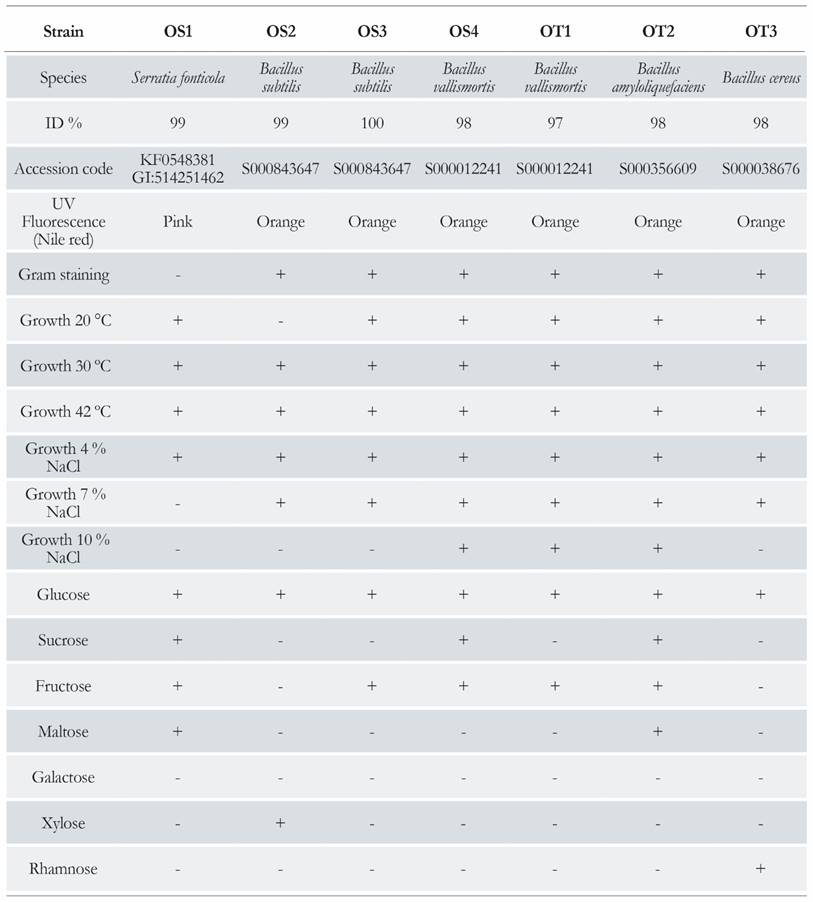
The fluorescent oxalotrophic strains were identified based on their 16S rRNA gene sequences. SeqMatch (Ribosomal Database Project) and BLAST (NCBI) were used to compare and classify the sequences. Six strains were identified as Bacillus and one as Serratia. A complete description of the identified strains is presented in Table 2.
Production of PHA
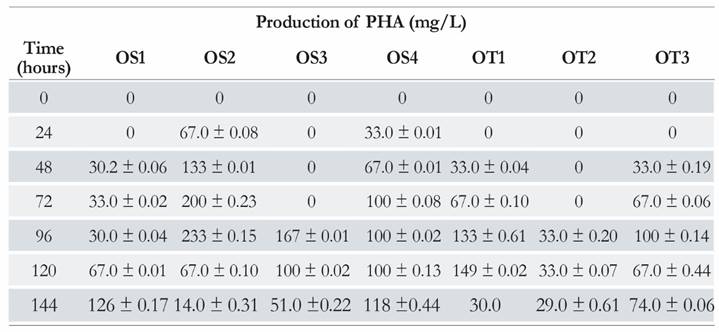
Regarding the production of PHA, it was observed that of the seven oxalotrophic bacteria, the isolates OS2, OS3, OT2 and OT3 showed high accumulation of the biopolymer at 96 h; i.e., 233 mg/L, 167 mg/L, 33 mg/L and 100 mg/L, respectively. The isolate OT1 showed high accumulation at 120 h (149 mg/L). Isolates OS1 and OS4 had the highest production of the biopolymer at 144 hours: 126 mg/L and 118 mg/L, respectively (Table 3).
Detection of PHA by gas chromatography
Gas chromatography (GC) was employed to perform monomer identification. The retention times of the methylated monomers of PHA synthesized by the isolate were identified by comparing the sample retention times with the retention times of the commercially available copolymer of ethyl 3-hydroxybutyrate and methyl 3-hydroxyvalerate (3HB-co-3HV) as standards; the methylated derivatives of the commercial copolymer formed two peaks with retention times of 4.5 minutes and 6.4 minutes. The strains OS1, OS2, OS3, OS4, OT1, OT2 and OT3 synthesized the monomer poly-3-hydroxybutyrate.
Discussion
Plant exudates containing oxalic acid provide an abundant source of carbon and nitrogen which are used by microorganisms in the rhizosphere, leading to an increase in the organic-matter content of the soil. Interestingly, a large fraction of the oxalotrophic bacterial isolates from Africa and Latin America correspond to the rhizospheric bacteria that may play a role in long-term soil pedogenic processes such as the OCP [14,44,45].
The two genera found in this study have been previously reported as oxalotrophic bacteria. Serratia marcescens WW4 (L7ZNH5), S. liquefaciens ATCC 27592 (S5EPA6), S. proteamaculans 568 (A8GE83), S. nematodiphila (A0A086GF10) and S.grimesii (A0A084YYH3) all have an oxalate decarboxylase gene that is necessary to catalyse oxalate. Some species of the Bacillus genus are reported to have oxalate-metabolizing activity in soil from the sagebrush steppe [46] and in tropical soils [14]. The ecological role of this genus is important because bacteria of this genus have been isolated from rhizospheric soil of different regions [5,47]. It is known that Serratia quinovorans and S. liquefaciens provide substances for plant growth and help with nitrogen fixation [47,48]. In addition, these bacteria have antifungal properties and act as insect pathogens; this would probably explain why the presence of fungi was not observed in our study. Moreover, it has been shown that Bacillus strains produce substances that promote plant growth, including gibberellins, indole acetic acid [49], and enzymes that act on phytic acid (phytases) [50].
Oxalotrophic bacteria can be ‘generalists’, i.e., when they can metabolize other substrates, or ‘specialists’, i.e., when they use oxalate as the sole carbon and energy source [51,52]. The bacteria in this study are generalists because they can metabolize oxalate and other compounds such as glucose. Therefore, these bacteria can be improved for future applications (such as ecological and clinical applications and the production of pharmaceuticals (e.g., anti-stress agents, antioxidants, and immune stimulants) from fruit extracts) because they can produce PHA from oxalate or glucose [12,14,26,52].
The identified oxalotrophic bacteria produced PHA. Serratia fonticola exhibited accumulation of P(3HB), coinciding with a report [51]. Bacillus vallismortis, B. amyloliquefaciens, and B. cereus produce P(3HB), and B. subtilis produces the copolymer P(3HB-co-3HV). The Bacillus genus shows rapid growth, and the hydrolytic enzymes of this genus produce structural copolymers from countless sources of carbon [53,54]. These strains have potential for future use because there is extensive information about Bacillus PHA producers, with PHA yields ranging mainly from 11% to 69%. Meanwhile, given the characteristics and potential applications of these organisms, there are different favourable PHA products with a wide range of monomer compositions: PHB, P(3HBco-3HV), P(3HB-co-3HHx), P(3HB-co-4HB-co-3HHx), P(3HB-co-6HHx-co-3HHx), P(3HB-co-4HB), and P(3HB-co-3HV-co-4HB). The formation of these products depends on the substrate and the fermentation conditions [55-60]. The system can be optimized to produce good yields of the copolymer for industrial applications of PHA.
The PHA observed in these strains is an alternative carbon source when the oxalate is completely transformed into carbonate and calcium ions. The oxido-reduction of oxalate-carbonate occurs biochemically via bacterial activity [61]. The energy produced by oxalate forces the microbial metabolism to perform other functions such as regulating the redox potential of the cell and decreasing oxygen concentrations; this allows for a concurrent increase in cellular levels of NADP [62]. Oxalate requires a high activation energy, but the energy efficiency is quite high for catabolic metabolism (-328 kJ/e- pair [26] compared to a glucose yield of -287.3 kJ/e- pair [63]) because one mole of oxalate can yield two ATPs; the Krebs cycle via glycerate/succinate is reactivated. Oxalate is a carbon and energy source, and the synthesis of PHA can likely occur via assimilation of oxalate with other carbohydrate sources, such as glucose. It is thought that the glycolate or serine pathway form the 3 P-glycerate [5]; these are substances considered precursors of acetyl-CoA, which is a metabolic molecule that yields PHB [22]. Various factors are involved in the formation of PHA. It would be interesting to propose a biosynthetic pathway based on experimental knowledge of the oxalate-carbonate pathway.
In this study, seven PHA-producing bacteria are reported. Six of these bacteria belong to the genus Bacillus (B. subtilis with two strains; B. vallismortis with two strains; B. amyloliquefaciens with one strain; and B. cereus with one strain) and one belongs to the genus Serratia (S. fonticola). PHA production on an industrial scale is mostly conducted with gram-negative bacteria, such as Ralstonia eutropha and recombinant Escherichia coli, because these bacteria grow rapidly and allow a high accumulation of biopolymers; however, these bacteria contain endotoxins that generate a strong immunogenic reaction in humans. Therefore, the application of PHA obtained from gram-negative bacteria is mainly restricted to the biomedical field. Additionally, gram-negative bacteria require expensive substrates such as propionic acid and valeric acid for the synthesis of biopolymers. In this sense, gram-positive bacteria such as those reported in this study have potential for application because they do not produce endotoxins and can also use inexpensive substrates or waste substances and transform them into polymers of interest, such as PHA, significantly lowering the cost of obtaining the polymer [64,65]. In this regard, it is worth mentioning that strains of the genus Bacillus grow very fast and in a wide variety of substrates [65].
The bioprospection of PHA in different environments is important. The future applications of the PHA polymers are mostly dependent on the challenge of increasing the production levels to obtain polymers with desired properties in an economical fashion. PHAs have been shown to be good candidates for tissue engineering applications, included in biomedical devices and as controlled drug carriers; agricultural applications; and protein purification applications, as well as candidates for use as biofuels [66-68]. For applications of PHA, the current technology -the whole process from the beginning to the final step- needs improvement. This suggests the selection and development of bacterial strains that are capable of efficient consumption and transformation of various substrates into a range of PHAs with different properties at a high yield, with high productivity using high-performance fermentations and efficient extraction and purification methods to lower the price. The utilization of cheap, locally available substrates, such as agri-waste, and cultivation processes combining batch and fed-batch fermentations may result in the highest productivity compared to other reported methods. However, considering the controllable nature of chemostats, these bioreactors have the greatest potential to provide higher productivities. Therefore, several current research efforts are devoted to lowering the production cost and meeting the target pricing by developing more efficient fermentation routes and recovery processes using recombinant or wild-type organisms. This field of research requires further investigation to enhance productivity and lower production costs rendering the product more competitive. All efforts at the laboratory scale will need to be validated at a pilot scale for future industrial production. The challenges of scaling up might hinder the use of procedures and processes that have been proposed as promising. Therefore, it is expected that ongoing research worldwide will soon potentially lead to PHA production at an acceptable cost [29,65,69,70,71].
Conclusions
The identified bacteria were: Serratia fonticola, Bacillus amyloliquefaciens, Bacillus subtilis, Bacillus vallismortis, and Bacillus cereus. Increased oxalate capacity and the carbon storage mechanism are new characteristics that contribute to the understanding of the oxalate-carbonate pathway.
The identified species play an important role in the soil of Oxalis plants, as reported in previous studies. These bacteria are promoters of plant growth and exhibit high rates of oxalate degradation, and may be considered as potentially useful in acidic soils as an alternative to bioremediation processes.
The high capacity for oxalate bioconversion using carbon reserve mechanisms for the synthesis of biopolymers by the microbial community studied was confirmed.
Given the versatility of the bacterial metabolism, such as oxalotrophy, and the versatility of PHA polymers, there are various potential applications, and the demand for PHA-based devices will continue to grow especially given the continued research in environmental and medical applications.

