










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
versão impressa ISSN 0122-8706
Corpoica cienc. tecnol. agropecu. vol.14 no.2 Mosquera jul./dez. 2013
FISIOLOGÍA VEGETAL
1 I.A. Professional researcher. Corpoica. C.I. La Selva. Rionegro, Antioquia, Colombia. gfranco@corpoica.org.co
2 PhD. Associate professor. Universidad Nacional de Colombia. Medellín, Colombia. jrcartag@unal.edu.co
3 D.Sc. Associate professor. Universidad Nacional de Colombia. Medellín, Colombia. gcorrea@unal.edu.co
Fecha de recepción: 12/06/2013. Fecha de aceptación: 19/07/2013
ABSTRACT
Gulupa (Passiflora edulis Sims) is currently receiving increasing acceptance in European markets. However, its short postharvest life constitutes its main marketing limitation, and makes it necessary to characterize its ripening characteristics in order to assure that fruits reaching the consumer are of the best possible quality. Predictive models were fitted to estimate flesh carotenoid content from fruit age and color change. For this purpose, a research was conducted on plant material from the Colombian National Germplasm Bank, administered by Corpoica. Fruit samples of known age were subjected to CIE L* a* and b* space-based colorimetric determination, which confirmed the existence of appreciable color changes since 84 days after flowering (DAF). Deep purple color was reached at around 98 DAF and very dark purple color at 105 DAF. In fruits on the vine, luminosity (L*) was observed to decline, while the value of coordinate a* increased and that of coordinate b* decreased. These changes feature the fruit's characteristic color. A similar trend was observed in the evolution of color in fruits off the vine (harvested at 91 DAF). In both fruits on and off the vine, carotenoids were found to increase along with fruit age. Two regression models were adjusted to estimate carotenoid content: a multiple model with color coordinates as predictors, and a polynomial model based on fruit age.
Key words: b-carotene, Cielab color, functional foods, tropical fruits.
RESUMEN
La gulupa (Passiflora edulis Sims) es un fruto aceptado en mercados europeos; pero, su corta vida en poscosecha limita la comercialización. Por lo tanto, es necesario, caracterizar algunos atributos de la maduración, para que el fruto mantenga las mejores condiciones de calidad. La investigación tuvo como objetivo estimar, a partir de los cambios de color del fruto y de su edad, la evolución de los carotenoides, mediante el uso de modelos que predijeran esta relación. Se utilizaron frutos con edad conocida para determinar el color con base en el espacio CIE L*, a* y b*. La colorimetría confirmó que hay evolución apreciable a partir de los 84 días después de floración (DDF), con color morado oscuro hacia los 98 DDF y púrpura muy oscuro a los 105 DDF. En frutos madurados en la planta, se apreció disminución en la luminosidad (L*), mientras que en las coordenadas a* el valor aumentó y el b* disminuyó, cambios que definen el color del fruto. Se observó una tendencia similar en frutos cosechados a partir de los 91 DDF. Tanto en los frutos madurados en la planta como en los cosechados, el contenido de carotenoides se incrementó a medida que la senescencia del fruto avanzaba. Modelos de regresión permitieron estimar el contenido de carotenoides: uno múltiple, con las coordenadas de color como predictoras y el otro polinomial, basado en la edad del fruto.
Palabras claves: b-caroteno, color Cielab, alimentos funcionales, frutos tropicales.
INTRODUCTION
The so-called "high-Andean fruits", like cape gooseberry, lulo, tree tomato and several species of Passiflora, among others, are considered to be enjoyable and highly demanded products on the part of consumers, being fresh marketing the most common presentation. These fruits have a short shelf life, during which they undergo a rapid deterioration of their physiological activity, which, in turn, affects their quality. Gulupa (Passiflora edulis Sims) stands out among the wide range of fruit species with potential for commercial development that have been recognized in the Andean region (Lobo, 2000). Most of the production of this species in Colombia is exported, amounting to 2,250 tons in 2012, with an FOB value of US $9,750,000 mainly exported to European markets (Agronet, 2012).
Some of food quality attributes are color, texture and flavor. The two former are crucial, since they can be used by the consumer to accept or reject a product even before trying it. Regarded as one of the main indicators of food quality, rind color is often used by the consumer to judge the degree of maturity of a product, and by the producer to determine harvest time (Pinzón, 2000; Artigas et al., 2002, Zapata et al., 2007; Vianna et al., 2008).
The dominant color tones in nature are red, green, orange and yellow. Throughout history, humanity has adapted itself to this food color range, to the point of having modified the color of some foods. In fact, this attribute is frequently used to take prompt decisions on whether a given food is edible or not (Artigas et al., 2002).
Food color is due to the presence of pigments, among which carotenoids (red and yellow compounds found in fruits, flowers, leaves, and animal fats) play an important role. Carotenoids come from carotenes in which hydroxyl, carbonyl, epoxy or carboxyl groups have substituted for atoms of hydrogen or for carbon oxygenated derivatives.
Carotenoids help to absorb radiant energy during photosynthesis (Primo, 1998), where they act as photoprotectors, inasmuch as they inhibit the spread of reactive oxygen species (ROS) and other free radicals (Mínguez et al., 2013), thus playing a protective role as antioxidants. However, the form of action, bioavailability and antioxidant properties of carotenes have not been thoroughly cleared yet (Piñeiro and Zudaire, 2009).
Certain authors have suggested that carotenes might be associated to a decreased risk of developing chronic degenerative diseases due to oxidative stress, while some others see them as nutritional complements (Burgos and Calderon, 2009; Mínguez et al., 2013).
The ontogeny of gulupa has not been thoroughly assessed, while that of yellow passion fruit has been found to undergo transformations that alter the physicochemical characteristics of the product (Vianna et al., 2008). These authors, as well as Vianna et al. (2005), found that yellow passion fruit's shell starts shifting from green to yellow around 64 days after flowering (DAF). They attributed this change to chlorophyll degradation, which is associated to the appearance of pigments, mostly carotenoids. Besides, they observed that this color change is not related to fruit juice color.
Several carotenoids such as b-cryptoxanthin, prolycopene, cis-z-carotene, z-carotene, b-carotene, a-carotene and 13 - cis-b-carotene have been found in yellow passion fruit juice (Holden et al., 1999; Silva and Mercadante, 2002). For their part, Rodríguez et al. (2008) only found b-carotene in this fruit's juice. These authors observed that carotenoid content depends on fruit origin and on the enzymatic reactions that produce these pigments. In addition, they found that fresh, commercial fruit and juice are mainly featured by z-carotene and b-carotene, the latter contributing to vitamin A content.
Along with the ripening process, gulupa shell color changes from green to very dark purple. Purple color acquisition starts at 80 DAF in the case of fruits maturing on the vine, and around 70 DAF for fruits maturing on the vine (Shiomi et al., 1996; Pinzón et al., 2007; Rodríguez and García, 2010). Ripe fruit color has the same intensity both for the former as for the later and it is achieved at around 90 DAF. Fruit flesh color also undergoes observable changes, which have been graded in four stages, from light cream-yellow to dark orange (Pinzón et al., 2007).
Quality assessment methods and their relation with horticultural products have been developed and applied for centuries, in so far as instrumental techniques having appeared some 80 years ago. Non-destructive, real time sensors have been recently implemented for quality measurement. Thus, electromagnetic, mechanical, electrochemical and statistical methods are currently available for such purposes (Abott, 1999). According to Slaughter (2009), non-destructive fruit quality assessment currently covers diverse attributes like aroma, color, composition, firmness, form, size and specific gravity.
In studying mango varieties "Manila" and "Ataulfo", Ornelas et al. (2008) found some significative correlations between flesh carotenoid content and the a* value of the CIE (Commission Internationale de l'Éclairage) as determined on fruit rind. Seeking to predict carotenoid behavior in tomato (Lycopersicum esculentum Mill), Zapata et al. (2007) fitted models relating lycopene and b-carotene with the a*/b* index. Contrastingly, Urfalino et al. (2011) found no relation between this index and lycopene content, whereas Fischer and Martínez (1999) reported a nonsignificant correlation between cape gooseberry fruit color and carotenoid content.
In this context, the current research sought to establish carotenoid content evolution as related to fruit age and color changes. This was attained by fitting some regression models, looking forward to future developments in commercialization parameter predictors.
MATERIALS AND METHODS
Location. An experimental crop was set in the department of Antioquia (Colombia), municipality of Rionegro, rural district of Llanogrande, at Centro de Investigación La Selva (La Selva Research Center), which belongs to Corporación Colombiana de Investigación Agropecuaria, Corpoica (Colombian Corporation of Agricultural Research). According to Holdridge Ecological Life Zones, the experimental site corresponds to a Lower Montane Rain Forest (LM-rf ). It is located at 6°7'49'' North, 75°24'49'' East, and 2090 masl. Average yearly temperature and precipitation are 17 °C and 1,917 mm, respectively. Average relative humidity (RH) is 78%; average sunshine, 1,726 hours/year; and average evapotranspiration, 1,202 mm.
Biological material. The experiment made use of 10 gulupa materials coming from the departments of Antioquia, Putumayo and Nariño (Colombia) and preserved at Banco de Germoplasma de la Nación Colombiana (The Colombian Nation's Germplasm Bank), which is administered by Corpoica. According to Ortiz (2010), when analyzed through AFLP and SSR molecular markers, these materials showed low genetic variability.
Fruits were obtained from flowers at the homogamous stage, with and without herkogamy, which were marked with color threads following protocol by Ángel et al. (2011). This stage was taken as day zero of fruit age. Sampling periods and age of fruits were then labeled in terms of DAF.
Experimental procedure. Color analyses were performed through destructive weekly samplings from 7 to 112 DAF. Postharvest analyses were carried out on fruits collected at 91 DAF, which were weekly monitored for 21 days; that is, at 98, 105 and 112 DAF. For its part, carotenoid content analysis started at 49 DAF. On all these dates, the sampling unit was made up of 10 fruits taken at random from the materials in question and making up balanced and independent samples with regard to fruit size. These samples were transported in styrofoam boxes containing dry ice at an approximate inner temperature of 4 °C. Carotenoid determinations were carried out at the Food Science Laboratory of Universidad Nacional de Colombia-Sede Medellín. Color assessment was done at the postharvest laboratory of Corpoica. The fruits off the vine were stored at room temperature (20 °C) and 70% relative humidity.
Juice extraction was carried out by cutting the fruits in halves and separating the shell from the flesh, which was then sieved through a tulle cloth to take apart the seeds while the liquid was collected in an ice refrigerated container. Carotenoid analysis was carried out on three samples from a mix containing the juice of 10 fruits.
Taking place at the moment of each sampling, fruit shell color determination made use of a Chroma Meter CR- 400 Konica Minolta® colorimeter with illuminant D65 in the CIE L*, a* and b* color space. The measurements were always taken at two opposite points on the equatorial zone of the fruits, thus obtaining parameters L* (luminosity), a* (green - red chromaticity), and b* (blue - yellow chromaticity) (Menéndez et al., 2006).
Carotenoid content determination followed the methodology proposed by Biswas et al. (2011). For such purpose, 1 g of sample was placed into a test tube, added with 5 mL of cold acetone and then left to stand in the refrigerator at 4°C for about 15 min, after which it was shaken in Vortex for 2 min. The mix was centrifuged in a medium capacity and high speed Hermle Labnet® Z366 centrifuge at 1,370 gravities for 10 min, the supernatant being collected in another test tube. The precipitate was extracted with 5 mL of cold acetone and then processed again through the same protocol. Ketone extracts were mixed and filtered through Whatmann paper No. 42. Absorbance was measured at 449 nm wave length in a Thermo Scientific® Multiskan Spectrum Spectrophotometer V1.2. Carotenoid concentration was determined through the corresponding calibration curve, using b-carotene as reference substance. Data were obtained in a SkanIt software 2.4.2 RE for Multiskan Spectrum®.
Two regression models were fitted to estimate carotenoid content: a multiple one based on color coordinates and a polynomial one based on fruit age. The predictive variables were the CIE color space coordinates L*, a* and b* for the former one, and DAF for the latter model.
RESULTS AND DISCUSSION
The ripening process was similar to those reported by Shiomi et al. (1996), Pinzón et al. (2007), and Rodríguez and García (2010); fruit shell changing from green to purple since 84 DAF, to reach dominant purple color at about 91 DAF. This same color stage was attained by the fruits off the vine at 98 DAF. Fruit color change is significant as it promptly provides useful information about the ripening stage.
The tri-stimulus colorimetric assessment confirmed the observations above, indicating considerable transformations since 84 DAF, age when the fruit starts changing from green to purple, to reach more intense tones at 91 DAF, deep purple at around 98 DAF, and very dark purple at 105 DAF. Under the conditions studied by Shiomi et al. (1996), these changes were observed since 70 DAF.
Luminosity (L*) was observed to decrease from 58.1, at 7 DAF, to 33.3, at 112 DAF, indicating that the fruit tends towards opaqueness; however, between 14 and 91 DAF, the value of L* remained constant. With regard to chromaticity, value a* changed from -12.9 to 6.5 during the same period time, while at end of the ripening process, coordinate b* shifted from 24.9 to 3.57, indicating proximity to grey tones, which, combined with the mentioned luminosity, feature this fruit's characteristic color. Both a* and b* were found to undergo remarkable changes since 84 DAF, whereas luminosity started shifting by 91 DAF, coinciding with the visual observations and with data reported by Flórez et al. (2012) for gulupa from Cundinamarca (Colombia).
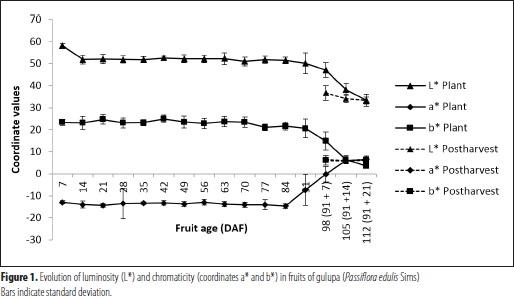
Coordinates L* and a* showed slower changes in the fruits on the vine than in those off the vine, in which both L* and b* were observed to decrease after harvest. This indicates that purple color also appears during postharvest, in an apparently faster process. In this respect, Shiomi et al. (1996) assert that postharvest development of purple color requires the fruit to be harvested between 80 and 90 DAF, as also observed in the current study. In related fruits like yellow passion fruit, shell color shift from green to yellow starts at 64 DAF on the lower part of the fruit and reaches 100% at 100 DAF (Vianna et al., 2008 and Menéndez et al., 2006). In these works, color change took place during the last third of the ripening process, as also reported by Shiomi et al. (1996) for gulupa, coinciding with the findings of the current research.
The a*/b* index followed the tendency of chromaticity a*, revealing changes since 84-91 DAF and varying between fruits on and off the vine. In the fruits on the vine, the a*/b* ratio underwent a remarkable increase since 84 DAF (Figure 2), coinciding with evident color changes from green to purple. For their part, the fruits off the vine showed a steeper slope in this ratio's increase, although they reached a lower final value. However, this difference was visually undetectable. Variation in this index is consistent with the evolution of each of the variables in the CIE color space.
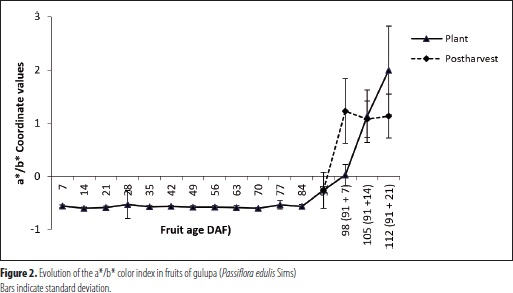
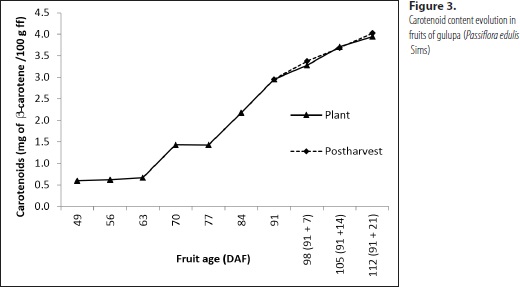
Figure 3 shows the evolution of carotenoid content in the analyzed gulupa juice samples. A steady increase can be observed to progress along with fruit age in both fruits on and off the vine, corresponding with previous reports on fruits that are also rich in this metabolite such as guava (Psidium guajava L.) (González, 2010) and cape gooseberry (Physalis peruviana L.) (Fischer and Martínez, 1999). In yellow passion fruit, Holden et al. (1999) determined that the content of b-carotene was 0.525 mg per 100 g of fresh fruit (f f ), whereas Burgos and Calderón (2009) found a total carotenoid content of 0.743 mg per 100 g f f. These values are lower than those found in the current research, which are, in turn, higher than those reported by Holden et al. (1999) for mango (Mangifera indica L.), orange (Citrus sinensis (L.) Osbeck), papaya (Carica papaya L.) and avocado (Persea americana Mill); by Montefiori et al. (2009) for kiwi (Actinidia sp.) and by Fischer and Martínez (1999) for cape gooseberry. Yet, our results regarding this parameter are lower than those reported for carrot (Daucus carota L.) (Holden et al., 1999), and some varieties of guava (González, 2010).
Carotenoid content was found to be significatively related to color coordinates and fruit age (P ≤ 0.05). In effect, by means of two regression models, carotenoid content was estimated as a function of: (i) color coordinates (a*, b* and a*/b*), in a multiple model; and (ii) DAF, in a polynomial model (Table 1 and Figure 4).

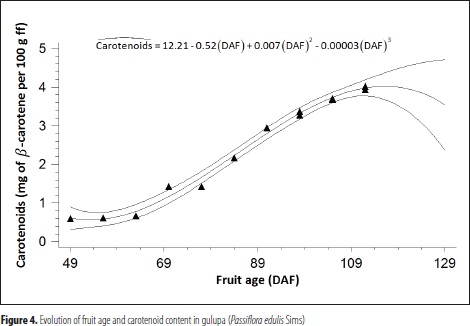
The central line represents the estimated function of the third order polynomial model. Upper and lower lines correspond to the 95% confidence limits for the expected values; rectangles indicate observed data.
In addition to previous reports by Zapata et al. (2007), who were able to estimate carotenoid content in tomato through its a*/b* index; and Ornelas et al. (2008), who associated value a* of the CIE in mango varieties "Manila" and "Ataulfo" with carotenoid content; the current research allowed correlating color coordinates a*, b*, as well as the a*/b* index, with carotene concentration in gulupa by means of a linear model (R2 = 0.89). These results differ from those obtained by Urfalino et al. (2011), who failed to find any association between lycopene and a*/b* color index in tomato. In the current research, carotenoid content correlated with fruit age (DAF) through a polynomial model at highly predictive levels (R2 = 0.98).
CONCLUSIONS
Two regression models are contributed to the estimation of carotenoid content in gulupa flesh. The multiple model is appropriated for predicting carotenoids content when using coordinates L*, a* and b* of the CIE color space; while the polynomial model is adequate when using DAF as a predictor variable.
Color variation was found to be associated to carotene accumulation and to the CIE L*, a* and b* color space. This allows estimating carotenoid content through these color parameters, including the a*/b* index.
Across the studied fruit development periods, the CIE L*, a* and b* color space showed differences that explained the visual shift from green to purple fruit color. Thus, this is a suitable color index for this fruit.
Fruit age (DAF) correlated with carotene accumulation in the flesh. Hence, it can be said that the riper the product, the more of this metabolite it contains.
ACKNOWLEDGEMENTS
The authors want to thank Dr. Benjamín Rojano and Chemical Engineer Ana María Piedrahíta C., Head and Young Researcher, respectively, of the Food Science Laboratory (Laboratorio de Ciencia de los Alimentos) of Universidad Nacional de Colombia, sede Medellín, for their collaboration in fruit total carotenoid content determination.
REFERENCES
Abbott JA. 1999. Quality measurement of fruits and vegetables. Postharvest Biology and Technology 15(3):207-225. [ Links ]
Agronet. 2013. Comercio exterior de Colombia. Exportaciones del sector agropecuario. http://www.agronet.gov.co; consulta: abril, 2013. [ Links ]
Ángel C, Nates G, Ospina R, Melo C, Amaya M. 2011. Biología floral y reproductiva de la gulupa Passiflora edulis Sims f. Edulis. Caldasia 33(2):433-451. [ Links ]
Artigas JM, Capilla P, Pujol J. 2002. Tecnología del color. España. Universidad de Valencia, 428 p. [ Links ]
Biswas AK, Sahoo J, Chatli MK. 2011. A simple UV-Vis spectrophotometric method for determination of b-carotene content in raw carrot, sweet potato and supplemented chicken meat nuggets. Food Science and Technology 44(8):1809-1813. [ Links ]
Burgos JT, Calderón FR. 2009. Determinación del contenido de carotenoides totales en ocho especies de frutas y verduras comercializadas en la zona metropolitana de San Salvador. Bachelor's thesis in Chemistry and Pharmacy. Universidad de El Salvador. Escuela de Química y Farmacia. San Salvador, El Salvador, Centro América. 143 p. [ Links ]
Fischer G, Martínez O. 1999. Calidad y madurez de la uchuva (Physalis peruviana L.) en relación con la coloración del fruto. Agronomía Colombiana 16(1-3):35-39. [ Links ]
Flórez LM, Pérez LV, Melgarejo LM, Hernández S. 2012. Caracterización fisicoquímica, fisiológica y bioquímica del fruto de gulupa (Passiflora edulis Sims) como indicadores para el punto óptimo de cosecha. In: Melgarejo LM. Ed. Ecofisiología del cultivo de la gulupa (Passiflora edulis Sims). Universidad Nacional de Colombia, Sede Bogotá, Ministerio de Agricultura y Desarrollo Rural, Ocati. Bogotá. p. 53-79. [ Links ]
González IA. 2010. Caracterización química del color de diferentes variedades de guayaba (Psidium guajava L.) colombiana (tesis Magister en Ciencias - Química). Universidad Nacional de Colombia. Escuela de Ciencias. Departamento de Química. Bogotá. 84 p. [ Links ]
Holden JM, Eldridge AL, Beecher GR, Buzzard IM, Bhagwat AS, Davis CS, Douglass LW, Gebhardt ES, Haytowitz D, Schakel S. 1999. Carotenoid content of U.S. Foods: An Update of the Data base. Journal of Food Composition and Analysis 12:169- 196 [ Links ]
Lobo M. 2000. Papel de la variabilidad genética en el desarrollo de los frutales andinos como alternativa productiva. In: 3er Seminario de Frutales de Clima Frío Moderado. Memorias. Centro de Desarrollo Tecnológico de Frutales. Manizales. p. 27-36. [ Links ]
Menéndez OM, Evangelista S, Arenas M, Bermúdez K, Martínez A, Jiménez A. 2006. Cambios en la actividad de a-amilasa, pectinmetilesterasa y poligalacturonasa durante la maduración del maracuyá amarillo (Passiflora edulis var. Flavicarpa Degener). Interciencia 31(10):728-733. [ Links ]
Mínguez MI, Pérez A, Hornero D. 2005 Pigmentos carotenoides en frutas y vegetales; mucho más que simples colorantes naturales. Revista CTC Alimentación 26:108-113. [ Links ]
Montefiori M, Mcghie TK, Hallett IC, Costa G. 2009. Changes in pigments and plastid ultrastructure during ripening of greenfleshed and yellow-fleshed kiwifruit. Scientia Horticulturae 119(4):377-387. [ Links ]
Ornelas JJ, Yahia EM, Gardea AA. 2008. Changes in external and internal color during postharvest ripening of 'Manila' and 'Ataulfo' mango fruit and relationship with carotenoid content determined by liquid chromatography-APcI+-time-of-flight mass spectrometry. Postharvest Biology and Technology 50(2- 3):145-152. [ Links ]
Ortiz DC. 2010. Estudio de la variabilidad genética en materiales comerciales de gulupa (Passiflora edulis f. var. edulis Sims) en Colombia. Tesis Magister en Ciencias Agrarias - Genética y Mejoramiento. Universidad Nacional de Colombia, Sede Bogotá. Facultad de Agronomía. 127 p. [ Links ]
Piñeiro E, Zudaire M. 2009. Los carotenoides de los alimentos. http://consumer.es; consulta: enero 2013. [ Links ]
Pinzón IM, Fischer G, Corredor G. 2007. Determinación de los estados de madurez del fruto de la gulupa (Passiflora edulis Sims). Agronomía Colombiana 25(1):83-95. [ Links ]
Pinzón MI. 2000. Propiedades físicas de cosecha y poscosecha de frutos de lulo "La Selva". In: 3er Seminario de Frutales de Clima Frío Moderado. Memorias, Centro de Desarrollo Tecnológico de Frutales. Manizales. p. 386-397. [ Links ]
Primo YE. 1998. Química de los alimentos. España. Editorial Síntesis. 461 p. [ Links ]
Rodríguez DB, Kimura M, Godoy HT, Amaya FJ. 2008. Updated brazilian database on food carotenoids: Factors affecting carotenoid composition. Journal of Food Composition and Analysis 21(6):445-463. [ Links ]
Rodríguez M, García C. 2010. Poscosecha, procesamiento y análisis nutracéutico de gulupa (Passiflora edulis Sims.) y curuba (Passiflora tripartita var. mollisima). Primer Congreso Latinoamericano de Passiflora. Resumen. Memorias. CEPASS, Huila. Neiva, Huila, Colombia. p 107. [ Links ]
Shiomi S, Wamocho LS, Agong SG. 1996. Ripening characteristics of gulupa on and off the vine. Postharvest Biology and Technology 7(1-2):161-170. [ Links ]
Silva SR, Mercadante AZ. 2002. Composição de carotenóides de maracujá-amarelo (Passiflora edulis flavicarpa) in natura. Cienc. Tecnol. Aliment. 22(3):254-258. [ Links ]
Slaughter DC. 2009. Evaluación de métodos no destructivos para la detección de la madurez en mangos: Revisión bibliográfica e identificación de las necesidades de investigación a futuro. Biological and Agricultural Engineering. University of California, Davis. http://mango.org; consulta: enero 2013. [ Links ]
Urfalino DP, Quiroga A, Worlock J. 2011. Efecto del deshidratado en el contenido de licopeno en distintos cultivares de tomate. Instituto Nacional de Tecnología Agropecuaria INTA. http://inta.gob.ar; consulta: enero 2013. [ Links ]
Vianna T, Resende ED, Pereira SMF, Viana AP, Rosa PC, Carlos LA, Vitorazi L. 2008. Influência dos estádios de maturação sobre as características dos frutos de maracujá - amarelo. Bragantia 67(2):521-525. [ Links ]
Vianna T, Resende ED, Viana AP, Carrielo RC. Pereira SMF, Carlos LA, Vitorazi L. 2005. Influência dos estádios de maturação na qualidade do suco do maracujá-amarelo. Rev. Bras. Frutic. Jaboticabal 27(3):472-475. [ Links ]
Zapata L, Gerard L, Davies C, Oliva L, Schvab M. 2007. Correlación matemática de índices de color del tomate con parámetros texturales y concentración de carotenoides. Ciencia, Docencia y Tecnología 34(18):207-226. [ Links ]
