










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
versão impressa ISSN 0122-8706
Corpoica cienc. tecnol. agropecu. vol.14 no.2 Mosquera jul./dez. 2013
FISIOLOGÍA DE SEMILLAS
1 Ingeniera Agrónoma, Banacol, Apartadó, Antioquia, Colombia.
2 PhD. Genética y Fisiología de semillas. C.I. La Selva, Corpoica, Rionegro, Antioquia y Universidad Nacional de Colombia, Medellín. mlobo@corpoica.org.co
3 Ingeniero Agrónomo y de Sistemas, MSc. Docente Universidad Católica de Oriente, Rionegro, Antioquia.
4 PhD. Ecofisiología C.I. La Selva, Corpoica, Rionegro, Antioquia. cmedina@corpoica.org.co
Fecha de recepción: 23/04/2013. Fecha de aceptación: 31/05/2013
RESUMEN
Se estudió la germinación y latencia de las semillas de Vasconcellea cundinamarcensis y V. goudotiana, para desarrollar procedimientos promotores de emergencia de los embriones. La categorización de las semillas viables no germinables se realizó con el empleo del tetrazolio. La fermentación prolongada de las semillas, para remover la sarcotesta, afectó la viabilidad de éstas, en especial las de V. cundinamarcensis. Aplicaciones de KNO3 y AG3 promovieron una germinación significativamente mayor, en comparación con el testigo, en semillas fermentadas previamente durante 15 días de V. goudotiana, lo cual no ocurrió en las de V. cundinamarcensis. La aplicación de Saccharomyces cerevisiae durante 36 horas, para fermentación y liberación de la sarcotesta, indujo germinación en las dos especies, con una respuesta significativamente superior en V. goudotiana. La adición de AG3 y KNO3, luego del tratamiento anterior, promovió mayor emergencia en ambos taxa, en especial en V. goudotiana. La fermentación de las semillas con Saccharomyces cerevisiae, en combinación con AG3 y KNO3 produjo germinación en las dos especies, con énfasis en V. goudotiana al aplicar 1000 ppm de AG3 + KNO3 al 2,5% y en V. cundinamarcensis, al utilizar 600 ppm de AG3 + AG3 al 2,5%, sin diferencia significativa con la aplicación de 1000 ppm de AG3 + KNO3 al 1,5%; se logró mayor germinación en V. goudotiana en el tratamiento precitado, con el cual se obtuvo con confiabilidad estadística la máxima brotación de plántulas, en relación con el resto de tratamientos evaluados con la propia especie y en V. cundinamarcensis.
Palabras claves: brotación de plantas, categorización, emergencia de embriones, viabilidad de semillas.
ABSTRACT
Seed germination and dormancy of Vasconcellea cundinamarcensis and V. goudotiana were studied in order to develop procedures for seed embryo emergency. Characterization of viable dormant seeds was done with the tetrazolium test. Long periods of seed fermentation, for sarcotestal removal, affected the seed viability, mainly in V. cundinamarcensis. AG3 and KNO3 application promoted a significantly higher germination, in V. goudotiana, but not in V. cundinamarcensis, in relation to control seeds, fermented during 15 days. AG3 and KNO3 application promoted a significantly higher germination in V. goudotiana, but not in V. cundinamarcensis, in relation to control 15 days fermented seeds. Seed fermentation with Saccharomyces cerevisiae during 36 hours for sarcotest removal, promoted a significant higher germination than that of the control, fermented without enzyme application, mainly in V. goudotiana. Seed fermentation with S. cerevisiae during 36 hours application, for sarcotest removal, promoted a significant higher germination than that of the control, fermented without enzyme application, mainly in V. goudotiana. Seed germination, after S. cerevisiae fermentation and application of AG3 and KNO3, occurred in both species, being obtained the best germination in V. goudotiana, with application of 1000 ppm of GA3 + 2.5% KNO3, with statistical differences, related to the other germination applied treatments. The best germination results, without significant differences, in V. cundinamarcensis were obtained with the application of 600 ppm of GA3 + 2.5% KNO3 or 1000 ppm of GA3 + 1.5% KNO3, after the fermentation procedure with S. cerevisiae.
Key words: plant growth, embryo emergency, seed viability.
INTRODUCCIÓN
Las llamadas papayuelas de altura, chamburo, papaya de tierra fría, chihualcán, siglalón, chichuacacón, titiish, papaya de altura y bonete, entre otras (Badillo, 1971; Fouqué, 1973), género Vasconcellea, comprenden un conjunto de 21 especies potenciales (Kyndt et al., 2005), relegadas y/o subutilizadas. Estas, según Cadavid et al. (2003), no han adquirido importancia como cultivo y se las encuentra como plantas individuales o en grupos pequeños en finca; sin embargo, en Chile, la especie Vasconcellea cundinamarcensis (V. pubescens), conocida como papayuelo o papayo de montaña, es un cultivo de exportación, con un área de siembra cercana a las 225 hectáreas, representadas principalmente por explotaciones comerciales establecidas en la zona norte, entre los 30° y 33° de latitud sur, cuya producción se aprovecha en la industria de conservas, jugos y mermeladas (Carrasco et. al., 2009). En dicho país, los agricultores obtienen sus plantas a partir de semillas, derivadas de los individuos con mejores atributos agronómicos (Ávila, 2007).
El taxón Vasconcellea cundinamarcensis, de acuerdo con Quintanilla (1995), se encuentra desde Colombia hasta Bolivia en forma espontánea; esto indica la necesidad de colectar los recursos de esta especie y taxa relacionados y, su preservación en colecciones de campo, con duplicados por la vía de semilla, la cual conduce a tener una representación mayor de la variabilidad poblacional y, de recombinantes entre los individuos de cada conglomerado de plantas. Al respecto, la Unión para la Conservación de la Naturaleza y los Recursos Naturales (IUCN) ha introducido cinco especies de Vasconcellea en la Lista Roja de taxa en peligro de extinción, las cuales corresponden a V. horovitziana, V. omnilingua, V. palandensis, V. pulchra, V. sprucei (IUCN, 2003). Incluso, se ha indicado que la lista de especies de este género en peligro podría ser más grande (Kindt et al., 2005) y, que la amenaza con mayor impacto es la destrucción de los hábitats donde éstas se encuentran, como secuela de la deforestación y reconversión de los bosques a tierras de cultivo o pastizales (IUCN, 2003).
Tomando como referente la potencialidad de la papayuela y especies relacionadas del género Vasconcellea, y el peligro de pérdida en condiciones in situ, se conformó a través de acciones de colecta una colección que cuenta actualmente con 80 accesiones de Vasconcellea, de los taxa: V. cundinamarcensis (pubescens), V. goudotiana, V. cauliflora, V. sphaerocarpa, V. longiflora y V. sp. (sin identificación), con 40, 32, 4, 2, 1 y 1 poblaciones, respectivamente. Este conjunto poblacional se vinculó, al Sistema de Bancos de Germoplasma de la Nación Colombiana para la Alimentación y la Agricultura - SBGNCAA-, y se conservan ex situ, en campo, con duplicados de semilla almacenados con 10% de humedad en ellas, dentro de bolsas de aluminio herméticamente selladas, en neveras a 12 °C.
El conjunto poblacional preservado, por la última vía, comprende semillas con y sin procesos de recombinación, con otras accesiones, por lo cual la colección de simientes no sólo sirve para reponer pérdidas de los materiales mantenidos en campo, sino también para la obtención de plantas derivadas de polinización abierta, con otras poblaciones. Lo mencionado puede alimentar procesos de selección de individuos élite, mediante clonación, destinados a evaluación y oferta posterior a los productores.
Todo lo anterior, señala la necesidad de categorizar la germinación y la latencia de las semillas, para resiembras de la colección de campo y monitorear la viabilidad de las unidades de propagación sexual conservadas de las diversas especies, para lo cual se precisa determinar la presencia de latencia en éstas y el desarrollo de protocolos para monitorear su viabilidad y aumentar los materiales conservados en condiciones de baja temperatura, antes de que lleguen a niveles reducidos de viabilidad, lo cual puede ocasionar erosión genética en el conjunto poblacional, en mantenimiento.
MATERIALES Y MÉTODOS
Localización: el estudio se llevó a cabo en el Laboratorio de Semillas del Sistema de Bancos de Germoplasma de la Nación Colombiana para la Alimentación y la Agricultura -SBGNCAA-, localizado en el centro de investigación La Selva, de Corpoica, Rionegro, Antioquia, Colombia.
Material biológico: en la investigación se incluyeron semillas de la especie cultivada V. cundinamarcensis (sinónimo: V. pubescens), al igual que del taxón promisorio V. goudotiana. Éstas se obtuvieron a partir de una mezcla aleatoria, balanceada, de 4 accesiones por especie, de la colección de campo establecida en el C.I. La Selva, Rionegro, Antioquia, a 2120 msnm, con una temperatura promedio de 17 °C, perteneciente a la formación ecológica bosque húmedo montano bajo.
Extracción de las semillas: se realizó mediante fermentación, durante tiempos variables, los cuales se indican para cada uno de los estudios; luego, se lavaron y se colocaron en desecadores herméticos, con sílica gel como desecante, con una relación de peso 1:1 entre éste y la semilla, hasta lograr 10% de humedad interna, cifra con la cual se almacenan los taxa con simientes intermedias, desde la óptica de tolerancia a la desecación, como es el caso de las papayuelas.
Pruebas de germinación: se emplearon germinadores Seedburo, con control de temperatura, a 28,5 °C, con colocación de las semillas sobre papel filtro humedecido con agua destilada. El número de semillas por unidad experimental se incluye en cada uno de los estudios. Las evaluaciones se llevaron a cabo durante 60 días, tiempo que permite detectar rompimientos iniciales de latencia.
Estudios realizados
Germinación de semillas extraídas a partir de frutos con diferente estado de madurez: las semillas se extrajeron a partir de frutos con diferente porcentaje de maduración, de acuerdo con el cambio de coloración de verde a completamente amarillo, en su epidermis: 25%, 50%, 75% y 100%. Dichas semillas se sometieron a fermentación, durante 15 días, para liberarlas de la sarcotesta. El estudio se hizo con un diseño factorial, completamente al azar, 2 X 4, los cuales corresponden a 2 especies y 4 estados de maduración; se emplearon unidades experimentales de 50 semillas y 5 repeticiones por tratamiento.
Estandarización del protocolo de tetrazolio para las semillas de papayuela: se desarrolló el procedimiento para semillas de papayuela, dado que no se encontró en la literatura. Para ello, se emplearon simientes recién extraídas y otras almacenadas en las n+veras, para semilla intermedia, del Banco de Germoplasma, las cuales se expusieron al tetrazolio durante 24, 48 y 72 horas; como métodos de aplicación se evaluaron tres procedimientos: incisión de la sarcotesta, con un bisturí, sin dañar el embrión; remoción manual de ésta; e inmersión de las semillas en la solución del tetrazolio, procedimiento recomendado, por la Sociedad Internacional de Analistas de Semillas, para detectar su viabilidad (ISTA, 2007). La prueba se basa en la actividad de las hidrogenasas, que catalizan las reacciones respiratorias en las mitocondrias, lo que permite correlacionar la viabilidad de las semillas con los cambios de color en los tejidos vivos (Grzybowski et al., 2012). Posteriormente, se hizo una evaluación visual del grado de tinción de los embriones, para seleccionar el protocolo que brindara un resultado confiable, mediante la tonalidad lograda en éstos, luego de la aplicación del tetrazolio; con indicación de Tunes et al. (2009), de que éste permite estimar rápidamente la viabilidad de las semillas, incluyendo las latentes.
Categorización de la viabilidad en semillas de V. cundinamarcensis y V. goudotiana, empleando tetrazolio, para la detección de simientes viables latentes: en el estudio se incluyeron semillas de las dos especies extraídas de: frutos podridos, sobrefermentados -método empleado por agricultores locales- y, frutos maduros, con fermentación de las simientes durante 15 días. La evaluación se llevó a cabo con un arreglo factorial 2X2, en el cual los factores correspondieron a los dos taxa y al tipo de fermentación al cual se sometió la semilla, con unidades experimentales de 100 simientes y 5 repeticiones. Éstas se pusieron a germinar, de acuerdo con el procedimiento descrito, para evaluar la emergencia de embriones. Los resultados se transformaron a valores de arcoseno de la raíz cuadrada del porcentaje, para su procesamiento estadístico y las diferencias entre tratamientos se procesaron a través de la prueba de Duncan, con una probabilidad de error de 0,05. Las simientes se clasificaron como: viables, las cuales corresponden a las germinadas y latentes sin emergencia de embriones, de acuerdo con la tinción con el tetrazolio; y, no viables, aquellas con embrión muerto, por falta de tinción con el indicador o sin presencia de la estructura. Para lo anterior, las semillas no germinadas se sometieron a tinción con tetrazolio, para determinar aquellas viables no germinables, de acuerdo con el protocolo obtenido, a partir de los resultados del acápite anterior. Éstas se sumergieron en la solución de tetrazolio, 0,1% en 100 ml de agua, durante 24 horas en condiciones de oscuridad, con separación posterior de los embriones, para evaluación de la coloración adquirida por éstos y, categorización de los viables, correspondientes a aquellos que exhibían color rojo brillante, en su superficie.
Efecto del ácido giberélico -AG3- y nitrato de potasio -KNO3- como promotores de germinación, en semillas provenientes de frutos fermentados y sobrefermentados de V. cundinamarcensis y V. goudotiana: dada la baja germinación, obtenida en la prueba anterior, se evaluó el efecto de los promotores de germinación giberelina AG3 y nitrato de potasio KNO3, en semillas provenientes de frutos sobremadurados, lo cual produce fermentación excesiva, y otro conjunto extraídas de frutos maduros luego de fermentación, durante 15 días; con secado posterior de los dos tipos de simiente y su humedecimiento previo. Con las semillas se evaluaron tres tratamientos por especie: KNO3 al 1,5%; AG3 200 ppm; KNO3 al 1,5% + AG3 200 ppm y un testigo absoluto, en los dos taxa, sin aplicación de los promotores de germinación. La evaluación se llevó a cabo con un diseño factorial 2X4, lo cual comprende la combinación de las dos especies, V. cundinamarcensis, V. goudotiana y los tratamientos relacionados. La germinación se llevó a cabo en germinadores, marca Seedburo, con control de temperatura, en condiciones controladas de 28 a 30 °C, con 50 semillas por unidad experimental y 5 repeticiones.
Efecto de la fermentación previa de las semillas de V. cundinamarcensis y V. pubescens, con la levadura S. cerevisiae para facilitar la eliminación de la sarcotesta y empleo posterior de los promotores de germinación, ácido giberélico -AG3- y nitrato de potasio -KNO3- para remoción de latencia: dado que la fermentación realizada por agricultores, por tiempos prolongados, para liberar la semilla de la sarcotesta causa pudriciones que pueden afectar la semilla por aspectos fisiológicos o patológicos, como alternativa se evaluó la fermentación previa con la levadura S. cerevisiae para facilitar la eliminación manual de la estructura mencionada. Para ello, se disolvieron 26 gramos de levadura y 105 de azúcar, en un litro de agua tibia, la cual se expuso al sol durante 30 minutos previo a la aplicación a las semillas, con adición del agente emulsificante monoestearato de sorbitán. Éstas se sumergieron en el preparado para su fermentación, a temperatura de 33 °C, por espacio de 36 horas. Posteriormente, se evaluó la germinación con un diseño completamente al azar con arreglo factorial 2X5, correspondientes a las dos especies: V. cundinamarcensis, V. pubescens y a los tratamientos: con sarcotesta, fermentación con levadura; con sarcotesta, fermentación sin levadura; sin sarcotesta, fermentación con levadura; con sarcotesta, fermentación con levadura, aplicación de 200 ppm de AG3 y KNO3 al 1,5%; sin sarcotesta, fermentación con levadura, aplicación de 200 ppm de AG3 y KNO3 al 1,5%. Las unidades experimentales estuvieron integradas por 80 semillas con 5 repeticiones.
Germinación obtenida con semillas previamente fermentadas, con sarcotesta y sin sarcotesta, con la levadura S. cerevisiae y con aplicación de los promotores de emergencia de embriones, ácido giberélico, AG3 y nitrato de potasio, KNO3, en diversas concentraciones: el estudio buscó evaluar la germinación luego de la aplicación de la enzima Sacharamyces cerevisiae, con la cual se redujo el tiempo de fermentación y los daños causados por el proceso, con aplicación posterior de combinaciones de diferentes dosis de AG3 y KNO3. La semilla provino de una mezcla balanceada de 4 accesiones de cada una de las especies, V. cundinamarcensis y V. goudotiana, extraídas a partir de frutos en completa maduración y sometidos a fermentación, con la sarcotesta, en presencia de la levadura mencionada, durante 36 horas. Posteriormente, las simientes fermentadas se secaron hasta un contenido de humedad de 10%, con silica gel como absorbente de humedad, en cámaras herméticas Fisher Scientific; luego, se rehidrataron para la aplicación de los tratamientos, con los promotores. El estudio se llevó a cabo con un diseño factorial 2X7, con cinco repeticiones y unidades experimentales de 50 simientes. Los factores correspondieron a las dos especies y los tratamientos de aplicación de los promotores de germinación. En total se evaluaron 6 combinaciones de los estimuladores de la emergencia, más un testigo, correspondiente a semilla fermentada con la enzima.
RESULTADOS Y DISCUSIÓN
Germinación obtenida en semillas extraídas de frutos con diferente grado de maduración: no se obtuvo germinación en la semilla proveniente de frutos con diversos grados de maduración, referenciados por el desarrollo de pigmentos carotenoides en la cáscara, en las especies V. cundinamarcensis y V. goudotiana; la semilla se extrajo luego de fermentación durante 15 días, procedimiento empleado por algunos agricultores. Lo precedente puede señalar una latencia profunda de las simientes o, un efecto nocivo del proceso prolongado de fermentación al que se sometieron para la remoción de la sarcotesta. En contexto con lo anterior, Baskin y Baskin (2001) indicaron que las semillas de muchas especies exhiben latencia en el estado de madurez completa de los frutos y que no presentan germinación, en un conjunto amplio de condiciones ambientales. Igualmente, en el género Vasconcellea, se han reportado efectos nocivos de la fermentación, durante períodos largos, con recomendación de fermentación de la semilla durante 24 horas (CAF, 1992), para posterior remoción manual de la sarcotesta (Scheldeman, 2002).
Protocolo de tetrazolio estandarizado para las semillas de V. cundinamarcensis y V. goudotiana: se encontró que la mejor tinción de la semilla con tetrazolio se obtenía luego de 24 horas de inmersión en la solución, al 1%, con buena penetración del producto, y que en las simientes con remoción manual de la sarcotesta se lograba una tinción nítida que permitía diferenciar claramente las viables. En contraste, en aquellas sometidas al indicador de viabilidad, sin remoción de la sarcotesta, no se produjo tinción alguna, con coloración leve en las que se realizaron incisiones con bisturí. Esto permitió seleccionar como procedimiento de evaluación de viabilidad, el tratamiento del colorante por 24 horas, a las unidades de propagación sexual, luego de remoción manual de la sarcotesta, lo cual brinda una tinción nítida y un indicador confiable de la viabilidad de las semillas. Con relación a la extracción de la sarcotesta, en forma previa a la aplicación del tetrazolio, Tokuhisa et al. (2006) indicaron que en la especie relacionada, Carica papaya, la sarcotesta -cubierta mucilaginosa que cubre externamente las semillas- podía ocasionar latencia física.
Viabilidad de las semillas de V. cundinamarcensis y V. goudotiana, validada a través de pruebas de germinación y del tetrazolio: los resultados porcentuales de germinación y latencia, obtenidos con semillas fermentadas y sobrefermentadas de las especies V. cundinamarcensis y V. goudotiana, con remoción de la sarcotesta, para la evaluación de la germinación y prueba posterior de viabilidad con tetrazolio se incluyen en la tabla 1. Como puede apreciarse, en el taxón V. cundinamarcensis no se obtuvo germinación alguna con las simientes extraídas mediante fermentación y sobrefermentación, en tanto que en V. goudotiana se obtuvo emergencia de embriones, 18,7%, con las semillas fermentadas, mas no en las sobrefermentadas. Lo precedente está en línea con las simientes no viables, las cuales superaron 70% en V. cundinamarcensis, sin diferencias entre los dos procesos de fermentación, con porcentajes significativamente superiores de semillas muertas, correspondientes a embriones no viables y a las carentes de embrión, con relación al taxón V. goudotiana.
El mayor porcentaje de semillas viables, no emergidas, detectadas por el protocolo del tetrazolio y las germinadas, ocurrió en los lotes de simiente de V. goudotiana, provenientes de fermentación, con valores significativamente superiores a la sobrefermentada de la especie y aquellas de semilla fermentada y sobrefermentada de V. cundinamarcensis. En concordancia con lo anterior, el menor porcentaje de simientes no viables se obtuvo, con confiabilidad estadística, en aquellas de V. goudotiana fermentadas, en relación con las de la propia especie sobrefermentada y, a aquellas de V. cundinamarcensis fermentadas y sobrefermentadas (tabla 1), lo cual indica una pérdida de la viabilidad, derivado de los procesos de fermentación y sobrefermentación.
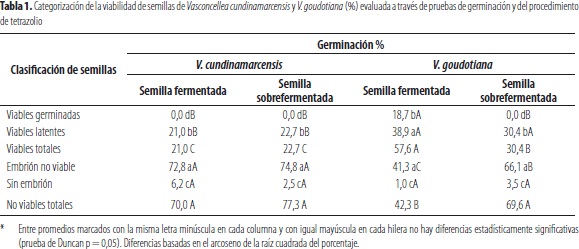
Los resultados obtenidos también indican que V. cundinamarcensis exhibió un porcentaje significativamente superior de inviabilidad en sus embriones, en comparación con V. goudotiana, lo cual no es atribuible a las simientes sin embrión, en las cuales no se detectaron diferencias significativas entre los tratamientos de fermentación y sobrefermentación, aplicados a las dos especies, con valores relativamente bajos que fluctuaron entre 1,0% y 6,2%. Por lo tanto, es presumible que los tratamientos previos de fermentación para la remoción de la sarcotesta hayan propiciado la pérdida de viabilidad de las simientes. Al respecto, aquellas con embrión no viable correspondieron, en alto grado, a pudrición causada por patógenos, con degradación de los tejidos, lo cual fue evidente en los dos taxa estudiados, durante la fermentación y sobrefermentación, para la remoción de la sarcotesta.
También se pudo inferir mayor afectación, derivada de los procesos de fermentación y sobrefermentación, en las simientes de V. cundinamarcensis que en aquellas de V. goudotiana, por la ausencia de germinación del primero en los dos procesos, lo cual sí ocurrió en la última especie, en la semilla sometida al tratamiento de menor duración. La ausencia de germinación de embriones viables, luego de los dos tiempos de fermentación para la eliminación de la sarcotesta, podría señalar inhibición de ésta por compuestos fenólicos presentes en la esclerotesta en mayor contenido que en la sarcotesta, considerados como inhibidores de la germinación por Tokuhisa et al. (2007) en Carica papaya; taxón con estrecha relación genética con las especies del género Vasconcellea, las cuales fueron incluidas, inicialmente, en forma conjunta con la papaya, en el género Carica (Badillo, 2000, 2001).
En contexto con lo anterior, Desai (2004) señaló que la emergencia de los embriones tiene dos componentes: uno genético y otro ambiental. Esto aplica a las unidades de propagación sexual de las papayuelas, ya que éstas provienen de polinización abierta, con presencia de flores unisexuales en diferentes plantas o en el mismo individuo; sobre lo cual, Scheldeman (2002) indicó que en el caso de V. cundinamarcensis existían plantas monoicas o dioicas, con presencia de flores macho, en ciertos períodos. Lo precedente, unido a la alogamia de la especies de Vasconcellea, señala constituciones genéticas diversas de las semillas de un mismo individuo e, incluso, de las plantas de cada accesión, lo que puede conducir a respuestas de germinación diferentes bajo condiciones similares de incubación. Adicionalmente, se ha reportado hibridación entre especies de Vasconcellea (Van Droogenbroeck et al., 2004), lo cual puntualiza, aún más, la variabilidad interna de las accesiones por posible introgresión espontánea interespecífica, en las poblaciones colectadas, en conservación; diversidad que debe estar relacionada con la latencia y la germinación obtenidas.
Aplicación de ácido giberélico y nitrato de potasio como promotores de germinación en semillas provenientes de frutos fermentados y sobrefermentados de V. cundinamarcensis y V. goudotiana
La germinación fue nula o casi nula en el taxón V. cundinamarcensis, tanto en las semillas fermentadas, 15 días en el proceso, como en las sobrefermentadas, obtenidas a partir de frutos podridos, con emergencia de embriones, en todos los tratamientos, en las unidades de propagación sexual en el caso de la especie V. goudotiana (tabla 2). En ésta última se apreció mayor germinación en semillas fermentadas con relación a las sobrefermentadas, en todos los tratamientos, lo cual incluyo el testigo, sin aplicación de promotores de brotación de plántulas. En el último taxón, la mayor germinación se obtuvo con los tratamientos de fermentación, con adición de 200 ppm del promotor ácido giberélico (AG3) y, aquel en el cual se aplicó 200 ppm de AG3, complementados con nitrato de potasio (KNO3), al 1,5%, con valores de 52,4% y 60,8% de emergencia, sin diferencias estadísticamente significativas entre éstos. Los resultados obtenidos apuntan a que el lote empleado de simientes de V. cundinamarcensis exhibía una latencia profunda o, su viabilidad fue afectada por los procesos de fermentación. Al respecto, Alarcón et al. (1997) informaron que las simientes de V. cundinamarcensis recién extraídas mostraban baja germinación. Igualmente, Varela et al. (2011) afirmaron que las semillas del taxón relacionado Vasconcellea monoica exhibían poca uniformidad en su germinación incluso en condiciones favorables para que esto ocurriera.
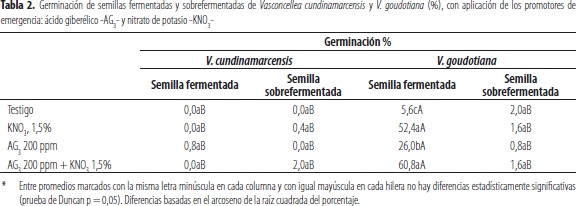
Con relación al efecto positivo de las giberelinas, AG3, y del nitrato de potasio KNO3, Gashi et al. (2012) señalaron que las primeras están más directamente implicadas en el control y promoción de la germinación que el otro compuesto. Esto se evidencia en el taxón V. goudotiana, en el cual no se encontraron diferencias significativas en la emergencia de plántulas entre los tratamientos derivados de la aplicación de AG3 y las de éste compuesto adicionado con KNO3. Al respecto, Finch-Savage y Leubuer-Metzger (2006) informaron que el ácido giberélico estimula la germinación por la vía de síntesis de amilasas; y Gashi et al. (2012) señalaron que el KNO3 es el compuesto químico más ampliamente utilizado para promover la germinación; a lo cual Alboresi et al. (2005) indican que los nitratos, tales como el KNO3, claramente estimulan la germinación de semillas latentes.
Respuesta germinativa desemillas de V. cundinamarcensis y V. goudotiana en combinaciones de tratamientos con y sin fermentación a través de la levadura S. cerevisiae, con remoción y no remoción de la sarcotesta y dos tratamientos adicionales de aplicación de AG3 200 ppm y KNO3 con y sin remoción de la sarcotesta: de acuerdo con los resultados, se detectó una respuesta germinativa superior en el taxón V. goudotiana, con relación a aquella de V. cundinamarcensis, con emergencia de embriones en todos los tratamientos, lo cual no ocurrió en el segundo (tabla 3). La mayor respuesta germinativa de V. goudotiana también fue evidente en los dos estudios presentados previamente (tablas 1 y 2). Al respecto, como se anotó, Alarcón et al. (1997) indicaron que la germinación en las semillas de V. cundinamarcensis es reducida en semillas recién extraídas. Esto también ha sido reportado en otras especies hermanas, como es el caso de V. monoica, de la que Varela et al. (2011) indicaron que en el taxón V. monoica, la emergencia de las semillas exhibía uniformidad baja, aun sometida a condiciones favorables, para la emergencia de los embriones.
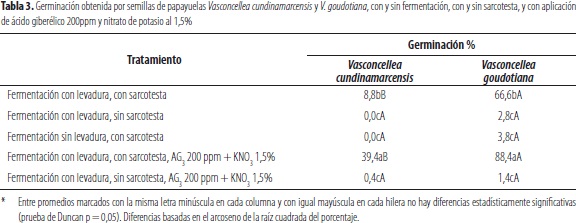
También pudo apreciarse un efecto positivo de la fermentación, con la aplicación de la levadura S. cerevisiae, la cual indujo alguna germinación en V. cundinamarcensis (8,8%), con valores significativamente superiores en V. goudotiana (66,6%), con una probabilidad de error de 0,05, en las semillas fermentadas con la sarcotesta (tabla 3). Los valores anteriores obtenidos con ambos taxa en las simientes fermentadas con levadura, fueron superados, con confiabilidad estadísticamente significativa de 0,95, en los tratamientos de fermentación con levadura en las semillas con sarcotesta, a las cuales se adicionó como promotores de germinación AG3 200 ppm y KNO3 al 1,5%. Al respecto, en las simientes fermentadas sin sarcotesta, lo cual se tradujo en germinaciones nulas o bajas, se puede apreciar incidencia de daños por microorganismos, sobre lo cual Cherry (1983) afirmó que el deterioro de las simientes se incrementa con condiciones favorables para los patógenos, los cuales desarrollan mecanismos inhibitorios a la emergencia de los embriones y, aun toxinas durante el proceso de germinación. Al respecto El-Saidy y El-Hai (2011) informaron que la levadura S. cerevisiae, aplicada a semillas de Arachis hypogaea, en almacenamiento, tiene efectos positivos en la germinación y las protege del ataque de patógenos en almacenamiento; esto podría indicar que su aplicación, en el proceso de fermentación, utilizado para remover la sarcotesta, actúa como agente controlador de los microorganismos patogénicos causantes de pudriciones en los procesos de fermentación.
Germinación lograda con semillas de V. cundinamarcensis y V. goudotiana, extraídas a través de fermentación durante tres días, por acción de la enzima S. cerevisiae, con aplicación posterior de diferentes combinaciones de dosis de AG3 y KNO3
En el estudio se presentó germinación en los dos taxa, en todos los tratamientos aplicados, incluso en el testigo, sin diferencias significativas entre éstas, con la sola aplicación de la levadura S. cerevisiae, en las cuales se obtuvo emergencia de embriones del orden de 24,4 y 25,0%, con V. cundinamarcensis y V. goudotiana, respectivamente (tabla 4). Lo anterior es atribuible a la reducción en el tiempo de fermentación a 36 horas y el efecto antipatogénico de la levadura, reportado por El- Saidy y El-Hai (2011).
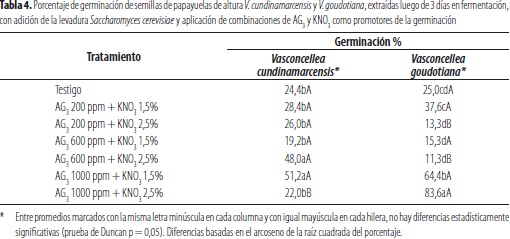
En los tratamientos aplicados luego de la fermentación con la levadura, se obtuvo la mayor germinación en el taxón V. cundinamarcensis, 51,2%, con la aplicación de 1000 ppm de AG3 más KNO3 al 1,5%, sin diferencias estadísticamente significativas con la obtenida al aplicar 600 ppm de AG3 y KNO3 al 2,5%, en los cuales se obtuvo una germinación de 51,20% y 48,0%, valores significativamente superiores a los obtenidos con los demás tratamientos evaluados con el taxón. Con la especie V. goudotiana se logró la máxima germinación, 83,6%, al tratar las simientes previamente fermentadas con S. cerevisiae, con 1000 ppm de AG3 y KNO3 al 2,5%, con diferencias estadísticamente significativas, con la respuesta obtenida con las demás combinaciones de los dos promotores de emergencia de embriones y, aun con todos los valores de germinación logrados con el taxón V. cundinamarcensis. Scheldeman (2002) señala que la aplicación previa de AG3 estimula la germinación, en concentraciones altas, y recomendó la dosis de 1000 ppm como tratamiento para acelerarla y maximizarla; también postuló que el nitrato de potasio igualmente induce emergencia de los embriones, lo cual está en concordancia con los resultados obtenidos.
Dado que la germinación de las especies de Vasconcellea es a menudo irregular, lenta y errática a lo largo del tiempo (Scheldeman, 2002), los resultados obtenidos deben ser confirmados con evaluaciones en semillas cosechadas en varias épocas y en accesiones diversas, para tener mayor confiabilidad, ya que el mismo autor informó diferencias entre poblaciones de la misma especie. Igualmente, reportó la mayor germinación en V. cundinamarcensis, en un conjunto de taxa del género Vasconcellea, en el cual no se incluyó V. goudotiana, taxón con respuestas germinativas superiores a las de V. cundinamarcensis, en el estudio presente.
Con relación a las simientes de la colección almacenadas a baja temperatura, en los monitoreos periódicos se requiere aplicar, aparte de la prueba de germinación, evaluaciones posteriores de viabilidad, con tetrazolio, para determinar la viabilidad real de cada una de ellas y así determinar la necesidad de colectar semillas a partir de la colección de campo en aquellas cuya germinación sea inferior al 80% de la inicial. Esto con el fin de mantener una representatividad adecuada de la variabilidad de los materiales sembrados, por la vía de simientes.
Igualmente, es recomendable la revalidación posterior de los resultados, con el fin de establecer protocolos confiables de remoción de la latencia, lo cual apoya la utilización del germoplasma con fines de investigación, así como desarrollo y posible entrega de cultivares superiores, seleccionados o procedentes de procesos de hibridación, para la implementación de posibles cultivos de papayuela. Ésta, con el Centro Primario de Diversidad en la Zona Andina, se siembra y exporta en Chile y, en el presente, está incluida, en Colombia, en el conjunto de especies relegadas, promisorias.
La colección de especies del género Vasconcellea puede servir para enriquecer la variabilidad genética de Carica papaya y la transferencia de atributos valiosos, a través de hibridación intergenérica de ésta, aspecto reportado por Vegas et al. (2003). Al respecto, Scheldeman (2002) informó que se han obtenido híbridos artificiales, intergenéricos, entre Vasconcellea y Carica papaya, para la transferencia de características como resistencia a enfermedades y tolerancia al frío. Estos exhiben barreras de incompatibilidad y aborto de las semillas (Magdalita et al., 1997), las cuales pueden superarse a través de rescate de embriones (Drew et al., 1998). Entre los atributos a transferir, a partir de los taxa de Vasconcellea; se han relacionado, entre otras, fragancias agradables, monoecia y cualidades ornamentales (Manshardt y Wenslaff, 1989), aparte de las anteriormente mencionadas.
CONCLUSIONES
No se obtuvo germinación alguna de semillas de las especies V. cundinamarcensis y V. goudotiana extraídas a partir de frutos en diferente grado de maduración, luego de fermentación, durante 15 días, para remoción de la sarcotesta. Esto puede indicar latencia profunda, por tratarse de taxa con un grado incipiente de domesticación y/o un efecto nocivo por la duración del proceso de fermentación.
El protocolo de tetrazolio, desarrollado y estandarizado para determinar latencia de las semillas de los taxa del estudio, permitió diferenciar las no germinables viables de las no viables. Esto apoyó las etapas de investigación para la elucidación de simientes latentes en aquellas que no emergieron. Igualmente, será una ayuda importante para los procesos de monitoreo de semillas vivas, con fines de regeneración oportuna y así evitar erosión genética de las semillas, en almacenamiento, como duplicado de seguridad de la colección de campo.
El taxón V. goudotiana exhibió respuesta germinativa (18,7%), a través de las semillas fermentadas durante 15 días, para remoción de la sarcotesta, mas no de aquellas provenientes de frutos podridos, sobrefermentados, procedimiento empleado por cultivadores de la especie; en V. cundinamarcensis con los dos tipos de simiente no hubo respuesta germinativa. Lo precedente está en línea con la pérdida de viabilidad, monitoreada por el tetrazolio, en el taxón, la cual fue superior a la de V. goudotiana. Esto señala que V. cundinamarcensis es más susceptible a pérdida de la viabilidad al someterse a los procesos de fermentación y sobrefermentación, lo cual puede atribuirse a aspectos fisiológicos de éste o a una mayor susceptibilidad a patógenos de las semillas durante el proceso.
El comportamiento germinativo del taxón V. cundinamarcensis también fue nulo o muy pobre, como secuela de la fermentación y sobrefermentación, y aplicación posterior de los promotores de germinación AG3 y AG3 + KNO3, con emergencia de embriones en todos los tratamientos de V. goudotiana, especialmente a partir de las simientes fermentadas. Esto reafirma el efecto negativo de los procedimientos de fermentación y sobrefermentación en el primer taxón, en el cual la germinación fue nula en los testigos, derivados de los dos procedimientos de fermentación y, casi nula al aplicar los promotores de emergencia de los embriones; lo que indica, en comparación con los resultados obtenidos con V. goudotiana, daño de las simientes por el proceso en sí y ataques de patógenos, lo cual se observó en el estudio.
De todas formas, es conveniente revaluar lo anterior, en la especie V. cundinamarcensis, con semillas de diversas accesiones, colectadas en épocas distintas, ya que en esta entidad biológica se han detectado comportamientos erráticos de la semilla, con relación al tiempo de colecta y almacenamiento, al igual que respuestas diversas entre accesiones.
La aplicación de la levadura S. cerevisiae para la fermentación de la sarcotesta indujo germinación per se en los dos taxa, con una respuesta significativamente superior en V. goudotiana, con germinaciones significativamente superiores al adicionar al proceso fermentativo del microorganismo, con la aplicación como promotores de emergencia de AG3, 200 ppm y KNO3 al 1,5%; los cuales tuvieron un efecto mayor en V. goudotiana. Lo anterior se confirmó en un estudio posterior, con obtención de germinación de alrededor de 25%, con la sola aplicación de la levadura S. cerevisiae, en las dos especies, y un efecto positivo al aplicar posteriormente AG3 y KNO3 como promotores de emergencia; en concentraciones de 600 ppm y al 2,5% y de 1000 ppm y al 1,5%, en el caso del taxón V. cundinamarcensis y de 1000 ppm y KNO3 al 2,5% en el de la especie V. goudotiana.
La combinación de la levadura para la fermentación de las semillas y los promotores de germinación AG3 y KNO3 en las concentraciones indicadas, para cada una de las dos especies, conforman un protocolo preliminar para la germinación, el cual es conveniente revalidar. De todas formas, como la latencia puede tener diferente profundidad, es recomendable, para el monitoreo de las semillas en conservación, realizar adicionalmente la prueba del tetrazolio para la determinación de la viabilidad real de las semillas, compuesta por la sumatoria del porcentaje de germinación y el de aquellas vivas latentes.
REFERENCIAS
Alarcón L, Benavides L, Parodi G. 1997. Efecto de la temperatura y período de almacenamiento sobre la germinación de la semilla de papayuela (Carica pubescens Lenné et Koch). Proceedings of the Interamerican Society for Tropical Horticulture. 41:242- 245. [ Links ]
Alboresi A, Gestin C, Leydecker MT, Bedu M, Meyer C, Truong HN. 2005. Nitrate a signal relieving seed dormancy in Arabidopsis. Plant Cell. Environ. 28(4):500-512. [ Links ]
Ávila P. 2007. Diversidad y estructura genética de Vasconcellea pubescens cultivada en Chile evaluada mediante marcadores ISSR. En: dspace.utalca.cil/bitstream/1950/6150/avila_figueroa.pdf [ Links ]
Badillo VM. 1971. Monografía de la familia Caricaceae. Universidad Central de Venezuela Maracay, Venezuela 221 p. [ Links ]
Badillo VM. 2000. Carica L. vs. Vasconcella St Hil. (Caricaceae) con la rehabilitación de este último. Ernstia 10(2):74-79. [ Links ]
Badillo VM. 2001. Nota correctiva Vasconcellea St Hill. (Caricaceae) y no Vasconcella (Caricaceae). Ernstia 11:75-76. [ Links ]
Baskin CC, Baskin JM. 2001. Seeds. Ecology biogeography and evolution of dormancy and germination. Academic Press. San Diego, California, USA. 627 p. [ Links ]
Cadavid A, Villegas E, Medina CI, Lobo M, Reyes C. 2003. Caracterización morfológica de caricáceas de altura. En: Memorias IV Seminario Nacional de Frutales de Clima Frío Moderado. Medellín, Colombia CDTF UPB Corpoica. p. 55- 60. [ Links ]
CAF. 1992. Manual técnico del cultivo de chamburo. Centro Agrícola de Quito Corporación Andino de Fomento. Quito, Ecuador 36 p. [ Links ]
Carrasco B, Ávila P, Pérez-Díaz J, Muñoz P, García R, Lavandero B, Zuita-Silva A, Retamales JB, Caligari PDS. 2009. Genetic Structure of higlland papayas Vasconcellea pubescens (Lennéet C. Koch) Badillo) cultivated along a geographic gradient in Chile as revealed by Inter Simple Sequence Repeats (ISRR). Genetic Resources and Crop Evolution 56(3):331-337. [ Links ]
Cherry JP. 1983. Protein degradation during seed deterioration. Phytopathology. 73(2):317-321. [ Links ]
Desai B. 2004. Seeds Handbook. Biology production processing and storage. 2nd edition. (Marcel Dekker). 787 p. [ Links ]
Drew RA, O'Brien CM, Magdalita PM. 1998. Development of Carica interspecific hybrids. Acta Horticulturae 461:285-291. [ Links ]
El-Saidy AEA, El-Hai KMA. 2011. Alleviation of seed peanut deterioration during storage using biotic and abiotic agents. Research Journal of Seed Science 4(2):64-81. [ Links ]
Finch-Savage WE, Leubuer-Metzger G. 2006. Seed dormancy and the control of germination. New Phytologist 171(3):501-523. [ Links ]
Fouqué A. 1973. Espèces fruitières d'Amérique tropicale. Caricaceae. Fruits 28: 648-652. [ Links ]
Gashi B, Kasamedin AK, Mata V, Kongjika E. 2012. Effect of gibberellic acid and potassium nitrate on seed germination of the resurrection plants Ramonda serbica and Ramondana thaliae. African Journal of Biotechnology 11(20):4537-4542. [ Links ]
Grzybowski CR, de Castro O, Carvalho R, Panobianco M. 2012. Viability of barley seeds by the tetrazolio test. Revista Brasileira de Sementes 34(1):47-54. [ Links ]
International Seed Testing Association -ISTA-. 2007. International rules for seed testing. 46 p. [ Links ]
IUCN—The World Conservation Union. 2003. The IUCN Red List of Threatened Species. IUCN Species Survival Commission. IUCN Gland Switzerland and Cambridge UK. Available at website http://www.iucnredlist.org/. [ Links ]
Kyndt T, Romein-Peeters E, Van Droogenbroeck B, Romero-Motochi J, Gheyen G, Goetghebeur P. 2005. Species relationships in the genus Vasconcellea (Caricaceae) based on molecular and morphological evidence. American Journal of Botany 92:1033-1044. [ Links ]
Magdalita PM, Drew RA, Adkins SW, Godwin ID. 1997. Morphological molecular and cytological analysis of Carica papaya × C. cauliflora interspecific hybrids. Theoretical and Applied Genetics 95:224-229. [ Links ]
Manshardt RM, Wenslaff TF. 1989. Interspecific hybridization of papaya with other Carica species. Journal of the American Society for Horticultural Science 114(4):689-694. [ Links ]
Quintanilla YD. 1995. Caracterización de la papaya (Carica pubescens Lenné et Koch) de Cobquecura para aptitud industrial. Memoria título Ingeniero Agrónomo. Facultad de Agronomía Universidad de Concepción. Chillán, Chile. [ Links ]
Scheldeman X. 2002. Distribution and potential of cherimoya (Annona cherimola Mill.) and highland papayas (Vasconcellea spp) en Ecuador. Thesis submitted in fulfillment of the requirement for the degree of Doctor (Ph. D.) University of Gent Belgium. 165 p. [ Links ]
Tokuhisa D, Cunha D, Mantovani E, Hilst PC, Demuner AJ. 2007. Compostos fenólicos inibidores da germinação. Revista Brasileira de Sementes 29(3):180-188. [ Links ]
Tokuhisa D, Diaz DC, Alvarenga EM, Diaz LA, Marín SL. 2006. Tratamientos para superacao da dormencia em sementes de mamao. Revista Brasileira de Sementes 29:131-139. [ Links ]
Tunes LM, Bandinelli PG, Olivo F, Barros AC. 2009. Tratamento para superamento da dormencia em sementes de cevada. Scientia Agraria 10(1):15-21. [ Links ]
Van Droogenbroeck B, Kyndt J, Maertens I, Romein-Peeters E, Scheldeman X, Romero-Motochi JP, Van Damme P, Goethebeur P, Gheysen G. 2004. Phylogenetic analysis of the Highland papayas (Vasconcellea) and allied genera (Caricaceae) using PCRRFLP. Theoretical and Applied Genetics 108:1437-1486. [ Links ]
Varela M, Abreu A, Galon K, Brekemkamp DM, Pereira J, Queiroz M, Martins G, Coelho RI. 2011. Influencia de períodos de imersão em agua na emergência de plântulas de mamão ornamental. XV Encontro Latino Americano de Iniciação Científica XI Encontro Latino Americano de Pos Graduação. Universidade Federal de Espirito Santo, Guararena, Brasil. p. 1-4. [ Links ]
Vegas A, Trujillo G, Sandrea Y, Mata J. 2003. Obtención, regeneración y evaluación de híbridos intergenéricos entre Carica papaya y Vasconcellea cauliflora. Interciencia 28(12):710-714. [ Links ]
