










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
versão impressa ISSN 0122-8706
Corpoica cienc. tecnol. agropecu. vol.16 no.1 Mosquera jan./jun. 2015
1 Licenciada en Educación Agropecuaria, PhD. Investigadora Corpoica-CI La Selva. cmedina@corpoica.org.co
2 Agrónomo, PhD. Investigador Corpoica-CI La Selva. mlobo@corpoica.org.co
3 Ingeniero Forestal, Universidad Nacional de Colombia sede Medellín. aacastan@gmail.com
4 Ingeniero Industrial, MSc. Minería MTC. ecardon82@gmail.com
Fecha de recepción: 22/10/2014. Fecha de aceptación: 16/03/2015
Resumen
La investigación realizada en el CI La Selva de Corpoica, Rionegro-Antioquia, a 2.120 msnm, formación ecológica bosque húmedo montano bajo, buscó caracterizar el crecimiento y desarrollo de plantas de mortiño, Vaccinium meridionale Sw., provenientes de poblaciones espontáneas, obtenidas a partir de semilla y multiplicación vegetativa. Estas se trasplantaron a rizotrones, con vidrios laterales en dos de sus costados, para el registro del crecimiento de las raíces. Las plantas tomaron 1.663 y 1.367 días, desde la siembra de la semilla y el enraizamiento de estacas a floración, respectivamente. Las raíces de ambos tipos de material no presentaron pelos radicales. En las dos clases de plantas no se apreciaron períodos de alternancia en el crecimiento de las órganos aéreos y los subterráneos, con relaciones alométricas entre la parte aérea y las raíces del orden de 6,3 para los individuos obtenidos por semilla y 7,0 para los de multiplicación asexual. A pesar de que se detectaron flujos de floración, se apreció formación continua de flores en ambos materiales. Los registros de desarrollo, en las plantas propagadas sexualmente, exhibieron una mayor dispersión que aquellos obtenidos con las multiplicadas vegetativamente.
Palabras claves: Alometría, crecimiento y desarrollo, fenología, acumulación materia seca
Abstract
A research was carried out at Corpoica's La Selva, Experiment Station located at Rionegro, Antioquia, Colombia, at 2.120 masl in the ecological zone Low Mountain Rain Forest. The objective was to characterize growth and development of andean blueberry Vaccinium meridionale Sw., plants, obtained by seed and clonal propagation, from a wild population of the species. Four plants of each type were transplanted to rizhotrons, with glasses on two sides, which allowed the root growth recording. The plants coming from seed spent 1.663 days from sowing to flowering, and those derived from rooted cuttings 1.367 days. In both types, the roots lack root hairs. Also, in both kinds of materials, there was evidence for non alternating periods of root and aerial organs growth, with 6.28 and 7.00 as allometric indexes between the aerial part and the roots for the individuals obtained by sexual and asexual propagation. In spite of the occurrence of periods of increased flower production, after the beginning of the flowering to the end of the research, these organs were detected in both types of materials. There was a greater interplant variability in the growth values obtained in the sexual plants than those of the clonal ones.
Key words: Dry matter accumulation, growth, phenology, architectural traits
Introducción
El género Vaccinium, nombre asignado por Linneo en 1737 (Vander 2004), comprende cultivos importantes de la zona templada como el arándano, cuya base genética está conformada por V. corymbosum, V. ashei y V. angustifolium (Galleta y Ballington 1996). En la zona altoandina colombiana, crecen en forma espontánea, especies del clado anterior; una de estas es V. meridionale Sw., conocida en Colombia como mortiño, agraz, vichachá o camueza. Este es un frutal promisorio espontáneo (Corantioquia 2003), cultivado incipientemente y cuyo fruto tiene demanda creciente a nivel nacional e internacional por su valor antioxidante (Gaviria et al. 2009; Gaviria et al. 2012), con potencial dentro del marco del Tratado de Libre Comercio (TLC) con Estados Unidos de América. Las bayas se obtienen principalmente a partir de procesos extractivistas de los bosques que generan destrucción de algunas poblaciones naturales y erosión genética y de algunas parcelas pequeñas sembradas sin recomendaciones técnicas (Torres et al. 2009). El taxón exhibe un fruto dulce, comestible, con sabor agradable, el cual se emplea en la elaboración de mermeladas, pasteles y helados (Romero 1991).
Las especies de Vaccinium contribuyen a la diversidad biológica, por estar correlacionadas con la abundancia de mamíferos y aves presentes en los hábitats de su ocurrencia (Alaback y Herman 1988; Halpern y Spies 1995; Carey 1995; Carey y Johnson 1995). En las regiones de Colombia donde crece el mortiño, es una fuente importante de alimento para la fauna silvestre, la cual, a su vez, actúa como dispersora natural de la especie ya que transportan su semilla a otros lugares después de consumir los frutos (Corantioquia 2003).
En Colombia, el mortiño, Vaccinium meridionale, representa una alternativa potencial importante, ya que las poblaciones naturales están ubicadas en el piso montano (2.200-3.200 msnm), correspondiente al límite superior de la zona agroecológica de cultivos ilícitos como la coca (Erythroxylum coca Lam.) y la amapola (Papaver somniferum L.) (Matteucci y Morello 2001); lo anterior indica que podría servir como opción para la reconversión de estos cultivos.
El mortiño se puede propagar vegetativa o sexualmente (Torres et al. 2009). Esto puede causar diferencias en el desarrollo del tallo y las raíces, aspecto reportado en diversas especies arbóreas, como cacao, Theobroma cacao L. (León 2000), aguacate, Persea americana Mill. (Bernal y Díaz 2005) y matarratón Gliricidia sepium ( Jacq.) Kunth ex Walp (Torrico y Cardona s.f.). Al respecto, se ha señalado que la arquitectura del dosel juega un papel esencial en la captura de la luz por su efecto en el traslape de las hojas y el autosombrío. Esto determina la colocación de biomasa en los órganos productivos y la tasa de crecimiento (Kawamura y Takeda 2004); con indicación de que hay diferencias marcadas en la arquitectura de las copas a nivel intra e interespecífico (Givnish 1988; Valladares et al. 2000). Es necesario conocer detalladamente el crecimiento de la copa de los árboles y los tipos de ramas que conforman su estructura para establecer prácticas culturales relacionadas con el manejo tecnificado de la plantación (Avilán et al. 1995). Hallé et al. (1978) y Hallé y Orcutt (1978) consideran las ramas como las unidades básicas de la arquitectura de los árboles.
Con base en lo anterior, se realizó la presente investigación, la cual buscó conocer el desarrollo fenológico y los patrones de crecimiento en plantas propagadas sexual y asexualmente y se planteó como hipótesis de trabajo, que las relaciones alométricas entre la raíz y la parte aérea pueden ser un indicativo de los flujos de crecimiento de la planta. De comprobarse lo anterior, se dispondría de una herramienta para determinar prácticas de manejo en los cultivos, al igual que el comportamiento productivo asociado con el método de multiplicación.
Materiales y métodos
Localización. El trabajo se desarrolló en el Centro de Investigación La Selva de la Corporación Colombiana de Investigación Agropecuaria (Corpoica), en Rionegro, Antioquia, a 2.120 msnm, con temperatura promedio de 17 °C y humedad relativa media del 78 %, localizado en la zona de vida bosque húmedo montano bajo (bh-MB) a 06°08'06'' de latitud norte y 75°25'03'' de longitud oeste.
Material vegetal. El estudio se realizó con semillas y estacas enraizadas de la accesión ILS 4368, del Sistema de Bancos de la Nación Colombiana para la Alimentación y la Agricultura, que provienen de una población espontánea del corregimiento de Santa Elena, vereda Chorro Clarín, municipio de Medellín, con coordenadas N 06°15'45" W 75°29'41", ubicada a 2.500 msnm, en el sotobosque de árboles de pino pátula (Pinus patula Schelet y Cham) y ciprés (Cupressus pygmaea Sarg).
Siembra plantas. 8 rizotrones de 40 cm de largo por 40 cm de ancho y 50 cm de profundidad, con vidrios transparentes en 2 de los lados opuestos fueron empleados para la siembra del material vegetal. Para las lecturas se dibujaron en los vidrios cuadrículas de 2 x 2 cm2 las cuales se cubrieron con plástico negro, removible durante las lecturas, para la medición del crecimiento de las raíces. Como sustrato de siembra, por rizotrón, se utilizó una mezcla de aserrín, arena, turba y gallinaza en relación de 4:4:3:1 y, adicionalmente, fueron incorporados a la mezcla 100 gramos de micorriza. El estudio incluyó 4 plantas provenientes de semilla y 4 de propagación vegetativa, las cuales tenían al inicio de la investigación 1.073 y 534 días de sembradas, respectivamente. Por su parte, las plantas derivadas de multiplicación asexual se obtuvieron a partir de estacas enraizadas previamente, con aplicación de ácido indol-acético.
Diseño experimental. Los estudios se llevaron a cabo con un diseño completamente al azar con dos tratamientos (multiplicación sexual y vegetativa) y cuatro repeticiones. Las unidades experimentales correspondieron a una planta por rizotrón.
Registro de variables. Cada rama principal se enumeró y se marcaron las ramificaciones existentes y las emisiones desarrolladas a través del tiempo. Se realizaron observaciones mensuales en las cuales se registró la longitud y el diámetro basal, así como también el estado fenológico, de acuerdo a la metodología utilizada por Michigan State University (2003), con modificaciones para Vaccinium meridionale. Para evaluar el desarrollo de las raíces, se contó periódicamente el número de cuadrículas que estas cortaban en las dos caras del rizotrón.
Materia seca (MS). La acumulación de MS se midió en los órganos presentes en la planta luego de la finalización del registro de información en dos individuos de cada sistema de propagación. Después de extraídas las plantas, se tomó el peso húmedo de todos los órganos y, posteriormente, se secaron en una estufa Memmert tipo UL 80, a 70 °C hasta alcanzar peso constante por órgano.
Arquitectura de la planta. Con los cuatro individuos por tratamiento, se calcularon los promedios e intervalos y se aplicó la prueba de t para comparar, en cada uno de los rasgos arquitecturales, las medias entre las plantas propagadas sexual y asexualmente conforme a los procedimientos aplicados por Kawamura y Takeda (2002), correspondientes a las variables: altura de planta, crecimiento de los brotes, longitud media de estos, número de brotes vegetativos, crecimiento (altura cm/día), diámetro basal del tallo (mm) y área de la copa (m2).
Fenología. Con el número de estados fenológicos presentes en cada uno de los brotes marcados a través del tiempo, se obtuvo un promedio por fecha de cada estado fenológico presente y se graficaron estos valores para reconocer el comportamiento de cada fase a través del período de estudio.
Modelo de crecimiento del vegetal. Para modelar el crecimiento, con respecto al período de desarrollo de las plantas provenientes de semilla y estacas enraizadas y de acuerdo a las variables de respuesta registradas, se empleó el modelo Arima Estacional (Box et al. 1994), para la estimación de series temporales en las cuales el tiempo juega un papel fundamental. La metodología libera al investigador de seleccionar modelos, lo que permite que los propios datos temporales de la variable indiquen la característica de la estructura probabilística subyacente.
Resultados y discusión
Los estudios sobre el grupo de los Vaccinium en Colombia son relativamente escasos, a pesar de ser una planta promisoria, original de la zona altoandina, con un alto potencial de mercado, dado su capacidad antioxidante. Por ello, como un apoyo preliminar de la especie, se presentan los siguientes resultados relacionados con el crecimiento y el desarrollo de plantas derivadas de dos tipos de multiplicación.
Modelos del crecimiento de la parte aérea
Para explicar el crecimiento de los órganos aéreos se graficó la tendencia de la longitud de los brotes formados, en las cuatro plantas provenientes de semilla y las cuatro derivadas de multiplicación vegetativa. Esto permitió colegir un comportamiento similar de aumento a lo largo del período del registro de la información de los individuos por tipo de planta.
En los individuos provenientes de multiplicación sexual y vegetativa se pudo observar que el proceso de crecimiento promedio no es estacionario en cuanto a la media y la varianza, tal como se observa en las figuras 1 y 3 correspondientes a las dos categorías de plantas, las cuales fueron funciones del tiempo del proceso. Por ello, para estabilizar la varianza se transformaron los datos, en ambos casos, a través de la ecuación Yt=log (Zt), en la cual Yt corresponde a la lectura transformada de la lectura t y Zt al logaritmo natural del valor de crecimiento obtenido en dicha lectura, lo cual redujo la variabilidad del factor estudiado y produjo el efecto esperado en la varianza, como se aprecia al comparar las figuras 1 y 2, al igual que la 3 con la 4. Esto permitió obtener un modelo altamente significativo en ambos casos, que explica el crecimiento de las plantas derivadas de semilla y clonación, el cual corresponde a la ecuación:
Yt =Yt-1+at, donde Yt es la lectura a calcular, Yt-1 es la lectura anterior y at= error en el tiempo t.
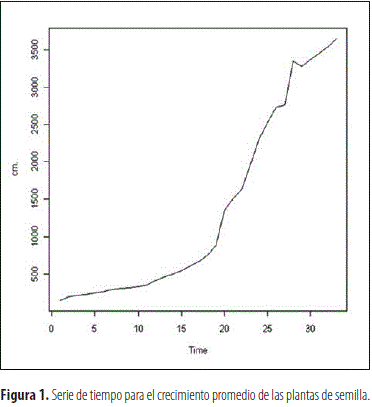
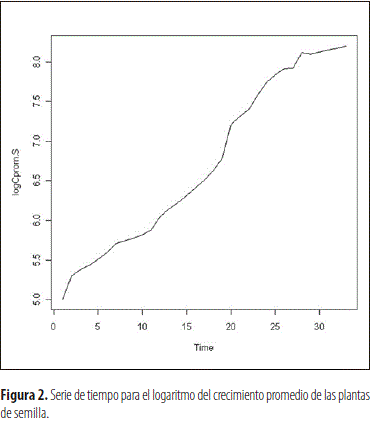
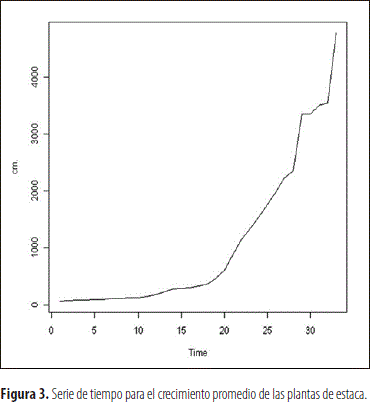
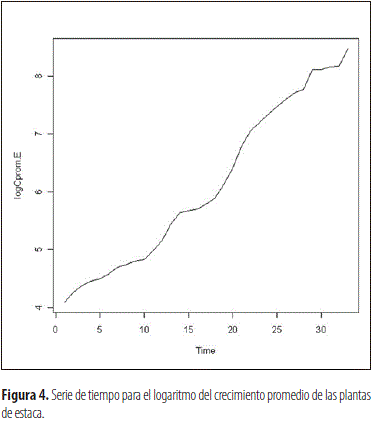
Evaluación de la parte radical de la planta
De acuerdo con observaciones realizadas en campo, bajo estereoscopio y microscopios, el sistema radical del mortiño, evaluado en este trabajo, está formado por finas raicillas, que presentan un desarrollo superficial de poca extensión que es incluido en las figuras 5A y B, caracterizado por la ausencia de pelos absorbentes, tanto en las plantas derivadas de semilla como de estacas (figuras 6A y B), resultado similar al reportado para los arándano (V. corymbosum), especie del género Vaccinium (Contreras 2010, Krewer et al. 1997). Esto en los arándanos conduce a una restricción en la absorción de agua, que causa un crecimiento más lento de las raicillas que el de otras plantas y las torna más sensibles a daños por déficit hídrico, aspectos que indican la necesidad del asocio con micorrizas ericoidales (Garret 1988).

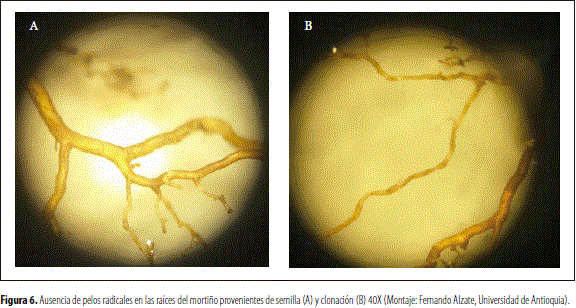
En las gráficas de crecimiento de la raíz, con las plantas derivadas de propagación sexual y asexual, se apreció una fase logarítmica caracterizada por un crecimiento lento pero continuo durante los 789 días después del trasplante, sin que se apreciaran períodos de reposo tal como se observa en las figuras 7A y B. Estas exhibieron dispersión de los datos, debido a que el desarrollo no se inició simultáneamente en todas las plantas.
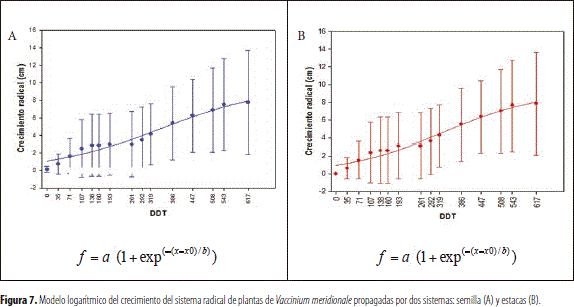
En el desarrollo de la parte aérea y la raíz, no fue evidente, en los períodos de toma de la información, la presencia de antagonismo entre el crecimiento de ambas estructuras (figura 7), con un patrón de desarrollo continuo. En el contexto anterior, se ha encontrado en el arándano (V. corymbosum) una reducción del crecimiento de las raíces nuevas y brotes durante el verano, cuando se termina la formación y el desarrollo de los frutos (Retamales y Lobos 2008); lo anterior es una consecuencia de la competencia con los frutos por carbohidratos y minerales durante el desarrollo; con indicación, por parte de los investigadores citados, de que la cosecha de las bayas permite un mayor crecimiento de brotes y raíces nuevas.
Acumulación de materia seca y peso fresco
En este primer análisis exploratorio, al tomar en consideración la definición de asignación de biomasa propuesta por Poorter y Nagel (2000), la cual señala que esta se refiere a la cantidad de materia seca presente en los diferentes órganos de la planta con respecto a la cantidad total, es clara una asignación diferencial de fotoasimilados entre los órganos evaluados en los dos tipos de materiales y una acumulación mayor de la materia seca en los tallos, seguida de las hojas y posteriormente en la raíz (tabla 1).
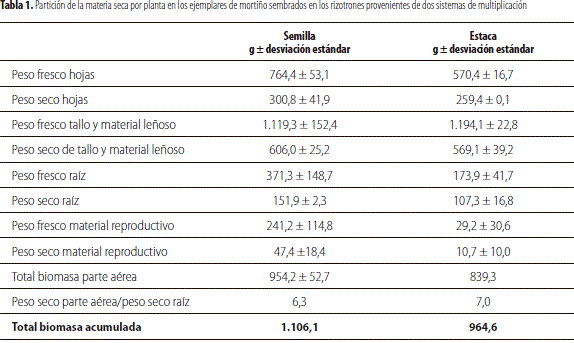
Como se puede apreciar en la tabla 1, la partición de materia seca, entre la parte aérea y la raíz en los individuos derivados de semilla y propagación vegetativa, señaló un índice alométrico con valores de 6,28 y 7,00, para ambos tipos, sin diferencia significativa entre estos, al considerar el bajo número de unidades muestrales por categoría, 2 plantas en cada caso; adicionalmente, se pudo observar que los materiales propagados asexualmente muestran una menor dispersión en los datos registrados en el peso fresco que en el seco, con una mayor dispersión en aquellas provenientes de estaca, en los pesos secos de tallo más material leñoso y de la raíz.
El peso seco es el criterio más apropiado para medir el crecimiento y la magnitud del sistema de asimilación de la planta, referido frecuentemente al área foliar (Taiz y Zeiger 1991). La distribución de la materia seca en las plantas está determinada por la ontogenia del cultivo, el genotipo, la presencia y la calidad de sumideros, y la distancia entre el sumidero y la fuente (Shibles 1987).
Como se puede apreciar en la tabla 1, la mayor acumulación de MS se obtuvo en los tallos de ambos tipos de materiales 606,0 g en los provenientes de semilla y 569 g en los de estaca, seguidos por las hojas 300,8 g y 259,4 g, respectivamente. Con una menor acumulación en la parte aérea de 151 g y 107 g en la raíz, con un total de MS total del orden de 1.106 g y 946 g en los individuos derivados de semilla y 1.106 de aquellos obtenidos por clonación.
La partición de la biomasa entre los tres órganos vegetativos de las plantas vasculares: hojas, tallos y raíces, y la localización de fotoasimilados para la elaboración de nuevos órganos, son componentes de la historia del vegetal y la dinámica poblacional para la modelación del cambio climático y las tendencias evolutivas (Niklas y Enquist 2002); sobre esto se ha indicado que la distribución de asimilados dentro de la planta está principalmente regulada por los órganos de demanda: tallos, raíces y frutos (Tekalign y Hammes 2005).
La mayor acumulación de materia seca en el tallo concuerda con la afirmación de que todas las funciones del órgano se deben integrar para el soporte de la planta y que la mayoría de estas estructuras, en el vegetal, no se pueden perder, es decir, que estas no se desprenden de la planta por efecto de la senescencia, con afirmación de que las hojas son el segundo órgano en importancia de acumulación de MS (Lewis 1997). Esto indica que el organismo está desarrollando, en esta primera etapa, toda la superficie de captación de energía y de producción de fotoasimilados para el sostenimiento de las partes formadas y para suplir las demandas de los nuevos vertederos en formación, al igual que para el reemplazo de follaje que entra en etapa de senescencia, por la falta de capacidad para cumplir sus funciones fisiológicas de biosíntesis (Lewis 1997). Sobre lo anterior, se ha afirmado que la distribución de la MS entre diversas partes de la planta corresponde a un equilibrio funcional que depende de la época y el estado de desarrollo del individuo (Bugarínn-Montoya et al. 2002).
En las plantas clonales se reconocen dos tipos de organización: genets y ramets (Albert et al. 2003). Los primeros están compuestos por todos los tejidos que se originan de un zigoto, mientras los segundos son la parte potencialmente independiente de un genet (Eriksson 1993). Se ha asumido que la diversidad genética es más baja en las plantas clonales (Harper 1981); sin embargo, estudios diversos han señalado que las poblaciones de estas se caracterizan por polimorfismo genético elevado (Hamrick y Godt 1989; Widén et al. 1994).
La dispersión menor de los datos en los materiales propagados por estaca se puede atribuir a que estos pertenecen al mismo o pocos genets, en tanto que las de semilla provienen de la F1 de polinizaciones entre estos, lo que produce recombinación de genets y segregación por esta vía. Al respecto, se encontró amplio polimorfismo en las plantas de la población de Santa Elena (Medina 2010), de la cual proviene el material sexual incluido en el estudio, con indicación de que la diferenciación genotípica entre las plantas de las poblaciones es común y que esta se puede dar en escalas espaciales tan pequeñas como a un par de metros e incluso en centímetros (Stenström et al. 2002).
La proporción de acumulación de materia seca entre las raíces y los tallos puede ser modificada por factores exógenos. En el contexto precedente se afirma que la fertilidad del suelo puede alterar la relación de biomasa entre las raíces y los tallos (Vanninen y Mäkelä 1999). Adicionalmente, se ha señalado que, durante la vida de los vegetales, la proporción de fitomasa entre las raíces y los tallos sufre cambios (Reed et al. 1995 y Vanninnen los tallos sufre cambios (Reed et al. 1995 y Vanni et al. 1996, citados por Vanninen y Mäkelä 1999).
Morfología y morfometría de las plantas de mortiño propagadas sexual y asexualmente
Mediante la descripción morfométrica de las plantas de mortiño o agraz se buscó conocer los patrones de desarrollo de los individuos multiplicados sexual y asexualmente.
Parte aérea
En la tabla 2 se incluyen los promedios e intervalos de variables relacionadas con la arquitectura de las plantas provenientes de semilla sexual y estacas; el diámetro basal del tallo exhibió, bajo el conjunto de condiciones del ensayo, un valor superior en los individuos provenientes de semilla al de las plantas clonales, con un área de copa mayor en estas últimas en comparación con las derivadas de simientes. En los atributos restantes, altura de planta, crecimiento, longitud media y número de brotes, al igual que en el incremento en altura, no fue evidente una diferencia con confiabilidad estadística entre los individuos de ambas clases.
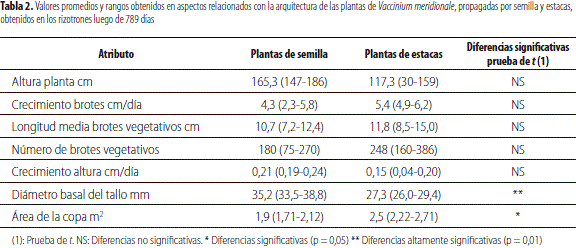
Con relación al área de la copa, se apreció cualitativamente un crecimiento más erecto (ortotrópico) en las plantas obtenidas por reproducción sexual y más postrado (plaugiotrópico) en aquellas derivadas de multiplicación vegetativa lo cual produce copas más extendidas. Esto podría indicar, en principio, que la siembra de estas últimas requeriría de un mayor espaciamiento entre los individuos o el desarrollo de sistemas de poda diferentes a aquellos a realizar en plantas logradas por semillas. Al respecto, se ha indicado que la arquitectura de la copa juega un papel importante en la captura de la luz por su efecto en el traslape de las hojas y el autosombrío de esta, en la partición en tejidos productivos y no productivos y en la tasa de crecimiento (diversos autores, citados por Kawamura y Takeda 2002), con afirmación de que las diferencias en características tanto entre especies como dentro de estas, se han interpretado en términos del potencial adaptativo a diferentes ambientes lumínicos (Givnish 1988; Valladares et al. 2000).
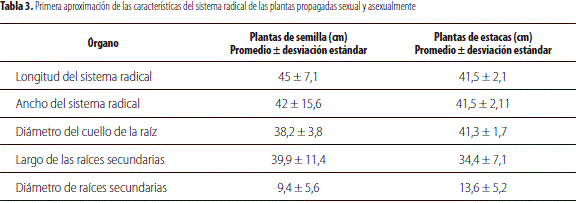
Sistema radical
En la tabla 3 se puede apreciar que el largo y el ancho de las raíces del mortiño, V. meridionale, es superficial, sin superar el diámetro de las copas. Al respecto, Retamales y Lobos (2008) indicaron que el arándano, V. corymbosum, exhibe una distribución superficial y que sus raíces están limitadas al área demarcada por la línea de goteo de la planta.
Fenología de las plantas
En la tabla 4 se presenta la duración en días de las diversas etapas ontogénicas para las plantas bajo estudio, obtenidas mediante propagación sexual y asexual sembradas en los rizotrones. En esta se puede apreciar que los tiempos acumulados fueron significativamente superiores en las plantas obtenidas por semilla, a partir de la tercera etapa: trasplante a rizotrones, hasta la sexta: floración a fruto maduro con un total de 1.663 días en las primeras y 1.377 en aquellas clonadas.
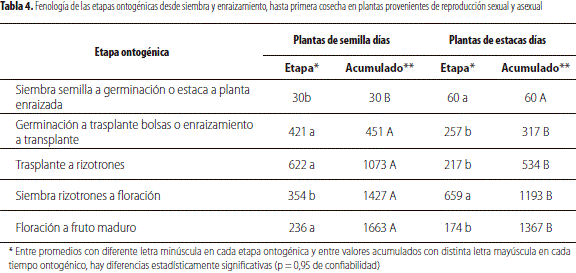
La duración del período de floración a fructificación fue superior en las plantas de origen sexual, por una mayor desuniformidad entre ellas al inicio de la floración (tabla 4). Al respecto, el tiempo reportado para el departamento de Antioquia desde la antesis floral a la cosecha, para el mortiño, fue de 165 días (Hernández 2010).
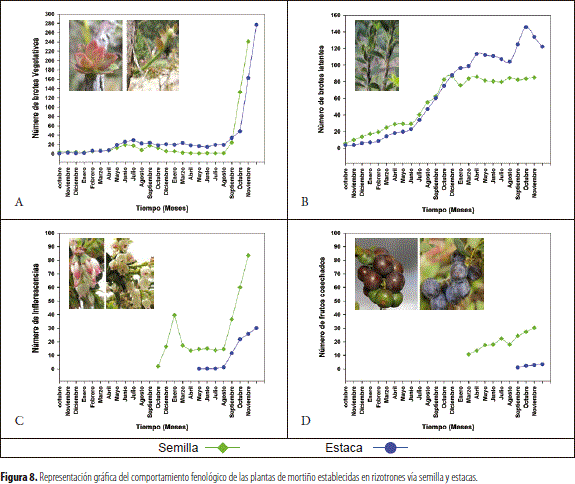
En el desarrollo de la investigación se contabilizó el número de brotes vegetativos, cuya formación se dio en forma continua a través del tiempo (figura 8) con un aumento considerable de estos a partir de los 677 días (figura 8A), tendencia que fue similar para los dos materiales. Al respecto, no se detectó un período de latencia generalizado en ambos tipos de planta.
En una población natural del oriente antioqueño se reportó que la formación de hojas ocurrió durante toda la investigación y se incrementó con la época lluviosa, la humedad relativa y la etapa de fructificación, con disminución de la floración (Gómez 2004). Este comportamiento difiere con lo reportado para muchas especies tropicales, en las cuales se ha observado que la brotación está estrechamente correlacionada con la época de menor precipitación, en la transición con la estación seca (Fournier 1976; Borchert 1980; Fournier 1986).
En especies de Vaccinium, como es el caso del arándano V. corymbosum, se presentan dos épocas marcadas en el desarrollo foliar, una de inducción y otra de desarrollo vegetativo (Bañados et al. 2007). Al respecto, en un estudio de los períodos de rebrotes de árboles tropicales se señaló que estos tienen una marcada periodicidad en extensión endógena, para luego presentar un período de rebrotes pronunciado, con hojas jóvenes de un color diferente el cual depende de la especie (Hallé et al. 1978) las cuales, en el caso de V. meridonale, son de color rojizo, por presencia de pigmentos antociánicos.
En el estudio se pudo apreciar que los brotes vegetativos, luego de su formación y elongación, entraban en un estado de latencia el cual se caracteriza por la ausencia en la formación de yemas vegetativas y el no crecimiento de estos y que, posteriormente al período de reposo, se forma un flujo nuevo de crecimiento (figura 8B).
En las especies del género Vaccinium, que se cultivan comercialmente, como es el caso del arándano (Vaccinium corymbosum), el crecimiento ocurre a través de flujos, con presencia entre uno y tres de ellos en cada temporada, cuyo número depende del vigor de la planta y del brote, al igual que de la época en la cual se suspende la fertilización nitrogenada; además, el mayor número de estos se obtiene con plantas y brotes vigorosos y en huertos con fertilizaciones nitrogenadas tardías (Bañados et al. 2007).
Los flujos de crecimiento han sido reportados en un número considerable de especies frutales como es el mango, el cual presenta entre 1, 2, o 3 períodos de crecimiento por año, proceso foliar que varía entre las plantas del mismo cultivar y aún entre las ramas de un solo individuo; en esta la formación de los brotes se reconoce por el cambio de tonalidades que tienen las hojas conforme van madurando ( Jipón 1996). Lo anterior también ha sido reportado en otras frutas como la guanábana (Annona muricata), la cual exhibe entre uno y tres flujos de crecimiento anuales (Parés-Martínez et al. 2005).
El inicio de la floración, figura 8C, se dio primero en el mes de octubre en el material proveniente de semilla, cuando se encontraban dos flujos de esta; no obstante, luego del inicio del proceso se apreciaron flores en forma continua en los dos tipos de multiplicación. Por su parte, los individuos provenientes de multiplicación vegetativa, iniciaron su floración al final de la investigación, en el mes de mayo (figura 8C).
Para la zona de influencia de la investigación se han reportado dos épocas de floración: febrero a mayo y septiembre a diciembre (Corantioquia 2003), con picos observados en febrero y agosto (Gómez 2004). Al respecto, en las Antillas, la floración de las especies de Vaccinium ocurre en estos mismos períodos (Berazain 1991).
La floración coincide con épocas secas y se asocia inversamente con el brote de hojas (Gómez 2004). La presencia de flores en esta época se puede dar como una respuesta al estrés hídrico (Céspedes 1991) y es una ventaja adaptativa, ya que al darse la producción de frutos en época lluviosa se favorece el desarrollo de las plántulas (Gómez 2004).
Como resultado de un censo fenológico en cuatro subpoblaciones de Vaccinium hirtum, realizado durante dos años, se detectó una variación dentro de estas en cuanto al período de floración, en lo cual influyó tanto el componente genético como las diferencias microambientales (Mahoro 2002). Sobre lo anterior, se ha señalado que son muchos los signos ambientales y endógenos que tienen efectos diferentes en la floración de diversas especies (Blazquez y Weigel 2000).
La fructificación se presentó primero en el material propagado por semilla (1.663 días), en el mes de marzo, y luego en el de propagación asexual (1.367 días), a partir del mes de septiembre. Al respecto, para el oriente antioqueño se han reportado en el mortiño dos períodos de fructificación: abril a junio y septiembre a diciembre (Corantioquia 2003), con información de que en las poblaciones naturales el mortiño fructifica durante todo el año, con porcentajes que oscilan entre 17 % al 39 % y la producción se correlaciona positivamente con la humedad relativa y el brote de hojas.
Cordero (2000) afirmó que es poco predecible la producción de frutos en los bosques tropicales, durante períodos definidos, con regularidad en los ciclos productivos. Por su parte, en un trabajo llevado a cabo durante diez años con Vaccinium corymbosum L., se reportó una fructificación periódica bianual (Vander y Cabilio 1996). Lo anterior podría indicar la posibilidad de comportamientos cíclicos en cuanto a la producción de frutos en el taxón Vaccinium meridionale afectados por factores climáticos diversos.
Los datos obtenidos indican que este es un cultivo que se debe planificar cuidadosamente desde la óptica del material para la siembra ya que la cosecha de frutos toma un período de tiempo considerable, lo cual no admite errores en la selección del genotipo. Esto sirve tanto para los productores como para los posibles ofertantes de material de siembra. Se consideran más apropiados para el establecimiento de cultivos plántulas clonadas, ya que son más uniformes genéticamente que las obtenidas por semilla, las cuales segregan; aspecto no recomendable en plantaciones perennes y, adicionalmente, las plantaciones establecidas a partir de multiplicación vegetativa toman menos tiempo para entrar en cosecha.
Con la investigación actual se obtuvo el primer reporte del tiempo transcurrido desde la siembra hasta el inicio de la floración, desde la plantación de la semilla y el enraizamiento de estacas, el cual fue de 1.427 días para las plantas provenientes de semilla y 1.193 para aquellas derivadas de enraizamiento de estacas, sobre lo cual no fue posible ubicar reportes previos.
Conclusiones
Las plantas de mortiño, bajo condiciones del hábitat del oriente antioqueño, toman un período considerable de tiempo desde la siembra de semillas y el enraizamiento de estacas hasta la madurez del fruto, 1.663 días en el primer caso y 1.367 en el segundo.
Lo anterior puntualiza la necesidad de una selección cuidadosa de materiales con adaptación al sitio de siembra e igualmente que reúnan los atributos de calidad y productividad requeridos para procesos productivos, al igual que para su aceptación por parte de los consumidores.
Las raíces de la especie carecen de pelos absorbentes, el cual es el primer reporte encontrado.
Tanto para los materiales obtenidos mediante propagación sexual y asexual, hubo un crecimiento continuo de la parte aérea y la raíz, sin períodos de alternancia entre las estructuras aéreas y subterráneas del vegetal.
En las plantas derivadas de clonación se observó una menor dispersión en los datos de las diversas variables registradas, que en aquellas provenientes de semilla.
En los dos tipos de materiales hay un índice alométrico, relativamente grande, entre los órganos aéreos y la raíz, los cuales fueron del orden de 6,28, en el caso de las plantas provenientes de semilla, y de 7,00, para aquellas multiplicadas en forma asexual. Esto indica la necesidad de restringir el crecimiento de los órganos aéreos a través de poda.
No se detectaron diferencias en número de brotes y altura entre los dos tipos de plantas, pero sí entre el diámetro basal del tallo y el área de copa, con mayor valor en la primera variable en las de semilla y en la segunda en las de propagación vegetativa.
Agradecimientos
Este trabajo fue financiado por el Ministerio de Agricultura y Desarrollo Rural de Colombia (MADR) mediante el proyecto Poda y Niveles de Sombrío en el Mortiño (Agraz) y Arándano y su Relación con el Comportamiento Productivo y Fenológico en Diversos Ambientes 052-2008L1363-3473.
Referencias
Alaback PB, Herman FR. 1988. Long-term response of understory vegetation to stand density in Picea-Tsuga forest. Can J For Res. 18(12):1522-1530. [ Links ]
Albert T, Raspé O, Jacquemart AL. 2003. Clonal structure in Vaccinium myrtillus L. revealed by RAPD and AFLP markers. Int J Plant Sci. 164(4):649-655. [ Links ]
Avilán L, Leal F, Bautista D. 1995. El aguacatero: principios y técnicas para su producción. Caracas: Espasande. [ Links ]
Bañados P, Donnay D, Uribe P. 2007. Poda en verde en arándanos. Agronomía y forestal. (31):16-19. [ Links ]
Berazain R. 1991. Revisión del género Vaccinium L. en las Antillas. Revista Jard Bot Nac Univ Habana. 12:3-13. [ Links ]
Bernal JA, Díaz CA. 2005. Tecnología para el cultivo del aguacate. Rionegro: Corporación Colombiana de Investigación Agropecuaria. [ Links ]
Blazquez MA, Weigel D. 2000. Integration of floral inductive signals in Arabidopsis. Nature. 404:889-892. [ Links ]
Borchert R. 1980. Phenolog y and ecolog y of tropical trees. Erytrina peoppigiana. Ecology. 39(5):1065-1074. [ Links ]
Box G, Jenkins G, Reinsel G. 1994. Time series analysis, forecasting and control. 3a ed. Englewood Cliffs: Prentice Hall. [ Links ]
Bugarínn-Montoya R, Galvis-Spinola A, Sánchez-García P, García-Paredes C. 2002. Acumulación diaria de materia seca y de potasio en la biomasa aérea total de tomate. Terra. 20:401-409 [ Links ]
Carey AB. 1995. Sciurids in Pacific Northwest managed and old-growth forests. Ecol Appl. 5(3):648-661. [ Links ]
Carey AB, Johnson ML. 1995. Small mammals in managed naturally young, and old-growth forests. Ecol Appl. 5(2):336-352. [ Links ]
Céspedes R. 1991. Fenología de Quercus seemanni Lieb. (Fagaceae), en Cartago, Costa Rica. Rev Biol Trop. 39(2):243-248. [ Links ]
Contreras MR. 2010. Efecto de la aplicación de CPPU sobre la calidad de fruta en arándano alto (Vaccinium corymbosum L.), cultivar Elliott [tesis]. [Temuco]: Universidad de la Frontera. [ Links ]
Corantioquia. 2003. Conozcamos y usemos el Mortiño. 1a ed. Medellín: Corantioquia. [ Links ]
Cordero LH. 2000. Aspectos fenológicos de algunas especies del bosque andino páramo en el norte de Boyacá. En: Desarrollo sostenible en los Andes de Colombia. Cárdenas F, editor. Bogotá: Universidad Javeriana. [ Links ]
Eriksson O. 1993. Dynamics of genets in clonal plants. Trends Ecol Evol. 8(9):313-316. [ Links ]
Fournier LA. 1976. Observaciones fenológicas en el bosque húmedo premontano de San Pedro de Montes de Oca, Costa Rica. Turrialba. 26(1):54-59. [ Links ]
Fournier LA, Herrera ME. 1986. Fenología y ecofisiología de Gliricidia sepiwn ( Jacq.) Steud, "Madero negro" en Ciudad Colón, Costa Rica. Rev Biol Trop. 34(2):283-288. [ Links ]
Galleta GJ, Ballington JR. 1996. Blueberries, cranberries and lingonberries. En: Janick J, Moore JN, editores. Fruit breeding. Volumen 2, Vine and small fruits. New York: John Wiley y Sons. pp. 1-107 [ Links ]
Garret R. 1988. Riego y poda en arándonos; [consultado 2010 jul 7]. http://www.inia.cl/medios/biblioteca/seriesinia/NR06977.pdf. [ Links ]
Gaviria CA, Ochoa CI, Hernández JD, Lobo M, Medina CI, Rojano BA. 2012. Cambios en la actividad antioxidante en frutos de mortiño (Vaccinium meridionale Sw.) durante su desarrollo y maduración. Rev Fac Nac Agron. 65(1):6487-6495. [ Links ]
Gaviria CA, Ochoa CI, Sánchez NY, Medina CI, Lobo M, Galeano PL, Mosquera AJ, Tamayo A, Lopera YE, Rojano BA. 2009. Propiedades antioxidantes de los frutos de agraz o mortiño (Vaccinium meridionale Swartz). En: Ligarreto GA, editor. Perspectivas del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en la zona altoandina Colombiana. Bogotá: Universidad Nacional de Colombia. pp. 93-112. [ Links ]
Givnish TJ. 1988. Adaptation to sun and shade: a whole-plant perspective. Aust J Plant Physiol. 15:63-92. [ Links ]
Gómez CP. 2004. Autoecología del mortiño (Vaccinium meridionale Sw.). [tesis de maestría]. [Bogotá]: Universidad Nacional de Colombia. [ Links ]
Hallé F, Oldeman RAA, Tomlinson PB. 1978. Tropical trees and forests: an architectural analysis. Berlin: Springer. [ Links ]
Hallé MG, Orcutt DM. 1978. The physiolog y of plants under stress. Nueva York: John Wiley & Sons. [ Links ]
Halpern CB, Spies TA. 1995. Plant species diversity in natural and managed forest of the pacific northwest. Ecol Appl. 5(4):913-934. [ Links ]
Hamrick JL, Godt MJ. 1989. Allozyme diversity in plant species. En: Brown AHD, Clegg MT, Kahler AL, Weir BS, editores. Plant population genetics, breeding and genetic resources. Sunderland: Sinauer Associates. pp. 43-63. [ Links ]
Harper JL. 1981. The concept of populations in modular organisms. En: May RM, editor. Theoretical ecology: principles and applications. Oxford: Blackwell Scientific Publications. pp. 53-77. [ Links ]
Hernández JD. 2010. Caracterización físico química del fruto de mortiño (Vaccinium meridionale Sw) [tesis]. [Medellín]: Universidad Nacional. [ Links ]
Jipón P. 1996. Cultivo de mango. San José de Costa Rica: For-Export. [ Links ]
Kawamura K, Takeda H. 2004. Rules of crown development in the clonal shrub Vaccinium hirtumin a low-light understory: a quantitative analysis of architecture. Can J Bot. 82(3):329-339. [ Links ]
Kawamura K, Takeda H. 2002. Light environment and crown architecture of two temperate Vaccinium species: inherent growth rules versus degree of plasticity in light response. Can J Bot. 80(10):1063-1077. [ Links ]
Krewer G, Ruter JM, Nesmith DS, Thomas DL, Sumner P, Harrison K, Westberry G, Mullinix B, Knox D. 1997. Preliminary report on the effect of tire chips as a mulch and substrate component for blueberries. Acta Hort. 446:309-318. [ Links ]
León J. 2000. Botánica de los cultivos tropicales. 3a ed. San José de Costa Rica: IICA. [ Links ]
Lewis AM. 1997. Plant stems: physiology and functional morphology. Tree Physiol. 17(8-9):603-604 [ Links ]
Mahoro S. 2002. Individual flowering schedule, fruit set, and flower and seed predation in Vaccinium hirtum Thunb. (Ericaceae). Can J Bot. 80(1):82-92. [ Links ]
Matteucci SD, Morello J. 2001. Aspectos ecológicos del cultivo de la coca. Revista Encrucijada. 1(8):82-91 [ Links ]
Medina CI. 2010. Variabilidad poblacional y ecofisiológica del mortiño (Vaccinium meridionale Sw.), especie con potencial productivo y agroexportador en el tropicoaltoandino [tesis doctoral]. [Medellín]: Universidad Nacional de Colombia. [ Links ]
Michigan State University. 2003. Blueberry growth stages; [consultado 2007 dic 18]. http://www.blueberries.msu.edu/pdf/growthstages.pdf. [ Links ]
Niklas KJ, Enquist BJ. 2002. Canonical rules for plant organ biomass partitioning and annual allocation. Am J Bot. 89(5):812-819. [ Links ]
Parés-Martínez J, Arizaleta M, Bautista D. 2005. Crecimiento y topología de la ramificación de la guanábana y el manirote. Pesq Agropec Bras. 40(9):867-872. [ Links ]
Poorter H, Nagel O. 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Aust J Plant Physiol. 27(6):595-607. [ Links ]
Poorter L, Werger MJ. 1999. Light environment, sapling architecture, and leaf display in six rain forest tree species. Am J Bot. 86(10):1464-1473. [ Links ]
Retamales JB, Lobos GA. 2008. Sistema radical de arándanos y su interacción con la nutrición vegetal En: Muñoz C, editor. El arándano en Chile. [ Links ]
Romero R. 1991. Frutas silvestres de Colombia. Bogotá: Instituto de Cultura Hispánica. [ Links ]
Shibles R. 1987. Crop physiolog y. Iowa: Iowa State University. [ Links ] Stenström A, Jónsdóttir IS, Augner M. 2002. Genetic and environmental effects on morpholog y in clonal sedges in the Eurasian Arctic. Am J Bot. 89(9):1410-1421. [ Links ]
Taiz L, Zeiger E. 1991. Plant physiology. San Francisco: Benjamin Cummings. [ Links ]
Tekalign, T, Hammes. PS. 2005. Growth and productivity of potato as influenced by cultivar and reproductive growth: II. Growth analysis, tuber yield and quality. Sci Hort. 105(1):29-44. [ Links ]
Torres WS, Montoya IA, Ligarreto GA. 2009. Aspectos sociales y económicos de la producción de agraz o mortiño (Vaccinium meridionale Swartz). En: Ligarreto GA, editor. Perspectivas del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en la zona altoandina colombiana. Bogotá: Universidad Nacional de Colombia. pp. 113-134. [ Links ]
Torrico JC, Cardona JO. s. f. Ganadería Ecológica. Guía para las buenas prácticas ganaderas. Experiencias en el Sumapaz-Colombia. [ Links ]
Valladares FE, Martinez-Ferri E, Balaguer L, Perez-Corona E, Manrique E. 2000. Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: a conservative resource-use strategy? New Phytol. 148(1):79-91. [ Links ]
Vander SP. 2004. Vaccinia gloriosa. Small Fruits Rev. 3(3/4):221-227. [ Links ]
Vander SP, Cabilio P. 1996. Ten year study of annual variation in berry and seed production in a population of Vaccinium corymbosum L. Am Midl Nat. 135(2):349-356. [ Links ]
Vanninen P, Mäkelä A. 1999. Fine root biomass of scots pine stands differing in age and soil fertility in southern Finland. Tree Physiol. 19(12):823-830. [ Links ]
Widén B, Cronberg N, Widén M. 1994. Genotypic diversity, molecular markers and spatial distribution of genets in clonal plants, a literature survey. Folia Geobot Phytotax. 29(2):245-263. [ Links ]
