










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
Print version ISSN 0122-8706
Corpoica cienc. tecnol. agropecu. vol.16 no.2 Mosquera July/Dec. 2015
Artículo de investigación
Crecimiento en vivero de las palmas aceiteras Elaeis oleífera x Elaeis guineensis y Elaeis guineensis x Elaeis guineensis en Tumaco Colombia
Growth in Tree Nursery of Oil Palms Elaeis oleifera x Elaeis guineensis and Elaeis guineensis x Elaeis guineensis in Tumaco, Colombia
Lizardo Norbey Ibarra-Ruales,1 Rafael Reyes-Cuesta2
1 Ingeniero Agrónomo. Universidad Nacional de Colombia Sede Palmira. Asistente de Investigación, Cenicafé. Chinchiná, Colombia. lizardo.ibarra@cafedecolombia.com
2 PhD, Universidad de Costa Rica. Investigador PhD, Corpoica. Palmira, Colombia. rreyes@corpoica.org.co
Fecha de recepción: 30/09/2013 Fecha de aceptación: 8/07/2015
Para citar este artículo: Ibarra-Ruales LN, Reyes-Cuesta R. 2015. Crecimiento en vivero de las palmas aceiteras Elaeis oleífera x Elaeis guineensis y Elaeis guineensis x Elaeis guineensis en Tumaco Colombia. Corpoica Cienc Tecnol Agropecu. 16(2):239-250
Resumen
El conocimiento del crecimiento en vivero de una especie vegetal perenne es básico para desarrollar prácticas de manejo agronómico y la selección de las plantas que se llevarán a campo. Esta información es requerida para los híbridos tipo OxG de palmas aceiteras, en condiciones de las regiones donde se están cultivando en Colombia. El crecimiento en vivero de las palmas aceiteras híbrido OxG F1 (Elaeis oleífera x Elaeis guineensis) y Ténera DxP (Elaeis guineensis x Elaeis guineensis) se evaluó en dos suelos con diferente retención de humedad y espacio aéreo. Las plantas del material OxG presentaron mayor acumulación de biomasa en la mayoría de sus órganos constituyentes, que las plantas del material DxP. Asimismo, presentaron diferencias en el orden de preferencia de distribución y acumulación de asimilados, en sus órganos base peciolar, tallo y peciolo. El material OxG mostró la característica diferencial respecto al DxP de poder destinar mayor proporción de sus asimilados para formar sistema radical en la planta. Por otra parte, se determinó que, para la etapa de vivero, en condiciones de suelos con alta retención de humedad y baja aereación, las plantas del material OxG presentan mejor crecimiento y desarrollo que las plantas del material DxP. Adicional-mente, se identificaron y seleccionaron tres características morfológicas con capacidad discriminante, para diferenciar el comportamiento del crecimiento de los dos tipos de palmas aceiteras respecto al suelo empleado en el vivero: área sección del peciolo (PxS), largo del raquis (LR) y emisión foliar (EF).
Palabras clave: Elaeis guineensis, híbridos, fisiología vegetal, viveros, biomasa, Tumaco-Colombia.
Abstract
The knowledge of the growth in tree nursery of perennial plant species is important in order to develop good agronomical practices for the selection of the plants that will be taken to the fields. This information is required for the hybrids OxG of oil palms under the conditions of the palm growing Colombian region. In two soils with different moisture retention and air space, the growth and development in nursery of hybrid oil palms OxG F1 (Elaeis oleifera x Elaeis guineensis) and Tenera DxP (Elaeis guineensis x Elaeis guineensis) were evaluated. Plants of OxG material were larger and accumulated more biomass in most of its constituent bodies in comparison with plants of DxP material. Likewise, both plants were quite different in the order of preference distribution and accumulation of assimilated species at the petiole basis, stem and petiole. In comparison with DxP, the OxG material showed the differential characteristic to allocate more of their assimilated species growth the plant root system. Moreover, it was determined that OxG plants exhibit better growth and development compared with DxP plants in nursery stage with high moisture retention and low aeration soils. Additionally, we identified and selected three morphological features with discriminatory authority to differentiate the growth behavior of the two types of oil palms regarding the soil used in the nursery: section area of the petiole (PxS), rachis length (RL) and leaf emission (LE).
Key words: Elaeis guineensis, Hybrids, Plant physiology, Plant nurseries, Biomass, Tumaco-Colombia.
Introducción
El crecimiento se manifiesta como un aumento irreversible del tamaño o la masa de un organismo vivo. En las plantas, está condicionado por la distribución de asimilados entre las partes que la conforman (Hunt 2003; Salisbury y Ross 2000; Villalobos et al. 2001). El monitoreo de patrones del crecimiento, en etapas tempranas de desarrollo de especies explotadas comercialmente, contribuye a determinar la calidad de las plantas que se llevarán a campo, lo que resulta muy importante cuando se trata de especies perennes como las palmáceas (Bulgarelli et al. 1993; Coto et al. 2002; Reyes et al. 2008).
Para el crecimiento y desarrollo de las plantas de la palma aceitera africana (Elaeis guineensis), la utilización del sustrato adecuado en la etapa de vivero tiene gran influencia, lo que permite obtener plantas de buena calidad para llevar a campo, acortar los periodos de inmadurez y elevar sus producciones iniciales de racimos en sitio definitivo (Chee et al. 1998; Umaña et al. 1990; Reyes et al. 2009).
El deterioro de alrededor de 30.000 ha de plantaciones de palmas E. guineensis, en Tumaco, Colombia, como consecuencia del efecto del disturbio pudrición de cogollo (PC), ha llevado a la renovación de ese cultivo con materiales del híbrido OxG, obtenidos mediante el cruzamiento de la palma aceitera americana (Elaeis oleífera) con la palma aceitera africana (E. guineensis), considerados tolerantes a ese disturbio (Corredor et al. 2008; Hartley 1988; Meunier 1991; Zambrano 2004; Bastidas et al. 2010). Sin embargo, es escasa la investigación disponible para el establecimiento y manejo de estos nuevos materiales en la región, en las etapas de vivero y campo. Por ello, el manejo dado a palmas del híbrido OxG es similar al utilizado para palmas E. guineensis, cuyas características genotípicas, fenotípicas y de comportamiento en cultivo, son diferentes al de las palmas E. guineensis cultivadas en la región.
Actualmente, para la palma aceitera E. guineensis, se dispone de estudios de crecimiento determinados en África, Asia y América (Breure 2003; Bulgarelli et al. 1993; Corley et al. 1971; Corley y Tinker 2003; Reyes et al. 2008), pero son escasos para E. oleífera y sus híbridos OxG con E. guineensis (Contreras 1996; Corley y Tinker 2003; Taylor et al. 2008). Específicamente para Tumaco, se dispone de algunos estudios sobre el crecimiento y desarrollo en vivero de materiales E. guineensis en diferentes tipos de suelo (Reyes et al. 2008 y 2009), sin embargo, no se han encontrado trabajos para materiales del híbrido OxG.
El presente estudio tuvo por objetivo evaluar, en condiciones de Tumaco, el crecimiento y desarrollo en vivero del híbrido OxG comparado con el de E. guineensis, sembrados en dos tipos de suelo de uso común en palma aceitera, este conocimiento resulta básico y necesario para el manejo agronómico y la selección en vivero de plantas del material OxG.
Materiales y métodos
El estudio se realizó en el Centro de Investigación El Mira de Corpoica, ubicado en el municipio de Tumaco (Nariño, Colombia), a 1º 31' 58" LN y 78º 41' 21" LO, región de características agroecológicas de bosque húmedo tropical (bh-T), que presenta una altura de 16 msnm, temperatura promedio anual de 25,5 ºC, precipitación acumulada de 3.067 mm año-1, humedad relativa promedio de 88 %, brillo solar acumulado de 1.008 horas sol año-1 y radiación acumulada anual de 15.080 J/cm2 (Reyes 2012).
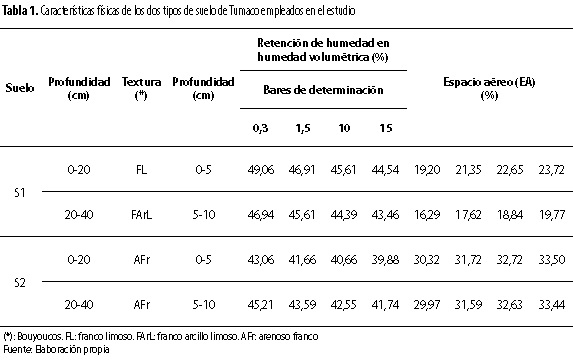
Se evaluaron plantas del híbrido OxG F1 (Elaeis oleífera x Elaeis guineensis) y el material Ténera DxP (Elaeis guineensis tipo Deli x Elaeis guineensis tipo Yangambi) de palma aceitera producidos por Corpoica (Bastidas et al. 2006; Preciado et al. 2011). Dichas plantas fueron sembradas en dos suelos de características físicas contrastantes; uno franco arcillo limoso (S1) y el otro arenoso franco (S2), el primero con mayor retención de humedad y menor espacio aéreo que el segundo (tabla 1); suelos contenidos en materos plásticos con capacidad de 24.740 cm3 (30 cm de diámetro x 35 cm de alto). Dichos materos fueron dispuestos en cuadro, a una distancia de 1,2 m, de acuerdo a un diseño experimental completamente al azar con arreglo factorial de 2x2 (suelos x materiales) con cinco repeticiones, donde cada repetición correspondió a un matero con su respectiva palma por material. El manejo agronómico proporcionado a las plantas tuvo como guía el descrito por Bastidas et al. (2002).
El experimento se realizó durante 330 días (84 días de previvero y 246 días de vivero). Al final del periodo, se registraron los datos de las siguientes variables biométricas: altura de la planta desde el nivel del suelo del matero hasta el punto de inserción de los foliolos rudimentarios de la hoja cuatro (AP), número de raíces primarias (NRP), diámetro del bulbo (DB), número total de hojas (NTH) y área foliar por planta (AFP). En la hoja cuatro se determinó el largo del raquis (LR), largo del peciolo (LP), área de la sección del peciolo de la hoja (PxS), número de foliolos en la hoja (NFH) y área de la hoja (AH), de acuerdo a las metodologías planteadas por otros autores (Corley et al. 1971; Corley y Tinker 2003; Reyes et al. 2008 y 2009; Taylor et al. 2008).
Adicionalmente, se determinó la acumulación y distribución de biomasa por planta. Para esto, se retiró a cada planta su sistema radical del suelo mediante lavado y, posteriormente, se seccionó toda la planta en sus órganos constitutivos y se secaron en estufa de ventilación forzada, a una temperatura de 75 ºC, hasta lograr peso constante. Las determinaciones de peso fresco y peso seco se realizaron con una balanza analítica. Con los valores por planta de peso seco total de los órganos aéreos (PA) y peso seco de las raíces (SR), se determinaron los valores de la razón parte aérea sistema radicular (PA/SR) (Reyes et al. 2008; Reyes 2012).
Para el análisis estadístico de los datos se utilizó el programa Infostat versión libre (2009) y el programa Excel. El estudio del crecimiento se efectuó a través de análisis de varianza y pruebas de comparación de medias (Tukey, α = 0,05). Para la selección de variables discriminantes se realizaron pruebas de análisis multivariado por componentes principales.
Resultados y discusión
Crecimiento vegetativo
Para la mayoría de variables vegetativas, se presentó efecto de al menos un factor de variación (suelos o genotipos). Solo para las variables diámetro del bulbo, número total de hojas por planta y área de la hoja se presentó efecto de la interacción de los dos factores de variación.
En cuanto al efecto simple de un solo factor de variación, se observó que el suelo presentó efecto para la emisión foliar (EF, p < 0,05) y largo de raquis (LR, p < 0,05). El mayor valor de EF (2,16 hojas/mes) se obtuvo en el suelo arenoso franco y el de LR (56,0 cm) en el franco arcillo limoso (tabla 2). Por su parte, el factor genético presentó solo efecto para el área de la sección del peciolo de la hoja (PxS, p < 0,01), el largo del peciolo de la hoja (LP, p < 0,01) y el área foliar por planta (AFP, p < 0,05). Sobre el particular, se encontró que el material DxP exhibió mayor valor de PxS (1,49 cm2) respecto al material OxG (1,29 cm2); mientras que para el LP y el AFP el comportamiento fue contrario, mayor LP (21,2 cm) y AFP (4,0 m2) para el material OxG, respecto al material DxP (13,5 cm y 3,2 m2, respectivamente) (tabla 2).
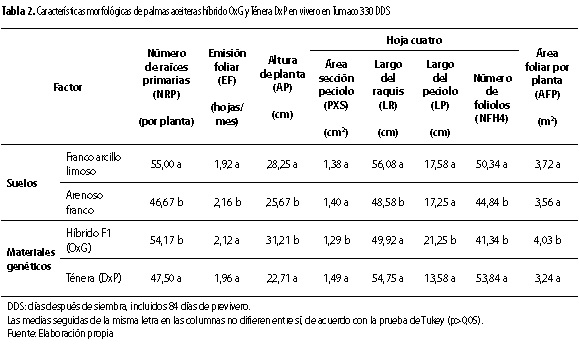
Con relación al efecto simple de los dos factores de variación, se encontró que el suelo y el factor genético, presentaron efecto diferente (p <0,05 y p<0,01, respectivamente) sobre la altura de la planta (AP). Mientras que esos mismos factores presentaron efecto similar (p <0,01) sobre el número de foliolos de la hoja (NFH) y el número de raíces primarias (NRP). Se observó en el suelo franco arcillo limoso la mayor AP (28,2 cm) y para el material OxG (31,2 cm). También para el suelo franco arcillo limoso se observó el mayor NFH (50 foliolos) y para el material DxP (53 foliolos). Mientras que el mayor NRP fue emitido también en el suelo franco arcillo limoso (55 raíces/planta) y por el material OxG (54 raíces/planta) (tabla 2). Para el material OxG, esto se puede explicar por la adaptación a condiciones de exceso de humedad y suelos pesados en ambientes naturales, conocida para su parental femenino Elaeis oleífera (Patiño 1959; Hartley 1988). En general, los valores de las variables vegetativas obtenidos para el material DxP, están dentro de los rangos de valores observados para varios genotipos de ese tipo de material de palma aceitera en otros estudios de crecimiento en vivero realizados en Tumaco (Reyes et al. 2008 y 2009; Reyes 2012).
En lo concerniente a las interacciones de los factores de variación, con relación a la expresión de las variables vegetativas, se encontró que el número total de hojas por planta (NTH), el diámetro del bulbo (DB) y el área de la hoja (AH) variaron (p<0,05) dependiendo de la combinación suelo por genotipo. Destacándose que esto sucedió para las tres variables en el material DxP, porque en el OxG para NTH y DB fue prácticamente invariable (figuras 1a, b y c).
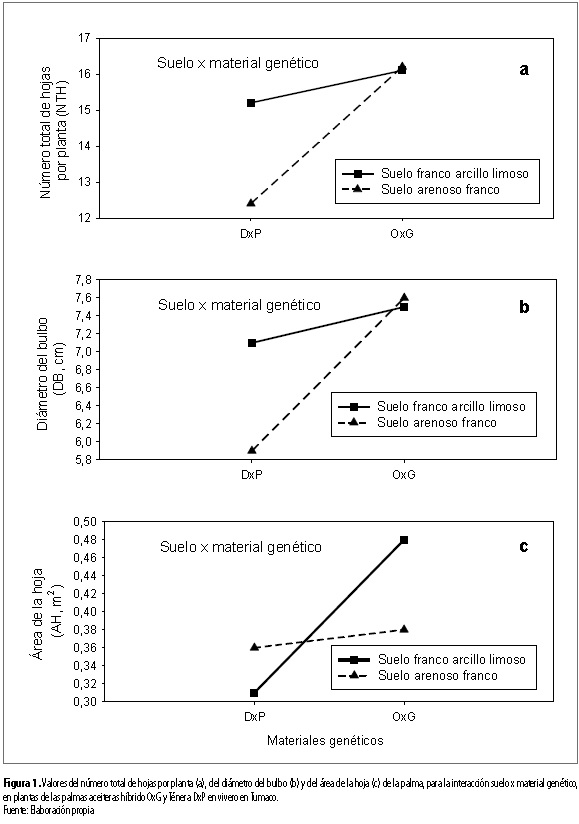
El material DxP presentó mayor NTH (15,2 hojas/planta) y DB (7,1 cm) en el suelo franco arcillo limoso respecto al suelo arenoso franco (13,1 hojas/planta y 5,9 cm, respectivamente) (figuras 1a y b). En cuanto al AH, esta fue mayor para el material OxG cultivado en suelo franco arcillo limoso (0,48 m2), respecto a cuándo fue cultivado en suelo arenoso franco (0,38 m2) (figura 1c). Por su parte, para el material DxP, el AH fue mayor en el suelo arenoso franco (0,36 m2) con relación al suelo franco arcillo limoso (0,31 m2) (figura 1c). Con esto se evidenció que el área de la hoja (AH) de esos materiales puede aumentar o disminuir dependiendo del tipo de suelo utilizado en el vivero. Lo que sugiere que los materiales evaluados adaptan su conformación morfológica de manera diferencial a modificaciones ambientales en el suelo. Esto es concordante con lo informado por Benincasa (1988) y Bulgarelli et al. (1993), quienes indican que el comportamiento del crecimiento de las plantas puede ser diferente, dependiendo de las condiciones ambientales que se le suministren, y que estas diferencias se pueden presentar aún entre materiales con orígenes genéticos similares.
En síntesis, se observó que el material OxG, independiente del tipo de suelo en el cual se cultivó, presentó mayores valores de NTH, DB y AH en comparación con el material DxP (figuras 1a, b y c).
En general, el material OxG se caracterizó por presentar mayores valores que el material DxP, para la mayoría de variables vegetativas evaluadas (tabla 2). De igual forma, se pudo constatar que el material OxG mostró mayor crecimiento en el suelo tipo franco arcillo limoso, para las variables vegetativas que presentaron efecto de la combinación suelos por genotipo (figuras 1a, b y c), suelo de tipo pesado, con alta capacidad de retención de humedad y con baja aereación (tabla 1). Comportamiento de crecimiento explicable, por la adaptación de su parental femenino Elaeis oleífera a condiciones de suelos pesados en ambientes naturales (Patiño 1959; Hartley 1988).
En cuanto al material DxP, el trabajo desarrollado por Reyes et al. (2009), demostró que, en las condiciones ambientales de Tumaco, los suelos del tipo arenoso franco son los más adecuados para viveros de ese tipo de palmas, con relación a los suelos del tipo franco arcillo limoso, lo que coincidió con el comportamiento vegetativo observado para ese material en este estudio.
Los resultados anteriores indican un comportamiento diferencial y contrastante, del crecimiento vegetativo en vivero, para las palmas de los materiales OxG y DxP evaluados, respecto al tipo de suelo en que crecen.
En general, el material OxG se caracterizó por presentar mayores valores que el material DxP, para la mayoría de variables vegetativas evaluadas (tabla 2). De igual forma, se pudo constatar que el material OxG mostró mayor crecimiento en el suelo tipo franco arcillo limoso, para las variables vegetativas que presentaron efecto de la combinación suelos por genotipo (figuras 1a, b y c), suelo de tipo pesado, con alta capacidad de retención de humedad y con baja aereación (tabla 1). Comportamiento de crecimiento explicable, por la adaptación de su parental femenino Elaeis oleífera a condiciones de suelos pesados en ambientes naturales (Patiño 1959; Hartley 1988).
En cuanto al material DxP, el trabajo desarrollado por Reyes et al. (2009), demostró que, en las condiciones ambientales de Tumaco, los suelos del tipo arenoso franco son los más adecuados para viveros de ese tipo de palmas, con relación a los suelos del tipo franco arcillo limoso, lo que coincidió con el comportamiento vegetativo observado para ese material en este estudio.
Los resultados anteriores indican un comportamiento diferencial y contrastante, del crecimiento vegetativo en vivero, para las palmas de los materiales OxG y DxP evaluados, respecto al tipo de suelo en que crecen.
Acumulación y distribución de biomasa
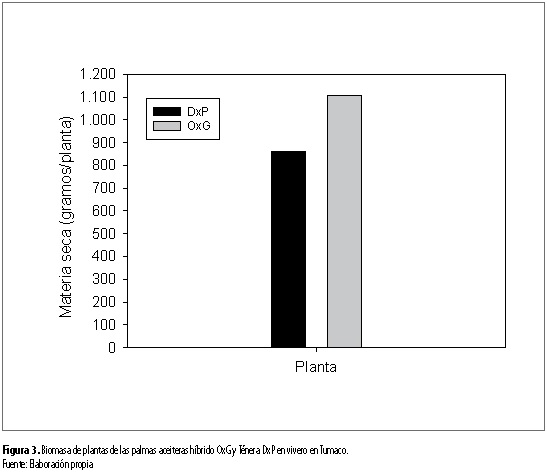
La acumulación de biomasa por órgano y para toda la planta no dependió de la interacción entre los factores suelo y genotipo. Dependió del factor genético, para la mayoría de los órganos constituyentes de la planta y la planta como un todo. Efecto simple que se destacó para la biomasa de raíces (p<0,05), bulbo (p<0,05), foliolos (p<0,05), hojas flecha (p<0,01), parte área (p<0,01) y total planta (p<0,01). El material OxG presentó los mayores valores de biomasa para las anteriores variables con relación al material DxP (tabla 3 y figura 2). Dicha superioridad se destacó para las raíces (239,3 g/planta), el bulbo radical (287,5 g/planta) y los foliolos (303,3 g/ planta) con relación al DxP (171,7 g/planta, 191,7 g/ planta y 225 g/planta, respectivamente) (tabla 3 y figura 2). Esos órganos de las palmas son importantes para el anclaje y absorción de agua y nutrientes de la solución del suelo, así como para la actividad fotosintética de las palmas. Estas características pueden favorecer la actividad fisiológica de los materiales OxG con relación a los DxP, creciendo en condiciones ambientales limitantes. En este sentido, se pudo observar cómo los mayores valores de biomasa total por planta se observaron para el material OxG (1.106,9 gramos) respecto al DxP (860,3 gramos) (tabla 3 y figura 3).
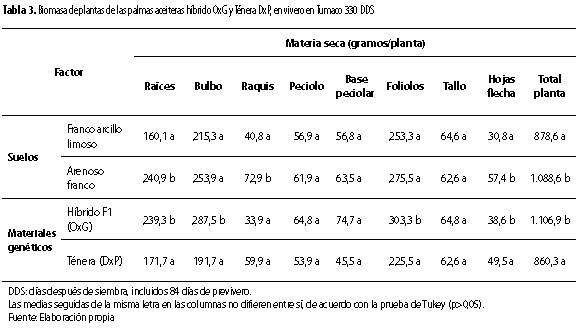
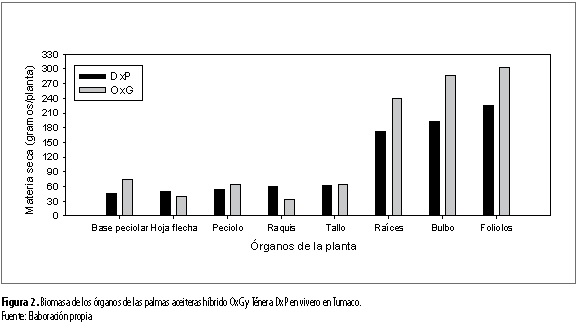
Por otra parte, se determinó que para la biomasa de raíces (p <0,01), biomasa del raquis (p<0,01), biomasa de hojas flecha (p<0,05) y biomasa total de la planta (p<0,05) también se presentó efecto del suelo, observándose los mayores valores de biomasa para esos órganos y el total de la planta, en el suelo arenoso franco (tabla 3). Lo que sugiere, que condiciones ambientales de mejor aereación del suelo favorecen el desarrollo y el crecimiento de órganos como las raíces y la planta como un todo, en las palmas aceiteras.
En cuanto al comportamiento de las variables vegetativas, estos resultados sobre la acumulación y distribución de biomasa, ratifican un mayor crecimiento y desarrollo en vivero del material OxG sobre el DxP.
Asimismo, la razón parte aérea sistema radical (PA/SR) de las palmas fue afectada por los factores suelo (p<0,01) y genético (p<0,05), pero no por su combinación. Con relación a los valores de PA/SR observados (3,5 a 4,4), están dentro del rango de valores observados en Tumaco para cultivares del genotipo DxP (Reyes et al. 2008).
En cuanto a suelo, la razón PA/SR presentó el menor valor (3,5) en el suelo arenoso franco (tabla 4). Esto indica que ese suelo favoreció una mayor acumulación de biomasa en el sistema radical de las palmas con relación al franco arcillo limoso. Resultado corroborado con lo observado en este estudio, relacionado con la mayor acumulación de biomasa de raíces por planta en el suelo arenoso franco (tabla 3). Este comportamiento puede estar relacionado con la mayor capacidad de aereación y, por consiguiente, mayor disponibilidad de oxígeno, que puede ofrecer el suelo arenoso franco a similar contenido de humedad que el suelo franco arcillo limoso, porque se conoce que la deficiencia de oxígeno perjudica el proceso de respiración de la raíz y, por lo tanto, los procesos metabólicos de la planta y su crecimiento (Taiz y Zeiger 1991).
En cuanto al factor genético, el menor valor para la razón PA/SR lo presentó el material OxG (3,6) (tabla 4), lo cual indica, que ese material presentó mayor capacidad de acumulación de biomasa en el sistema radical de las palmas con relación al material DxP. Esto sugiere que el material OxG presenta el atributo de una menor razón PA/SR con relación al DxP, resultado relacionado con una mayor acumulación de biomasa de raíces y en el bulbo por planta para el material OxG (tabla 3). Asimismo, es favorecido por la mayor emisión de raíces primarias por parte del material OxG (tabla 2), porque, en los materiales estudiados, las raíces primarias se emiten del bulbo radical y, a partir de ellas, se genera el sistema radical de las palmas que es fasciculado, conformado por raíces primarias, secundarias, terciarias y cuaternarias. Por su parte, para el material DxP, Reyes et al. (2008) no encontraron en Tumaco diferencias entre cultivares sembrados en suelo arenoso, para la razón PA/SR, cuyo rango de valores (3,4 a 4,9) incluye los valores observados en este estudio (tabla 4).
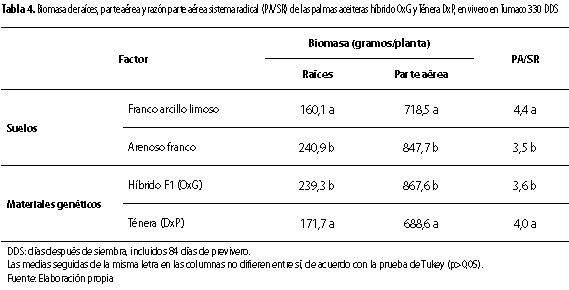
En general, la discusión sobre lo encontrado para el comportamiento de la razón PA/SR en este estudio es soportado por varios autores, quienes manifiestan que esa razón está determinada genéticamente, pero puede ser modificada por el ambiente (Marschner 1986; Rodríguez y Leihner 2006). Por otra parte, el material OxG mostró una característica diferencial respecto al DxP, de poder destinar mayor proporción de sus asimilados para formar sistema radical en la planta, característica muy importante para la actividad de absorción de agua y nutrientes.
La distribución porcentual de biomasa entre los órganos de la planta solo fue afectada por el factor suelo (p <0,01) en lo relacionado con las raíces, correspondiendo el mayor valor (24,53 %) al suelo arenoso franco (tabla 5). Esto es confirmado por lo encontrado en este estudio, para la acumulación de biomasa en ese órgano de la planta en el suelo arenoso franco (tabla 3). Lo que sugiere, que las propiedades físicas de un suelo influyen en la distribución de asimilados de las palmas en vivero y, principalmente, en los destinados para las raíces, determinación que concuerda con Reyes et al. (2009), en un estudio sobre el crecimiento en vivero de la palma DxP realizado en Tumaco.
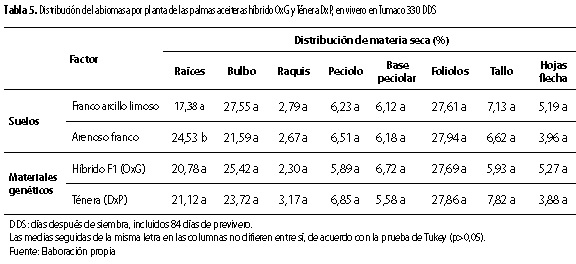
En cuanto a la preferencia u orden de destino, para la distribución de asimilados o biomasa entre sus órganos, por parte de las palmas de los materiales evaluados (tabla 5 y figura 4), se encontró que: Material DxP: foliolos > bulbo > raíces > tallo > peciolo >base peciolar > hojas flecha > raquis Material OxG: foliolos > bulbo > raíces > base peciolar > tallo > peciolo > hojas flecha > raquis
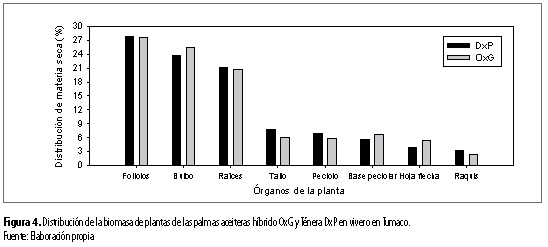
Con relación al material DxP, el orden de preferencia de destino de asimilados observado concuerda, en gran parte, con el determinado en vivero en Tumaco por Reyes et al. (2008).
Para el material OxG, se encontró, respecto al DxP, modificaciones para el orden de preferencia de destino de asimilados para el órgano base peciolar, con relación a los órganos tallo y peciolo (tabla 5 y figura 4). Resultado que indica la existencia de modificaciones para la preferencia de distribución de asimilados según el material de palma aceitera sembrado, para los órganos base peciolar, tallo y peciolo. Condición que puede estar relacionada con la fuerza de sumidero intrínseca o atributo de cada uno de los componentes de la planta en cada material genético, para organizar la biomasa total y la arquitectura de la planta (Villalobos et al. 2001; Wareing y Patrick 1975; Reyes et al. 2008).
En cuanto al material DxP, los valores observados para biomasa de órganos, biomasa total y razón parte aérea sistema radical de las palmas (tablas 3 y 4) están dentro de los rangos de valores observados en estudios de vivero realizados en diferentes suelos en Tumaco (Reyes et al. 2008 y 2009).
Cabe resaltar que, a pesar de las diferencias observadas entre materiales para el orden de distribución de sus asimilados entre los componentes de las palmas, los dos tipos de materiales genéticos estudiados presentaron crecimiento armónico de sus plantas, en cada tipo de suelo en que fueron plantados y cultivados.
En general, en este estudio, el material OxG se caracterizó por presentar mayores valores que el DxP para la mayoría de variables vegetativas evaluadas. Por su parte, en cuanto al comportamiento de estas últimas, la acumulación y distribución de biomasa de los dos materiales ratificaron el mayor crecimiento y desarrollo en vivero del material OxG sobre el DxP. Resultado que está relacionado con el mayor vigor de las plantas de OxG (E. oleífera x E. guineensis) respecto a las de DxP (E. guineensis x E. guineensis), por ser el OxG un híbrido interespecífico (E. oleífera x E. guineensis).
El crecimiento y el desarrollo diferencial y contrastante en vivero, respecto a los dos tipos de suelo donde crecieron las plantas de los materiales OxG y DxP, sugiere la existencia de atributos fisiológicos propios en esos materiales, que les permiten adaptarse de manera diferencial a modificaciones ambientales en el suelo. Características y adaptación que probablemente se manifiestan también en el crecimiento y desarrollo de sus palmas en campo y condiciones de cultivo, como ya se ha observado en Tumaco, Colombia (Bastidas et al. 2007 y 2010). Al respecto, ya es conocido que la aereación del suelo afecta el crecimiento y la sanidad de las plantas y de la palma aceitera (Bulgarelli et al. 1993; Forsythe 1997). Esto es debido a que la deficiencia de oxígeno perjudica el proceso de respiración de la raíz, lo que causa una carencia de energía (ATP) para los procesos metabólicos básicos de la planta y su crecimiento (Taiz y Zeiger 1991). Además, ya se han determinado valores críticos de espacio aéreo (EA) en el suelo para varios cultivos, respecto a sus curvas de rendimiento, los cuales se encuentran entre 11 % y 25 % de EA (Forsythe 1997). Por su parte, en cuanto a los suelos empleados en esta investigación, es de recordar que el arenoso franco presenta, con relación al franco arcillo limoso, condiciones de mejor espacio aéreo y aereación, menor retención de humedad y mayor drenaje, que le permite poseer mayor disponibilidad de oxígeno a un contenido de humedad similar de los dos suelos.
Por otra parte, todo lo anterior ratifica la necesidad de adelantar trabajos de investigación enfocados a comprender el comportamiento del crecimiento y el desarrollo de genotipos de estos materiales, creciendo en condiciones ambientales específicas propias de cada zona en donde se van a cultivar, conocimientos básicos indispensables para el desarrollo de sistemas de producción sostenibles y competitivos.
Identificación y selección de variables morfológicas discriminantes
El resultado del análisis multivariado, aplicado a los datos de características vegetativas, mostró que con solo dos componentes principales se puede explicar el 91 % de la variabilidad de dichos datos de crecimiento. Se determinó que el área de la sección del peciolo (PxS), el largo del raquis de la hoja cuatro (LR) y la emisión foliar (EF) fueron las variables que presentaron más sensibilidad discriminante para diferenciar el comportamiento del crecimiento y el desarrollo de las plantas en vivero de materiales OxG y DxP, esto en función del tipo de suelo donde crecieron. Variables cuyo registro no requiere la utilización de métodos destructivos y son de fácil medición en vivero y campo (Bulgarelli et al. 1993; Reyes et al. 2008). Además, las variables PxS y EF ya han sido identificadas por otros autores, utilizando otras metodologías, como discriminantes para diferenciar el comportamiento del crecimiento y el desarrollo de palmas DxP (Bulgarelli et al. 1993; Corley y Tinker 2003; Reyes et al. 2008 y 2009). Por lo tanto, en futuros estudios con materiales OxG y DxP similares al presente, son esas las variables vegetativas indicadas a registrar en las condiciones ambientales de Tumaco. Este resultado es coherente con el comportamiento del crecimiento y desarrollo de los dos tipos de materiales de palma aceitera evaluados en este estudio. Lo que corrobora la aplicabilidad en estudios de palmas aceiteras de la metodología estadística de análisis multivariado por componentes principales, utilizada en este trabajo para la identificación y la selección de variables discriminantes, ya informada y empleada en palma aceitera DxP por otros autores (Martínez et al. 1999; Brito et al. 2006; Reyes 2012).
Conclusiones
En la etapa de vivero, en condiciones de suelos con alta retención de humedad y baja aereación, las plantas del híbrido OxG (Elaeis oleífera x Elaeis guineensis) presentan mejor crecimiento y desarrollo que las plantas del material Ténera DxP (Elaeis guineensis x Elaeis guineensis).
Las plantas del material OxG acumulan más biomasa en la mayoría de sus órganos constituyentes que las plantas del material DxP, destacándose dicha superioridad para las raíces y el bulbo radical, órganos importantes para el anclaje y absorción de agua y nutrientes de la solución del suelo, así como para los foliolos, órganos importantes para la actividad fotosintética de las palmas.
Los materiales OxG y DxP poseen atributos propios para organizar la biomasa total y la arquitectura de sus plantas, indicado por sus preferencias del destino, la distribución y la acumulación de asimilados para los órganos base peciolar, tallo y peciolo.
Para evaluar el comportamiento del crecimiento de las palmas aceiteras OxG y DxP, en la etapa de vivero, en función del tipo de suelo, se pueden utilizar tres características morfológicas: área sección del peciolo (PxS), largo del raquis (LR) y emisión foliar (EF), para cuya determinación no se requiere el empleo de metodologías destructivas convencionales.
Los resultados obtenidos en este trabajo manifiestan la necesidad de adelantar investigaciones sobre el comportamiento del crecimiento y el desarrollo de genotipos de materiales OxG y DxP en las condiciones ambientales de cada zona de cultivo, conocimientos indispensables para el desarrollo de sistemas de producción sostenibles y competitivos.
Agradecimientos
Los autores agradecen a Corpoica y Colciencias por el aporte financiero para realizar esta investigación dentro de la ejecución de la beca-pasantía Corpoica-Colciencias, asimismo, Lizardo Norbey Ibarra Ruales al Programa Jóvenes Investigadores e Innovadores de Colciencias con tutoría de Rafael Reyes Cuesta.
Referencias
Bastidas S, Peña E, Reyes R, Medina C, Villa A, Tolosa W. 2010. Pruebas agronómicas para certificar la tolerancia de los híbridos OxG a pudrición del cogollo. Fase I. Ministerio de Agricultura y Desarrollo Rural; [consultado 2011 jun]. http://www.agronet.gov.co/BibliotecaDigital.html. [ Links ]
Bastidas S, Peña E, Reyes R, Pérez J, Tolosa W. 2007. Comportamiento agronómico del cultivar híbrido RC1 de palma de aceite (Elaeis oleífera x Elaeis guineensis) x Elaeis guineensis. Corpoica Cienc Tecnol Agropecu. 8(1):5-11. [ Links ]
Bastidas S, Peña E, Reyes R, Casas H. 2002. Recomendaciones generales para el manejo de semilla germinada y viveros de palma de aceite (Elaeis guineensis Jacq.). Boletín divulgativo N.o 12. Tumaco: Corpoica. [ Links ]
Bastidas S, Reyes R, Peña EA. 2006. Palma de aceite Ténera Corpoica El Mira: alternativa nacional. Revista Innovación y Cambio Tecnológico. 4(2-3):47-50. [ Links ]
Benincasa MMP. 1988. Analise de crescimento de plantas (Noções básicas). Jaboticabal: Funep. [ Links ]
Breure K. 2003. The search for yield in oil palm. Basic principles. En: Fairhust TH, Hardter R, editores. Oil palm. Management for large and sustainable yields. Sigapure: Potash & Phosphate Institute/Potash & Phosphate Institute of Canada (PPI/PPIC) and International Potash Institute (IPI). pp. 59-98. [ Links ]
Brito OL, Méndez JR, Barrios R. 2006. Caracterización de variables de crecimiento de 17 progenies de palma aceitera (Elaeis guineensis Jacq.) en el estado Monagas, Venezuela. UDO Ag. 6(1):33-40. [ Links ]
Bulgarelli J, Chinchilla C, Richardson D L, Salas A. 1993. Crecimiento en vivero de los materiales Deli x Avros, Deli x Calabar y Deli x Ekona. Curvas de referencia. San José: ASD de Costa Rica. [ Links ]
Chee KH, Chiu SB, Chan SM. 1998. Plántulas de pre-vivero cultivadas en bandejas de macetas. Revista Palmas. 19(4):51-54. [ Links ]
Contreras AP. 1996. Estimación y cálculo de parámetros de crecimiento en palma africana (Elaeis guineensis Jacq.) Nolí (E. oleífera) y el híbrido (E. guineensis x E. oleífera) [trabajo de grado]. [Bogotá]: Universidad Nacional de Colombia. [ Links ]
Corley RHV, Hardon JJ, Tang GY. 1971. Analysis of growth of the oil palm (Elaeis guineensis Jacq.) I. Estimation of growth parameters and application in breeding. Euphytica. 20(2):307-315. [ Links ]
Corley RHV, Tinker PBH. 2003. The oil palm. 4.a edición. Oxford: Blackwell Science. [ Links ]
Corredor A, Martínez G, Silva A. 2008. Problemática de la pudrición del cogollo en Tumaco e instrumentos para su manejo y la renovación del cultivo. Revista Palmas. 29(3):11-16. [ Links ]
Coto E, Chinchilla C, Bulgarelli J, Palma T. 2002. Vegetative growth in prenursery of five commercial varieties of oil palm (Elaeis guineensis Jacq.). ASD Oil Palm Papers. 23:1-13. [ Links ]
Forsythe W. 1997. Las condiciones físicas, la producción agrícola y la calidad del suelo. Agron Costarric. 21(1):35-47. [ Links ]
Hartley CWS. 1988. The oil palm (Elaeis guineensis Jacq.). 3.a edición. Londres: Longman. [ Links ]
Hunt R. 2003. Growth analysis, individual plants. En: Thomas B, Murphy DJ, Murray D, editors. Encyclopaedia of applied plant sciences. Londres: Academic Press; [consultado 2010 feb]. pp. 579-588. http://people.exeter.ac.uk/rh203/EAPS_article.pdf. [ Links ]
Marschner H. 1986. Mineral nutrition of higher plants. London: Academic Press. [ Links ]
Martínez R, Ochoa IE, Bastidas S. 1999. El mejoramiento de la palma de aceite (Elaeis guineensis Jacq.) en Colombia. Metodología estadística. Revista Palmas. 20(2):9-21. [ Links ]
Meunier J. 1991. Una posible solución genética para el control de pudrición de cogollo en la palma aceitera. Híbrido interespecifico E. oleífera x E. guineensis. Revista Palmas. 12(2):39-42. [ Links ]
Patiño VM. 1977. El corozo o nolí (Elaeis oleifera (HBK) Cortés ex Wessels Boer) recurso natural oleaginoso de Colombia. Cespedesia. 6(21-22):7-122. [ Links ]
Preciado CA, Bastidas S, Betancourth C, Peña E, Reyes R. 2011. Predicción y control de la cosecha en el híbrido interespecífico Elaeis oleífera x Elaeis guineensis en la zona palmera occidental de Colombia. I. Determinación del periodo de madurez para obtener racimos con alto contenido de aceite. Corpoica Cienc Tecnol Agropecu. 12(1):5-12. [ Links ]
Reyes R. 2012. Respuesta fisiológica de la palma aceitera (Elaeis guineensis Jacq.) a la disponibilidad de agua en el suelo [tesis de doctorado]. [San José de Costa Rica]: Universidad de Costa Rica. [ Links ]
Reyes R, Rodríguez N, Peña E, Bastidas S. 2008. Crecimiento en vivero de materiales comerciales de palma aceitera (Elaeis guineensis Jacq.) en Tumaco, Colombia. Corpoica Cienc Tecnol Agropecu. 9(2):12-18. [ Links ]
Reyes R, Rodríguez N, Peña E, Bastidas S, Ibarra N. 2009. Evaluación del crecimiento de cuatro genotipos de palma aceitera (Elaeis guineensis Jacq.) en dos tipos de suelo de diferentes características físicas. I. Vivero. Ponencia presentada en: II Seminario Experiencias en la Inversión en Ciencia y Tecnología para el Sector Agropecuario Colombiano. Bogotá, Colombia. [ Links ]
Rodríguez W, Leihner D. 2006. Análisis de crecimiento vegetal. Fisiología de la producción de cultivos tropicales. San José: Editorial Universidad de Costa Rica. [ Links ]
Salisbury FB, Ross C W. 2000. Fisiología de las plantas 3. Desarrollo de las plantas y fisiología ambiental. Madrid: Thompson Ediciones. [ Links ]
Taiz L, Zeiger E. 1991. Plant physiology. San Francisco: Benjamin Cummings. [ Links ]
Taylor M, Chinchilla C, Rodríguez W. 2008. Leaf architecture and estimates of leaf area and leaf dry weight in young compact oil palm clones (Elaeis guineensis Jacq.). ASD Oil Palm Papers. 32:27-47. [ Links ]
Umaña CH, Chinchilla C, Richardson DL. 1990. Material de desarrollo avanzado en viveros de palma aceitera (Elaeis guineensis Jacq.). II. Condiciones del sustrato. Turrialba. 40(4):440-451. [ Links ]
Villalobos E, Marín W, Berninger F. 2001. Transporte y partición del carbón. En: Villalobos E, editor. Fisiología de la producción de cultivos tropicales. Procesos fisiológicos básicos. Fascículo I. San José: Editorial Universidad de Costa Rica. pp. 83-118. [ Links ]
Wareing PF, Patrick J. 1975. Source-sink relations and the partition of assimilates in the plant. En: Cooper JP, editor. Photosynthesis and productivity in different environments. London: Cambridge University Press. pp.431-499. [ Links ]
Zambrano JE. 2004. Los híbridos interespecíficos Elaeis oleífera H.B.K x Elaeis guineensis Jacq. Una alternativa de renovación para la zona oriental de Colombia. Revista Palmas. 25(2):339-349. [ Links ]
