










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
versão impressa ISSN 0122-8706
Corpoica cienc. tecnol. agropecu. vol.16 no.1 Mosquera jan./jun. 2015
1 Agronomic engineer, MSc. Profesional support ICA. Medellín, Colombia. luisa.torres@ica.gov.co
2 Agronomic engineer, PhD. Full professor. Universidad Nacional de Colombia. Medellín, Colombia. jrcartag@unal.edu.co
3 Forest engineer PhD. Associate professor. Universidad Nacional de Colombia. Medellín, Colombia. gcorrea@unal.edu.co
4 Agronomic engineer. Researcher Corpoica-CI El Nus. San Roque, Colombia. dmonsalveg@corpoica.org.co
5 Agronomic engineer, MSc. Researcher Corpoica-CI La Selva. Rionegro, Colombia. mlondono@corpoica.org.co
Fecha de recepción: 15/09/2014. Fecha de aceptación: 12/02/2015
Abstract
The effect of environmental factors on populations of Monalonion velezangeli Carvalho & Costa was studied in avocado cv. Hass planted in the Colombian Andes. Climate data for the 2010-2011 period were obtained from six automatic weather stations. Insect activity was monitored by recording the presence of M. velezangeli individuals and fresh damage signals on 12 tagged branches per tree, oriented towards the four cardinal points at low, medium and high tree strata. Relations were established between population indexes and temperature, relative humidity and wind speed by means of Biplot representations and correlation analyses. These analyses were carried out not only between records taken the same day, but also included backward cumulative values of climatic factors, i.e., the sum over a period of days before the assessment. The results suggest that temperature and relative humidity have a direct influence on the insect, while wind speed showed an inverse relationship. This study illustrates how the analysis of climate information can improve our predictive capacity on the establishment of M. velezangeli in avocado crops, which is a key aspect of integrated pest management.
Key words: Abiotic factors, weather variables, population dynamics, pest monitoring
Resumen
Se estudió el efecto de los factores ambientales sobre la población de Monalonion velezangeli Carvalho & Costa en aguacate cv. Hass, plantado en los Andes colombianos. Se obtuvieron datos de clima correspondientes al período 2010-2011, usando seis estaciones meteorológicas automáticas. La actividad del insecto fue monitoreada mediante el registro de individuos de M. velezangeli y la presencia de daño fresco en doce ramas marcadas en cada árbol, ubicadas en los cuatro puntos cardinales, en los estratos bajo, medio y alto. Se establecieron relaciones entre los índices de población obtenidos con valores de temperatura, humedad relativa y velocidad del viento, mediante representaciones Biplot y análisis de correlación. Estos análisis se hicieron no solo con los registros tomados el mismo día, sino que incluían también los valores retroacumulados de los factores climáticos, esto es, la suma durante un período de días antes de la evaluación. Los resultados sugieren que la temperatura y la humedad relativa tenían una influencia directa sobre el insecto, mientras que la velocidad del viento tenía una relación inversa. Este estudio ilustra cómo el análisis de la información climática puede mejorar la capacidad predictiva sobre el establecimiento de M. velezangeli en cultivos de aguacate, un aspecto clave para direccionar el manejo integrado de la plaga.
Palabras claves: Factores abióticos, variables de clima, dinámica de poblaciones, monitoreo de plagas
Introduction
Avocado (Persea americana Mill.) cv. Hass is a fruit of great interest in international markets due to the ease of transportation -even across long distances- and to its nutraceutical and sensory attributes (Ortega 2003; Rojas et al. 2004). In Colombia, there are approximately 9,000 hectares of avocado cv. Hass distributed in the departments of Antioquia, Caldas, Cauca, Cundinamarca, Quindío, Risaralda, Santander, Tolima and Valle del Cauca (Palacio 2013). International markets, especially those of the United States and Europe, demand innocuous products with good sanitary quality. This is a challenge for growers who intend to harvest their products under clean production and high quality standards. This, in turn, implies an intelligent management of health problems both in pre and post-harvest, to assure the acceptance of Colombian avocado in consumer countries.
One of the sanitary limitations faced by farmers who produce avocado cv. Hass in Colombia is the attack of the bug Monalonion velezangeli Carvalho & Costa (Hemiptera:Miridae). This sucking insect affects the internal and external quality of the fruit, as well as the inflorescence and vegetative buds, causing losses of up to 42% of the production (Londoño 2012). In this regard, Torres et al. (2012) found that although M. velezangeli feeds on both vegetative and reproductive structures, it shows preference for the latter.
Climatic conditions may have important effects on insects both at the individual and the population levels. Climate may have direct influence on insects, either on their development and growth rate or as a mortality factor. Herbivorous insects must adapt to the host plant and the environment to successfully complete their life cycle (Bale et al. 2002; Wylie and Speight 2012).
Insects require a favorable range of climatic conditions for optimum development and reproduction. Out of this range, such processes are hindered or suppressed. Moisture, temperature, wind, rain and radiation are among the environmental factors that may influence the growth, development, incidence, distribution and dispersal of herbivorous insects (Bale et al. 2002; Thomson et al. 2010). In this respect, Hodkinson (2009) observed that the main factors driving the adaptation of insects to the environment are temperature and water availability.
Temperature is one of the most influential and thoroughly studied climate factors affecting the development and reproduction of insects. It may induce changes in herbivorous insect population density, dynamics and genetic composition, and also in the life span, dispersal ability, individual size and consumption patterns. These effects differ across species and at the same time are influenced not only by specific life cycle features and adaptive abilities, but also by the environment (Bale et al. 2002; Wylie and Speight 2012). Given that insects are sensitive to temperature changes, their metabolic rate tends to double with temperature increases of 10 °C. This, in turn, accelerates voracity, development and general activity, influencing population dynamics through effects on fertility, voltinism, survival and dispersal (Karuppaiah and Sujayanad 2012).
Rain affects insects in several ways. It directly satisfies essential water requirements for development and reproduction and indirectly provides increased food availability (Pellegrino et al. 2013). On the other hand, water droplets may cause physical damage to insects, while increased moisture not only raises the probability of diseases, but also reduces body temperature as a result of mistiness and the cooling effect of water (Wylie and Speight 2012).
As a microclimatic variable, relative humidity combines temperature and precipitation, which is the reason why it has a remarkable impact on arthropod populations. According to Child (2007) and to Wylie and Speight (2012), insects are susceptible to desiccation, while general increases in air moisture levels favor their reproduction, development and growth rates, as well as their dispersal capabilities; hence, they prefer considerably damp areas.
Wind is an important climate factor in insect migration and in their active and passive dispersal (Westbrook and Isard 1999; Wylie and Speight 2012). Small air movements can constitute necessary chemical signal channels for adult mating or for locating host plants (Blackmer and Cañas 2005; Wylie and Speight 2012). However, while some species find it difficult to fly or control their direction under the influence of strong winds (Wood, 2007), some others fly more easily under these conditions (Sridhar et al. 2012).
According to Bale et al. (2002), the effects of climate change on herbivorous insects can be immediate -through impacts on their physiology and behavior- or collateral -when insect responses are mediated by other factors such as host plants-. Provided that climate factors, mainly temperature, may influence compatibility with host plants, potential harm caused by most insect pests depends on their abundance and on the moments when crops are more susceptible to attacks (Bale 1991). Dwelling in different habitat types, polyphagous insects are capable of occupying a wider range of latitudes and altitudes. As a consequence, they are less likely to be negatively affected by climate change (Bale et al. 2002).
The design of comprehensive strategies for the prevention and management of M. velezangeli calls for understanding these relationships between insects and climate factors in orchards of avocado cv. Hass.
To date, the activity of M. velezangeli associated with climatic factors in Colombia was unknown. Hence, the aim of this study was to relate the presence of M. velezangeli with climate factors, as a contribution to the management of this insect.
Materials and methods
The study was carried out between September 2010 and August 2011 in six farms producing avocado cv. Hass with some records of damage by the pest. A WatchDog series 2000® weather station was installed in each of the six farms, in order to measure the following climatic variables: ambient, leaf and soil temperatures; relative humidity; leaf and soil moisture; precipitation; solar radiation; photosynthetically active radiation; wind speed and direction; wind gust and dew point.
In the six farms, we assessed the incident population of M. velezangeli and the number of fresh injuries caused by the insect (brown reaction areas with wet red exudate and lumpy consistency), approximately every 20 days. Eleven evaluations per farm were carried out in the orchards located in eastern Antioquia (El Cebadero, El Guarango, La Miranda and Persea), whereas in those located in the Coffee Zone (El Jordánand Chile), we performed 14 evaluations per farm, for a grand total of 72 assessments.
As to data collection, 20 trees between 4 and 8 years old were randomly chosen at each orchard. Twelve terminal branches were labeled in each tree, covering the four cardinal points across the three strata (low, medium and high). Thus, the final sampling unit was a branch pointing at a cardinal direction in a specific layer and farm at a given time. For each sample, we recorded the manifestation of the plague expressed as the number of nymphs and adults and the number of fresh damage signals observed on the 30 cm-distal-segment of the branch (M. velezangeli + fresh damage). Since the insect was visible to the naked eye, and the damage caused by this could be directly observed, monitoring did not require amplification instruments. In all cases, visual assessments were conducted with special attention to the tips of the leaves and floral buds.
Some exploratory Biplot representations prepared in Matlab® software package allowed analyzing the relationship of the studied climate factors and the presence and damage caused by M. velezangeli at each farm and time. These Biplots not only included the climatic conditions on the day when the assessment took place, but also a backward cumulative representation of each of the climatic variables, from the day of the assessment, with backward increments of one day, to the 30 previous days. The kth backward cumulative value of a variable was calculated as the sum of the values registered for that variable, day by day, from the kth day before the day of the assessment to the day of the visit. In the database elaborated this way, the day 0 corresponded with the raw non-cumulative version of the variable. We used this analysis instead of a time series one, because of the flexibility to correlate the dynamics of the insect with the climatic conditions of a period rather than the usual point by point correlation.
Once the climatic variables that showed the greatest absolute relationships with the presence and damage caused by M. velezangeli had been isolated, we calculated the Pearson correlation coefficient for each of these variables in their raw form as well as in their backward cumulative versions. This was done to check for any effect of the previous climatic conditions on the response variable.
Results and discussion
The analysis of the influence of the climatic conditions previous to the evaluation date showed that the Cumulative Minimum ambient temperature, the Cumulative Minimum leaf temperature and the Cumulative Minimum relative humidity have a positive correlation with the presence and damage caused by M. velezangeli. This correlation reached a maximum in correspondence with the backward cumulative data of the 12 days prior to the assessment (Pearson's r Cum Min AT = 0.37; Pearson's r Cum Min LT = 0.31; and Pearson's r Cum Min RH = 0.14, respectively), indicating that the activity of M. velezangeli was favored when the ambient and leaf minimum temperatures and also the minimum humidity records throughout the 12 days previous to the evaluation were not extremely low (Figures 1 and 3).
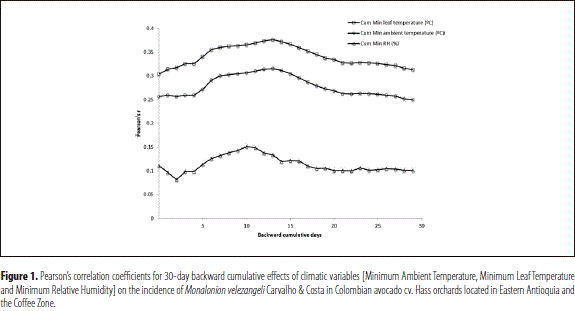
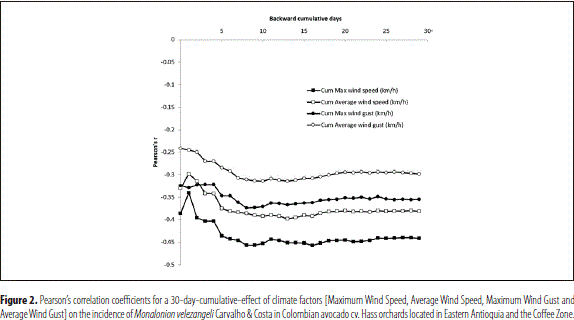
Conversely, there was a negative correlation between the variables Maximum wind speed, Average wind speed, Maximum wind gust and Average wind gust and the presence of M. velezangeli, reaching an absolute maximum in correspondence with the cumulative data of the 13 days previous to the assessment (Pearson's r Max WS = - 0.45; Pearson's r AWS = -0.40; Pearson's r Max WG = - 0.37; and Pearson's r AWG = - 0.31); all of which indicates that high gust and wind speeds diminished the incidence of the insect (Figures 2 and 3).
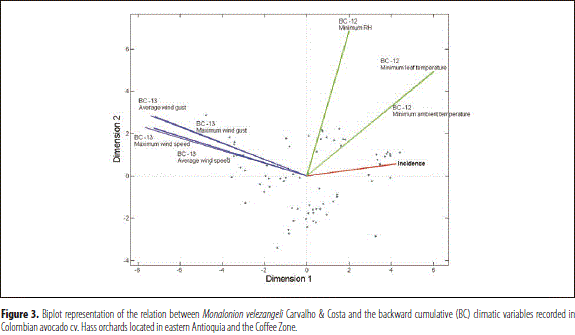
The above results show that the presence, damage, development and feeding activity of M. velezangeli in avocado cv. Hass orchards is favored by warm, moist, static environments; these effects being more decisive when the weather conditions are consistent for more than a week.
The described scenario is summarized by a Biplot representation including the incidence of M. velezangeli and the backward cumulative climatic variables that showed the greatest absolute correlations with incidence (Figure 3).
According to Alberts (2010) and Ju et al. (2011), insects are poikilothermic animals. Hence, they are strongly affected by several climate factors. Wylie and Speight (2012) argue that changes in ambient temperature affect insects' internal temperature and, consequently, their metabolic rate. This is consistent with the concept expressed by Child (2007) in the sense that, for most pest insects, temperatures above 15 °C and up to 35 °C increase energ y levels, which leads to greater mobility and higher feeding, reproduction and egg production rates, coupled to lower mortality.
A study conducted by Alberts (2010) on a Lygus sp. (Miridae) in South African avocado groves found that when minimum temperatures are low, the activity and number of individuals of this bug is diminished, as reflected in reduced insect feeding; whereas its activity increases with rising temperatures. Similarly, Rodríguez (2011) found that populations of Collaria scenica (Hemiptera: Miridae) increase when minimum temperatures are above average and evaporation is low.
Provided that the pest is susceptible to drought, this data show how its development is favored under greater water availability.
Relative humidity affects the development and survival of insects, which must keep body water content within certain limits, also influenced by the degree of permeability of the cuticle (Raghu et al., 2004). In this respect, Brenes et al. (2009) suggest that damage caused by herbivorous insects increase under elevated moisture conditions. Speight and Wylie (2012) utter that most tropical herbivorous insects are more abundant and voracious at the beginning of the rainy season which, as a consequence, appears as favorable to pest populations. Villacorta (1977) reports that high relative humidities promoted the presence of M. annulipes (Hemiptera: Miridae) in Costa Rican cocoa plantations. Yanhui and Kongming (2011) have observed that high relative humidity (70-80%) is directly related to increasing number of eggs, nymphal survival, longevity and fecundity in females of Apolygus lucorum (Hemiptera: Miridae), finally resulting in population growth. The above mentioned Miridae population growth under warm temperature and high relative humidity conditions is similar to that observed in the population of M. velezangeli evaluated in this study.
The wind is known to be a factor of dissemination of arthropod pest populations (Westbrook and Isard, 1999; Wood, 2007; Sridhar et al., 2012; Wylie and Speight, 2012), but above certain limits, wind speed reduces insect abundance by inducing strenuous flight (Turner, 2006; Møller, 2013). Symmons and Luard (1982) and Kakitani et al. (2003) suggest that winds faster than 3 km h-1 combined with low temperatures may also limit insect flight. Additionally, limited flight capacity has been observed in M. velezangeli adults (Ramírez et al., 2008) and other species of the family Miridae attacking avocado such as Dagbertus minensis (Yarita and Cisneros, 2010), which is consistent with the negative correlation found in the current study between the presence of M. velezangeli and increasing wind speed.
It was not our pretension to carry out a comprehensive research on the relationships between M. velezangeli and the climatic factors. Moreover, we are aware that this research only provides a partial explanation on the dynamics of M. velezangeli, leaving aside other factors that could affect it. Nonetheless, this research may be taken as a starting point by other researches on this subject.
Conclusions
M. velezangeli population size and damage in orchards of avocado cv. Hass are promoted by moderated temperature and relative humidity and hampered by strong winds.
The study of the influence of abiotic factors on the behavior of M. velezangeli stresses the need to record minimum necessary weather information for several years, so that it can be used in the design of an integrated pest management strategy.
Acknowledgements
The authors want to express their gratitude to CorporaciónColombiana de InvestigaciónAgropecuaria (Corpoica) and to Universidad Nacional de Colombia- Medellin campus, for contributing to the formation of useful human talent in the development of the current study; and to the Ministry of Agriculture and Rural Development for its financial support through the project "Development of strategies for the prevention and management of Monalonion velezangeli Carvalho & Costa". Just as well, we thank the avocado growers that allowed the conduction of the current study in their orchards.
References
Alberts AJ. 2010. Defining economical action levels for avocado bugs (Miridae) and the use of area wide IPMK to warn growers of possible outbreaks. South African Avocado Growers' Association Yearbook. 33:66-69. [ Links ]
Bale JS. 1991. Insects at low temperature: a predictable relationship? Funct Ecol. 5(2):291-298. [ Links ]
Bale J, Masters G, Hodkinson I, Awmack C, Bezemer M, Brown V, Butterfield J, Buse A, Coulson J, Farrar J, et al. 2002. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Glob Change Biol. 8(1):1-16. [ Links ]
Blackmer J, Cañas L. 2005. Visual cues enhance the response of Lygus hesperus (Heteroptera: Miridae) to volatiles from host plants. Environ Entomol. 34(6):1524-1533. [ Links ]
Brenes-Arguedas T, Coley PD, Kursar TA. 2009. Pests vs. drought as determinants of plant distribution along a tropical rainfall gradient. Ecology. 90(7):1751-1761. [ Links ]
Child RE. 2007. Insect damage as a function of climate. In: Padfield T, Borchersen K, editores. Museum Microclimates. Copenhagen: National Museum of Denmark. pp. 57-60. [ Links ]
Hodkinson ID. 2009. Life cycle variation and adaptation in jumping plant lice (Insecta: Hemiptera: Psylloidea): a global synthesis. J Nat Hist. 43(1-2):65-179. [ Links ]
Ju RT, Wang F, Li B. 2011. Effects of temperature on the development and population growth of the sycamore lace bug, Corythucha ciliata. J Insect Sci. 11(16):1-12. [ Links ]
Kakitani I, Ueno HM, Forattini PP. 2003. Paridade e influência do vento sobre a freqüência de Anopheles marajoara, São Pablo. Rev SaúdePública. 37(3):280-284. [ Links ]
Karuppaiah V Sujayanad GK. 2012. Impact of climate change on population dynamics of insect pests. World J Agric Sci. 8(3):240-246. [ Links ]
Londoño M. 2012. Informe técnico final proyecto Desarrollo de Estrategias para la Prevención y Manejo de Monalonion velezangeli Carvahlo & Costa. Rionegro: Corpoica-CI La Selva. [ Links ]
Lu YH, Wu KM. 2011. Effect of relative humidity on population growth of Apolygus lucorum (Heteroptera: Miridae). Appl Entomol Zool. 46(3):421-427. [ Links ]
Møller AP. 2013. Long-term trends in wind speed, insect abundance and ecology of an insectivorous bird. Ecosphere. 4(1):1-11. [ Links ]
Ortega M. 2003. Valor nutrimental de la pulpa fresca de aguacate Hass. En: Consejería de Agricultura y Pesca. Actas V Congreso Mundial del Aguacate. Málaga: Servicio de Publicaciones y Divulgación. pp. 741-748. [ Links ]
Pellegrino AC, Peñaflor MFGV, Nardi C, Bezner-Kerr W, Guglielmo CG, Bento JMS, McNeil JN. 2013. Weather forecasting by insects: modified sexual behaviour in response to atmospheric pressure changes. PLoS ONE. 8(10):e75004. [ Links ]
Portalfrutícola. 2013. 6 mil hectáreas de palta Hass entran en producción en Colombia. Santiago: Portalfrutícola; [accesed 2014 feb]. http://www.portalfruticola.com. [ Links ]
Raghu S, Drew RAI, Clarke AR. 2004. Influence of host plant structure and microclimate on the abundance and behavior of a tephritid fly. J Insect Behav. 17(2):179-190. [ Links ]
Ramírez-Cortés HJ, Gil-Palacio ZN, Benavides-Machado P, Bustillo-Pardey AE. 2008. Monalonion velezangeli la chinche de la chamusquina del café. Avances Técnicos Cenicafé No. 367. Chinchiná: Cenicafé [ Links ].
Rodríguez AO. 2011. Desarrollo de un sistema de alertas agroclimáticas tempranas para la chinche de los pastos, Collaria scenica, en la sabana de Bogotá [magister thesis] [Bogotá]: Universidad Nacional de Colombia. [ Links ]
Rojas JM, Peñuela AE, Gómez CR, Aristizábal GE, Chaparro MC, López JA. 2004. Caracterización de los productos hortofrutícolas colombianos y establecimiento de las normas técnicas de calidad. Chinchiná: Cenicafé [ Links ].
Sridhar V, Jayashankar M, Vinesh LS. 2012. Population dynamics of Zoophytophagous mirid bug, Nesidiocoris tenuis (Reuter) (Heteroptera: Miridae) and its prey, Bemisia tabaci Genn. (Homoptera: Aleyrodidae) on tomato (Solanum lycopersicum Mill). Pest Manag Hort Ecosyst. 18(1):35-38. [ Links ]
Symmons PM, Luard EJ. 1982. The simulated distribution of night-flying insects in a wind convergence. Aus J Zool. 30(2):187-198. [ Links ]
Thomson LJ, Macfadyen S, Hoffmann AA. 2010. Predicting the effects of climate change on natural enemies of agricultural pests. Biol Control. 52(3):296-306. [ Links ]
Torres LF, Correa GA, Cartagena JR, Monsalve DA, Londoño ME. 2012. Relationship of Monalonion velezangeli Carvalo & Costa (Hemiptera: Miridae) with the phenolog y of avocado (Persea americana Mill., cv. Hass). Rev Fac Nal Agr. 65(2):6659-6665. [ Links ]
Turner AK. 2006. The barn swallow. London: T & A D Poyser. [ Links ]
Villacorta A. 1977. Fluctuación anual de las poblaciones de Monalonion annulipes Sig. y su relación con la muerte descendente de Theobroma cacao en Costa Rica. An Soc Entomol Bras. 6(2):215-223. [ Links ]
Wood CR. 2007. The biometeorolog y of high-altitude insect layers [PhD thesis]. Reading : University of Reading. [ Links ]
Westbrook JK, Isard SA. 1999. Atmospheric scales of biotic dispersal. Agric For Meteorol. 97(4):263-274. [ Links ]
Wylie R, Speight M. 2012. Insect pest in tropical foresty. 2nd edition. London: CABI Publishing. [ Links ]
Yarita Y, Cisneros FH. 2010. Ciclo biológico y morfología de Dagbertus minensis Carv. & Fontes (Hemiptera: Miridae), en palto var. Hass, en la irrigación Chavimochic, Perú. Rev Peru Entomol. 46(1):15-19. [ Links ]
