










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
versão impressa ISSN 0122-8706
Corpoica cienc. tecnol. agropecu. vol.19 no.2 Mosquera maio/ago. 2018
https://doi.org/10.21930/rcta.vol19_num2_art:520
Plant breeding
Resistance evaluation of a Fl hybrid of Solanum quitoense Lam. to Neoleucinodes elegantalis (Guenée) and Meloidogyne incognita (Kofoid & White)
1 Occasional Lecturer, Universidad Nacional Abierta y a Distancia (UNAD), Escuela de Ciencias Agrícolas, Pecuarias y del Medio Ambiente (ECAPMA). Dosquebradas, Colombia. Email: manuel.polanco@unad.edu.co. orcid.org/0000-0002-4810-0081.
2 Occasional Lecturer, Universidad Nacional Abierta y a Distancia (UNAD), Escuela de Ciencias Agrícolas, Pecuariasy del Medio Ambiente (ECAPMA). Dosquebradas, Colombia. Email: susana.gomez@unad.edu.co.
3 Occasional Lecturer, Universidad Nacional Abierta y a Distancia (UNAD), Escuela de Ciencias Agrícolas, Pecuarias y del Medio Ambiente (ECAPMA). Dosquebradas, Colombia. Email: juan.padilla@unad.edu.co.
The aim of this study was to cross lulo plants of cv. La Selva to obtain a hybrid with tolerance to the fruit borer Neoleucinodes elegantalis Guenée (Lepidoptera: Crambidae) and to the nematode Meloidogyne incognita (Kofoid & White) (Nematoda: Heteroderidae), to obtain better-adapted plants to the conditions of the Colombian coffee-growing region and with bigger and non-dehiscent fruits. La Selva cultivar is a hybrid developed from the interspecific backcross of Solanum quitoense Lam. χ Solanum hirtum Vahl with plants of lulo cv. Castilla Larga Vida. The experiment was conducted in the municipality of Dosquebradas, Risaralda, Colombia, located at 1,465 m a.s.l. Fi plants were obtained from reciprocal crossings. When inoculated with the nematode M. incognita, plants showed susceptibility in their seedling stage; however, when we carried out the evaluation six months after transplantation under field conditions, nematode infestation was less than 1%, which likely indicates the tolerance of these materials to the nematode. When assessing the resistance of the hybrid to the attack of N. elegantalis, we found that the evaluated materials were resistant to this insect. The resulting hybrids showed good agronomic characteristics, such as a good morphological structure and vigor, high productivity, good solar exposure adaptation, large fruits (5.6 cm average diameter) similar to those of cv. Castilla, with yellow peel, green pulp and non-dehiscent fruits with pleasant aroma and flavor. As an undesirable characteristic plants had thorns on leaves and stems.
Keywords: diversity; fruit borer; hybrids; plant nematodes; plant breeding; Solanum quitoense
En este estudio se cruzaron plantas de lulo del cultivar La Selva, híbrido del retrocruzamiento interespecífico de Solanum quitoense Lam. χ Solanum hirtum Vahl (Solanaceae), con plantas del cultivar Castilla Larga Vida, con el fin de obtener un material de frutos grandes, no dehiscente, adaptado a la zona cafetera colombiana y con resistencia al pasador del fruto Neoleucinodes elegantalis Guenée (Lepidoptera: Crambidae) y al nematodo Meloidogyne incognita (Kofoid & White) (Nematoda: Heteroderidae). El experimento se realizó en el municipio de Dosquebradas, Risaralda (Colombia), a 1.465 m s. n. m. Al ser inoculadas con el nematodo M. incognita, las plantas F1, obtenidas de los cruzamientos recíprocos, mostraron susceptibilidad en la etapa de almácigo.
Sin embargo, al realizar la evaluación seis meses después del trasplante en condiciones de campo, se encontró menos del 1% de infestación del nematodo, lo que hace presumir la tolerancia de los híbridos a este organismo. De igual forma, se evidenció la resistencia de los materiales al ataque de N. elegantalis. Los híbridos resultantes presentaron buenas caracte rísticas agronómicas, como vigor y estructura buenos, alta productividad, buena adaptación a condiciones de libre exposición solar, frutos grandes (similares a los del lulo cv. Castilla), corteza amarilla y pulpa verde, sin rajamiento, y de aroma y sabor agradables. Las plantas presentaron una característica indeseable, que consistió en espinas en tallos y hojas.
Palabras clave: diversidad; fitomejoramiento; híbridos; nematodos de plantas; pasador del fruto; Solanum quitoense
Introduction
Lulo, Solanum quitoense Lam. (Solanacea) Castilla cultivar, is one of the Andean fruits with a huge commercial potential, given its wide acceptance in national markets, and because it is also considered a promissory product for international markets. However, the demand has been reduced due to the current production system used, characterized by a high consumption of pesticides, which causes the detection of traces of active ingredients not allowed in international markets (Muñoz, 2011).
During 2014, 33,522 t of lulo were produced in Colombia, with an average yield of 9.6 t/ha per year. Antioquia is the main producer department with 10,477 t, corresponding to 31.25% of the total production, followed by the departments of Huila, Cundinamarca and Santander.
In the period from January to July 2014, the main destinations of this fruit were France, contributing 49 % of the demand, with a value of US$38,990.50, followed by Panama (14 %), Spain (11 %) and Aruba (4 %) (National Administrative Department of Statistics [DANE], 2015).
Some authors have recognized two botanical varieties: Solanum quitoense var. septentrionale, which has thorns, and Solanum quitoense var. quitoense, which is thornless (Fory et al., 2010; Morton, 1987; Schultes, & Cuatrecasas, 1953; Whalen, Costich, & Heiser, 1981), but there is still controversy regarding this aspect.
Lulo cv. La Selva is the product of two backcrosses in search of a spineless population of the hybrid obtained by Professor C. Heiser (Indiana University, usa) of Solanum quitoense χ Solanum hirtum (locally known as "lulo de perro") (Bernal, Lobo, & Londoño, 1998), and is characterized for not having thorns neither in its leaves nor in the stem. It is a precocious cultivar, which begins its production between six and seven months after planting.
Its production can reach up to 20 t/ha and it is destined for the agribusiness because it has a very short postharvest life, since its pulp oxidizes quickly and has high dehiscence; this causes the fruit to crack even before reaching its full maturity. However, it has tolerance to nematodes and to N. elegantalis (Medina, Lobo, Martínez, & Riaños, 2004).
Lulo cv. Castilla and cv. Castilla Larga Vida have thorns in stems and leaves, they start producing between seven and nine months after transplantation and reach a production of 8 to 10 t/ha, destined mostly for the fresh market. Among the pests that affect severely the crop are Neoleucinodes elegantalis (Guenée) (Lepidoptera: Crambidae) which is the main pest, and the nematode Meloidogyne incognita (Kofoid & White) (Nematoda: Secernentea: Tylenchida: Heteroderidae) (Bernal & Díaz, 2006).
Neoleucinodes elegantalis is a moth that in its larval state drills and bores lulo fruits, feeding on these until it reaches adulthood. Larvae enters the fruit from the third instar and stays there until the fifth instar, and then drops to the ground where it pupates. However, when leaving, the larvae leaves an exit hole that allows the entry of other insects and microorganisms, damaging completely the fruit (Instituto Colombiano Agropecuario [ICA], 2011, Marcano, 1991, Obando, 2011, Parra, Barona, & Vallejo, 1994).
Due to their particular habits in which the larvae live inside the fruit, chemical control is ineffective (Díaz-Montilla, 2013). Moreover, it is considered a quarantine pest in Chile, the US, and Peru, which limits fruit exports (Obando, 2011).
Regarding the nematode M. incognita, according to Tamayo (2001) , it is a pest that attacks lulo in all producing areas of Colombia. The attack starts in the nursery, producing galls and nodes in roots, which prevents the absorption of water and nutrients; this weakens plants that show symptoms such as smaller leaves, chlorosis of lower leaves and temporary withering on sunny days. Moreover, it can reduce production by more than 50 % (Franco et al., 2002).
Control based on agrochemicals is the most used by farmers but has huge disadvantages. As products used are extremely toxic and residual, this pollutes the environment and, when applied repeatedly, they cause resistance from the organism to the product, aggravating further the problem (Andrés, 2002).
Therefore, the best control method for these two crop-limiting pests is the use of resistant cultivars and varieties. Lobo and Medina (2000) have reported that lulo cv. La Selva is resistant to nematodes and to N. elegantalis. The definition of varietal resistance to pests is variable, but, in practical agricultural terms, we can say that a resistant cultivar is one that yields more than a susceptible one when facing the invasion of a pest (Maxwell & Jennings, 1980). Therefore, the resistance of plants is relative, and is based on the comparison with those that lack that quality, that is, the susceptible ones. According to Panda and Khush (1995) , varietal resistance has the following fundamental characteristics: a) is heritable and controlled by one or more genes; b) is relative and can only be measured when compared to a susceptible cultivar of the same species; c) is qualitatively or quantitatively measurable; and d) is variable and can be modified by biotic and abiotic factors.
According to the aforementioned, the aim of this study was to evaluate the resistance or varietal tolerance of the Fi materials obtained from the reciprocal crossing between lulo cv. Castilla, natural selection Larga Vida and cv. La Selva to the nematode M. incognita and to the moth N. elegantalis.
Materials and methods
Collection of parental material
Plant material of lulo cv. La Selva was collected in the town of Caucayá, municipality of Belén de Umbría, Risaralda (Colombia), with the following geographic coordinates: 05°13' N and 75°48' W, at an altitude of 1,320 m a.s.l., with an average temperature of 26 °C and an annual rainfall of 2,150 mm.
Cuttings of basal suckers from selected plants were obtained due to their excellent production and good phytosanitary condition. Cuttings were planted in plastic bags with a substrate composed of three parts of soil plus one of organic matter. Once planted, a drench application with Trichoderma spp. was made to prevent the attack of the damping-off complex (Fusarium oxysporum, Phytophthora infestans, Sclerotinia sclerotiorum) (ICA, 2011).
For the case of natural selection of lulo cv. Castilla Larga Vida, fruits were harvested from the best plants of a commercial cultivation that were isolated from any other cultivar in the town of Andica de Belén de Umbría, Risaralda. The location has an altitude of 1,370 m a.sl, an average temperature of 25 °C, an average annual precipitation of 2,030 mm, and have the following geographic coordinates: 05°14' N and 75°52' W.
Seeds were extracted from fruits weighing more than 150 g and a diameter greater than 8 centimeters, mixed as a mass selection and planted in a germinator. Twenty-eight days after planting, seedlings that emerged were registered, achieving a germination of 70.3 %. When seedlings emitted the third pair of leaves, they were transplanted to plastic bags, where they remained for four months until they had three true leaves.
Parent plant management
Seedlings from both parents were planted in a plot of Universidad Nacional Abierta y a Distancia (UNAD) in the municipality of Dosquebradas, Risaralda, located at an altitude of 1,460 m a.s.l., with an average temperature of 21 °C, an average annual rainfall of 2,200 mm, and an average relative humidity of 85 %.
Plants were sown in holes of 30 χ 30 χ 30 cm in a square arrangement with distances of 2 m between plants and 3 m between rows. We added 1.5 kg of composted organic matter to each hole as well as 150 g of commercial mycorrhizae to favor a better phosphorus absorption.
All the traditional cultural management practices obtained due to their excellent production and such as weed control, soil fertilization with NPK and secondary elements, as well as foliar fertilization with minor elements were carried out. Shape pruning was also done to eliminate shoots or suckers from the stem that were below 30 cm in height.
Crossings
Four months after transplantation flowering occurred. At that time, flowers with long pistils were selected in both materials (lulo cv. La Selva and lulo cv. Castilla Larga Vida), and reciprocal crosses were made, that is, the two materials were used as parent, i.e. as mother and as father plants. Flowers were emasculated removing the stamens, and the chosen flowers were pollinated as females. In figure 1, we can see the fruits resulting of these crosses during their development process.

Figure 1 Fruits in development obtained from the crosses of lulo cv. Castilla Larga Vida × cv. La Selva.
From these crosses, fruits were obtained from which the Fi seed was extracted (figure 2). The seeds of all the F1 fruits of each reciprocal cross were kept separate as a mass selection. Three hundred and forty-four seeds of each of these crosses were sown and the percentage of germination was 73 %. One hundred F1 plants were selected from each crossing, which were transferred to plastic bags, and then a part of these was taken to the field to perform the morphoagronomic character evaluation.

Figure 2 F1 lulo fruits. a. cv. Castilla Larga Vida χ cv. La Selva; b. cv. La Selva χ cv. Castilla Larga Vida.
The F1 plants of the crosses of the lulo cv. Castilla χ cv. La Selva, cv. La Selva χ cv. Castilla, and cv. La Selva χ cv. Larga Vida were sown in a plot located in the installations of UNAD, in the municipality of Dosquebradas, Risaralda. The study plots were located at an altitude of1,460 m a.s.l. with an average temperature of 21 °C, an annual rainfall of 2,200 mm and a relative humidity of 80 %. The region corresponds to a very humid premontane forest (bmh-PM) according to the Holdridge life zone classification.
Planting was carried out in a completely random design with 25 plants per treatment, for 100 plants in total. Every seven days the following quantitative morphoagronomic descriptors were obtained: plant height, stem diameter, fruit volume, number of leaves per week, number of buttons per cushion, number of cushions, number of fruits per bunch, number of bunches, and leaf area. From each of the treatments 10 rooted plants were plucked six months after sowing to evaluate level of root infestation by nematodes.
Resistance evaluation to nematodes
Nematode isolation
Pure cultures of M. incognita were obtained from infected roots of lulo cv. Castilla and from the soil surrounding these, using the Baermann funnel method.
Experiment set up for evaluation of resistance to Meloidogyne incognita
To evaluate the resistance of F1 plants to nematodes, seedlings with four or five well-formed leaves were planted in plastic jars with a capacity of 2 kg, with a substrate composed of three parts of soil and one part of organic matter without disinfecting.
A completely random design was used. The expe rimental unit corresponds to 10 plants, and the sampling unit was a complete plant by treatment and by repetition. Treatments correspond to the interaction of crosses by inoculation with or without nematodes, as follows.
T1: Fi plants of cv. Castilla Larga Vida χ cv. La Selva, without nematode inoculation
T2: Fi plants ofcv. Castilla Larga Vida χ cv. La Selva, inoculated with nematodes
T3: Plants of cv. Castilla Larga Vida, inoculated with nematodes (control)
T4: Fi plants of cv. La Selva χ cv. Castilla Larga Vida, inoculated with nematodes
T5: Fi plants of cv. La Selva χ cv. Castilla Larga Vida, without nematode inoculation
For plant inoculation, four holes were made in the soil inside each plot and around the neck of the root, where 50 cm3 of distilled water containing approximately 10,000 nematodes, between adults and eggs, were inoculated. These plants were kept inside a greenhouse for 60 days, at an average temperature of 23 °C and 85 % relative humidity.
At the end of this period, plants were extracted from their pots, and the presence or absence of nodes in the root system was evaluated, according to a scale of 0 to 10, where 0 corresponds to a healthy plant without root nodes, and 10 corresponds to plants with a root system completely covered in nodes. Plants were considered resistant when the mean root-node index was less than 2 and susceptible when that value was higher. Response variables used were level of infestation, root length and fresh root weight.
Nematode resistance evaluation
Multiplication of Neoleucinodes elegantalis
Adults of N. elegantalis were obtained from lulo fruits (from an undetermined cultivar) collected from plants in the area, which had entry holes but no exit holes. They were taken to the laboratory where four or five fruits were placed inside transparent plastic jars with wet sand and pieces of cloth paper in the bottom (figure 3). Once the larvae left the fruits, they started the pupation process on paper fabrics and adults emerged later. Adults obtained were sexed by wing color and by their genital opening.

Experiment set up for the evaluation of tolerance to Neoleucinodes elegantalis
To evaluate the tolerance of the F1 generations to N. elegantalis, entomological sleeves (40 χ 15 cm) were used, selecting a branch of the middle third of the plant, with fruits in the first formation phase, in which two male and two female N. elegantalis moths were introduced.
Daily observations were made in order to record oviposition. For the evaluation of tolerance to N. elegantalis, a completely randomized design of the progenies of the reciprocal crosses of lulo cv. Castilla Larga Vida χ cv. La Selva, and as control, plants from cv. Castilla Larga Vida were used. An experi mental unit corresponded to a plant with its respective entomological sleeve, and a sampling unit with 10 plants per treatment.
Four quantitative descriptors were used: percentage of affected fruits, average number of exit holes per fruit, percentage of fruits with exit holes, and average number of larvae per fruit. Data obtained from the experiments in relation to the evaluation of resistance to nematodes and tolerance to N. elegantalis were subjected to an analysis of variance (Anova) and mean comparison tests (Duncan) using the statistical program SPSS V15.0.
Results
Resistance evaluation of the F1 materials to the nematode Meloidogyne incognita
An analysis of variance for the variables level of infestation, root length and fresh weight of the root in seedlings 60 days after inoculation were carried out in function of the resistance of F1 plants of the reciprocal crosses of lulo cv. La Selva χ cv. Castilla Larga Vida to the nematode M. incognita; results showed highly significant differences between treatments.
Infestation level of Meloidogyne incognita in seedlings
When analyzing the level of infestation of the root by the nematodes in seedlings 60 days after inoculation, treatment 1 (F1 plants cv. Castilla Larga Vida χ cv. La Selva, without nematode inoculation) showed a level of nematode infestation of zero (0). Moreover, treatments 2 (F1 plants cv. Castilla Larga Vida χ cv. La Selva, inoculated with nematodes) and 5 (F1 plants cv. La Selva χ cv. Castilla Larga Vida, without nematode inoculation) showed a low percentage (2.5 %).
On the other hand, treatments 3 (cv. Castilla plants, inoculated with nematodes [control]) and 4 (F1 plants cv. La Selva χ cv. Castilla Larga Vida, inoculated with nematodes), were the most susceptible, with a level of infestation of 4.2 % and 4.7 %, respectively. Furthermore, 50 % of the plants of treatment 2 showed levels of infestation between 0 and 2 and can be suggested as resistant lines (figure 4).
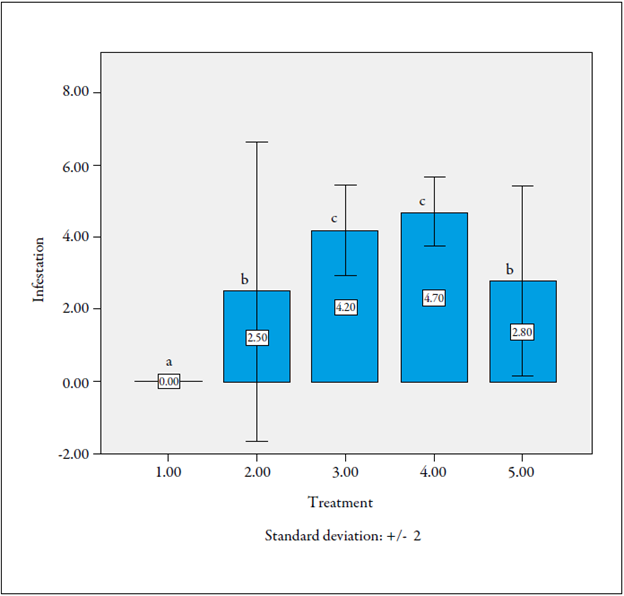
Source: Prepared by the authors
Figure 4 Evaluation of the level of infestation of the nematode Meloidogyne incognita in roots of seedlings of F1 lulo plants. Same letters on the bars mean that they do not differ statistically (p > 0.05).
Root length
There were significant differences among treatments. Roots of treatment 5 (F1 plants cv. La Selva χ cv. Castilla Larga Vida, without nematode inoculation) were the longest, with an average lenght of 40.5 cm, followed by those of this same material, but inoculated with nematodes (figure 5).
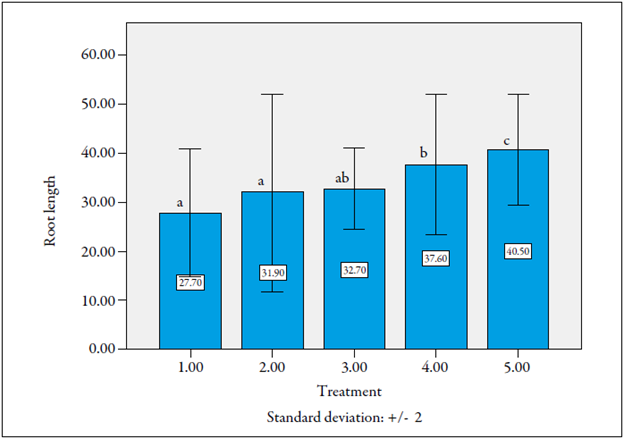
Figure 5 Root elongation affectation of F1 lulo plants by infestation of the nematode Meloidogyne incognita in seedlings. Same letters on the bars mean that they do not differ statistically (p > 0.05).
Source: Prepared by the authors Although in this case we can say that nematodes affected root lengthening, this did not occur in the F1 plants cv. Castilla Larga Vida χ cv. La Selva material. This might be due to an oligogenic effect provided by the parent cv. Castilla Larga Vida when it is used as a male (figure 5).
Fresh weight of roots
Fresh weight of roots alone did not prove to be a good susceptibility or tolerance indicator to the nematode M. incognita, although in treatment 4 (hibrid plants: cv. La Selva χ cv. Castilla Larga Vida, inoculated with nematodes) there was a highly significant difference of 39.3 g compared to all other treatments (figure 6).
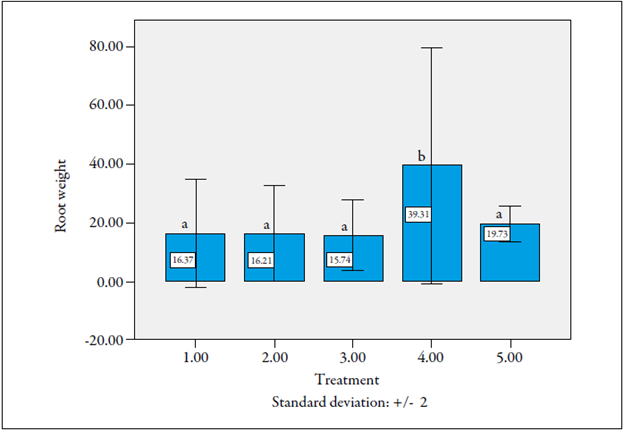
Source: Prepared by the authors
Figure 6 Root fresh weight in the seedling phase of F1 lulo plants evaluated for resistence to the nematode Meloidogyne incognita. Same letters on the bars mean that they do not differ statistically (p > 0.05).
A higher root weight found in this treatment may correspond to the hyperplastic effect of an infection by M. incognita, which increased root volume compared to those that were not infected, as reported in the literature (Hafez & Sundararaj, 2000; Hashmi, Huettel, Hammerschlag, & Krusberg, 1993, Perrotta, Magnano, & Bassi, 1980).
In addition, figure 7 shows the development of plants 60 days after the treatment was applied. In the first place, there is a lower plant vigor obtained from the crosses when the parent cv. Castilla Larga Vida was used as the mother; meanwhile the progenies of the crossing that used cv. La Selva as a mother showed a more vigorous development.

Control plants of cv. Castilla Larga Vida inoculated with M. incognita, in addition to showing a poor development, showed yellowing, i.e. symptom of the affectation by the nematode. Apparently, the F1 materials inoculated with M. incognita showed no symptoms of such affectation when plant infestation was evaluated.
From the previous results and subsequent from the comparison of treatments Ti and T5, we can conclude that there is no cytoplasmic effect on the resistance or tolerance to the nematode; however, this can be a result of an oligogenic effect, since a greater resistance is observed when the lulo cv. La Selva is used as a male parent.
Nonetheless, as stated by Dahms (1972) , it is necessary to bear in mind that this type of experiment, in which high inoculum pressures are exerted through the artificial inoculation of treatments, may mask the results, so it may be possible that we cannot detect if these F1 materials are really resistant.
Nevertheless, it should be considered that, because they are normally cross-pollinated plants, and since cv. La Selva is the result of a backcrossing, both parents are heterozygous, and segregation in the progeny obtained from crosses may be present. This causes that each plant shows differential levels of resistance to these crop pests.
When performing a correlation analysis between variables, we observed a non-significant inverse relationship between levels of nematode infestation and root weight, and a direct, non-significant relation ship between that level of infestation and root length. On the other hand, the correlation between root fresh weight and root length shows a direct and highly significant relationship, as is expected accor ding to the biological logic in plant development. However, this is a fact that does not account for the tolerance or susceptibility to nematodes (table 1).
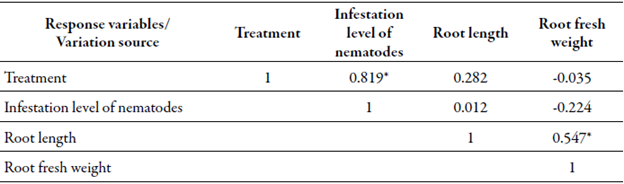
Infestation level of Meloidogyne incognita under normal field conditions
The F1 plants were evaluated under normal field conditions during the fruiting phase (six months after field transplantation). Because it was a destructive test, and since other phenological variables of the plants continued to be evaluated, only ten plants of each reciprocal crossing and of the evaluated cultivars were randomly removed; results show highly significant statistical differences between the materials related to the infestation ofM. incognita.
Based on the comparisons between the means of the evaluated materials regarding root infestation with M. incognita, the degree of infestation of both reciprocal crosses was less than 1 %, which indicates a high level of resistance of these materials to nematodes, e.g. cv. La Selva (0). On the other hand, cv. Castilla Larga Vida was highly susceptible (3.5 %) (figure 8).
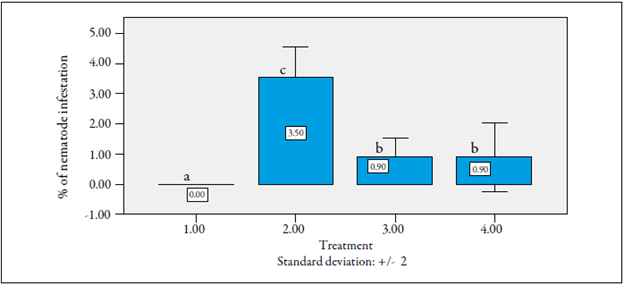
Evaluation of the resistance to Neoleucinodes elegantalis
The F1 materials of both reciprocal crosses of lulo cv. La Selva x cv. Castilla Larga Vida did not show N. elegantalis larvae inside the fruits, nor exit holes in these. On the contrary, plants of the cv. Castilla Larga Vida used as a control were largely affected (60 %) and showed, on average, two exit holes and two larvae per fruit (table 2).
Table 2 Average values of the evaluated descriptors to establish the resistance of the Fi plants of the crosses of lulo cv. La Selva x cv. Castilla Larga Vida to the moth Neoleucinodes elegantalis
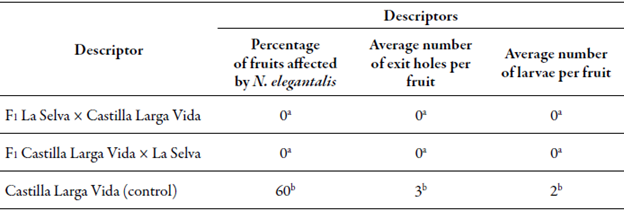
Averages with different letters in the same column are significantly different.
Source: Prepared by the authors
The high percentage of fruits affected and the high average of larvae that attacked each fruit in the control treatment indicated that, using the entomological sleeves, the level of infestation was adequate to evaluate the resistance of the F1 materials obtained with the reciprocal crosses of cv. La Selva x cv. Castilla Larga Vida to the nematode M. incognita.
The F1 materials showed resistance to the moth N. elegantalis, probably due to an antixenotic effect in the adult insect, based mainly on physical and chemical factors caused by the presence of trichomes throughout the plant and especially in the fruits, which secrete terpenoids, alkaloids, phenolic substances and others. These can be olfactory or gustatory repellents for the insect (Aragão, Dantas, & Gandolfi, 2000; Cardona, & Mesa, 2011; Simmons, & Gurr, 2005). However, it is necessary to continue with the observations of the lulo crop under commercial conditions.
Morphoagronomic behavior of hybrids
Hybrid plants of the reciprocal crosses proved to be vigorous, with an average height of 145 cm and a stem diameter of 4.5 cm, a good structure, large leaves, a length of 40.6 cm, a width of 41.5 cm, a petiole of 14 cm, and adapted to full solar exposure.
First flowering began 60 days after transplantation, with an average of four inflorescences per plant, which formed ca. 13 fruits, with an average diameter of 5.6 cm, similar to the lulo cv. Castilla Larga Vida. Fruit maturation and harvest occurred 155 days after the beginning of flowering.
Fruit epidermis was yellow and the pulp green, without dehiscence (cracking), and with pleasant aroma and flavor. However, plants showed abundant and pronounced thorns in stems, branches and leaves (figure 9), a condition that is classified as unfavorable by producers, since it hinders cultivation work.

Figure 9 F1 plants, four and a half months after transplantation to free sun exposure locations. a. Crossing of lulo cv. La Selva x cv. Castilla Larga Vida: stems and leaves with very strong thorns are observed in large quantity and with basal ramifications; b. Crossing of lulo cv. Castilla Larga Vida x cv. La Selva: shows thorns in stem and leaves.
Conclusions
When inoculated with a high level of infestation of the nematode M. incognita, the Fi plants obtained from the reciprocal crosses between the parents of lulo cv. Castilla Larga Vida and cv. La Selva turned out to be susceptible. However, when evaluating these same materials under field conditions, where the presence of the nematode was detected in previous samples, plants showed a root-node index lower than 1 %, which indicates that the materials are highly tolerant to the nematode, and that there is a possible segregation of this resistance character.
In seedlings, fresh weight of the root in inoculated plants was affected by the infestation of M. incognita, causing a reduction of its weight, which affects normal plant development. Although root growth was significantly affected in plants subject to a high inoculum pressure of M. incognita, superior vigor and higher tolerance were demonstrated in F1 plants, especially when the cv. Castilla Larga Vida was used as the mother parent.
Apparently, the F1 plants of the reciprocal crosses of cv. La Selva x cv. Castilla Larga Vida showed resistance to the moth N. elegantalis due to an antixenotic effect.
In subsequent trials, it is necessary to analyze separately fresh weight of roots and nodules of M. incognita, in order to establish real differences in weight attributable to the node formation caused by nematodes.
Moreover, it is necessary to continue with the evaluations of both the tolerance to nematodes and fruit quality (Brix degrees, total solids, ascorbic acid, among others), under commercial growing conditions, aiming at obtaining materials without thorns, but keeping the outstanding agronomic characteristics found in plants in this study, and those that have a high consumer acceptance.
How to cite this article: Polanco-Puerta, M. F., Gómez-Posada, S., & Padilla-Osorio, J. C. (2018). Resistance evaluation of a Fi hybrid of Solanum quitoense Lam. to Neoleucinodes elegantalis (Guenée) and Meloidogyne incognita (Kofoid & White). Corpoica Ciencia y Tecnología Agropecuaria, 19(2), 367-382.
Disclaimers: All co-authors made significant contributions to the document and agree with its publication. Moreover, all authors state that there are no conflicts of interest in this study.
Acknowledgements
The authors want to thank Universidad Nacional Abierta y a Distancia (unad) for their financial contributions to this research. Likewise, many thanks to farmers of the municipality of Belén de Umbría, Risaralda (Colombia), who allowed us to collect plant material in their farms. Moreover, we want to acknowledge the agronomist James Dávila, for his strong support during the fieldwork stage of this study, and also to our peer reviewers for their contributions that helped improve this manuscript.
REFERENCES
Andrés, M. (2002). Estrategias en el control y manejo de nematodos fitopárasitos. En Centro de Ciencias Medioambientales (ccma) (Ed.). Ciencia y Medio Ambiente - Segundas jornadas científicas sobre medio ambiente del CCMA-CSIC (pp. 221-227). Madrid, España: CCMA. [ Links ]
Aragão, C. A., Dantas, B. F., & Gandolfi, F. R. (2000). Tricomas foliares em tomateiro com teores contrastantes do aleloquímico 2-tridecanona. Scientia Agricola, 57(4), 813-816. [ Links ]
Bernal, J. A. & Díaz, D. (2006). Materiales locales y mejorados de tomate de árbol, mora y lulo, sembrados por los agricultores y cultivares disponibles para su evaluación en Colombia (Boletín divulgativo N.° 7). Bogotá, Colombia: Corporación Colombiana de Investigación Agropecuaria (Corpoica). [ Links ]
Bernal, J., Lobo, M., & Londoño, M. (1998). Documento de presentación del material Lulo La Selva. Rionegro, Colombia: Corporación Colombiana de Investigación Agropecuaria (Corpoica). [ Links ]
Cardona, C. & Mesa, N. (2011). Resistencia varietal a insectos. Palmira, Colombia: Universidad Nacional de Colombia. [ Links ]
Dahms, R. (1972). Tecniques in the evaluation and development of host-plant resistence. Journal of Environmental Quality, 1(3), 254-258. [ Links ]
Departamento Administrativo Nacional de Estadística (dane). (2015). El cultivo del lulo (Solanum quitoense) y los efectos del fenómeno del niño en la producción. Boletín mensual. Insumos y factores asociados a la producción agropecuaria, (42), 1-7. Recuperado de https://www.dane.gov.co/files/investigaciones/agropecuario/sipsa/Bol_Insumos_dic_2015.pdf. [ Links ]
Díaz-Montilla, A. E. (2013). Manejo integrado del gusano perforador del fruto de lulo y tomate de árbol (Boletín técnico). Rionegro, Colombia: Fondo Regional de Tecnología Agropecuaria (Fontagro), Corporación Colombiana de Investigación Agropecuaria (Corpoica). [ Links ]
Fory, P., Sánchez, I., Bohórquez, A., Ramírez, H., Medina, C., & Lobo, M. (2010). Variabilidad genética de la colección colombiana de lulo (Solanum quitoense Lam.) y especies relacionadas de la sección Lasiocarpa. Revista Facultad Nacional de Agronomía, 63(2), 5465-5476. [ Links ]
Franco, G., Bernal, E., Giraldo, C., Tamayo, M., Castaño, P., Tamayo, V., ... Ramírez, G. (2002). El cultivo de lulo (Manual técnico). Manizales, Colombia: Corporación Colombiana de Investigación Agropecuaria (Corpoica). [ Links ]
Hafez, S. L. & Sundararaj, P. (2000). Resistant reaction of alfalfa cultivars to the lesion nematode, Pratylenchus penetrans. The Journal of Nematology, 32, 433. [ Links ]
Hashmi, G., Huettel, R. N., Hammerschlag, F. A., & Krusberg, L. R. (1993). Optimal levels of Meloidogyne incognita inoculum for infection of tomato and peach in vitro. Journal of Nematology, 26(4), 531-534. [ Links ]
Instituto Colombiano Agropecuario (ica). (2011). Manejo fitosanitario del cultivo del lulo (Solanum quitoense Lam.). Medidas para la temporada invernal. Bogotá, Colombia: ICA. [ Links ]
Lobo, M. & Medina, C. (2000). Lulo (Solanum quitoense Lam.). En Caracterização de frutas nativas de América Latina. Serie: Frutas nativas de América Latina (Edição Comemorativa do 30° aniversário da Sociedade Brasileira de Fruticultura, pp. 41-43). Jaboticabal, Brasil: Fundação de Apoio a Pesquisa, Ensino e Extensão (Funep). [ Links ]
Marcano, R. V. (1991). Estudio de la biología y algunos aspectos del comportamiento del perforador del fruto del tomate Neoleucinodes elegantalis (Lepidoptera: Pyralidae) en tomate. Agronomía Tropical, 41(5), 257-263. [ Links ]
Maxwell, F. G. & Jennings, P. R. (1980). Breeding plants resistant to insects. Nueva York, EE. UU.: John Wiley & Sons. [ Links ]
Medina, C. I., Lobo, M., Martínez, E., & Riaños, N. (2004). Estudios fisiológicos del lulo en el bosque húmedo montano bajo del oriente antioqueño. I. Crecimiento y desarrollo. En Memorias del V Seminario Nacional e Internacional de Frutales. Manizales, Colombia: Universidad Nacional de Colombia. [ Links ]
Morton, J. (1987). Naranjilla. En J. F. Morton (Ed.). Fruits of warm climates (pp. 425-428). Miami, EE. UU.: Julia F. Morton. Recuperado de https://www.hort.purdue.edu/newcrop/morton/naranjilla_ars.html. [ Links ]
Muñoz, J. (2011). Análisis de la competitividad del sistema de la producción de lulo (Solanum quitoense Lam.) en tres municipios de Nariño (tesis de maestría). Universidad Nacional de Colombia, Bogotá, Colombia. Recuperado de http://www.bdigital.unal.edu.co/6333/1/790781.2011.pdf. [ Links ]
Obando, V. (2011). Variabilidad morfométrica de Neoleucinodes elegantalis (Guenée): perforador de fruto de solanáceas de importancia económica (tesis de maestría). Universidad Nacional de Colombia, Medellín, Colombia. Recuperado de http://www.bdigital.unal.edu.co/5346/1/37084704.2011.pdf. [ Links ]
Panda, N. & Khush, G. (1995). Host plant resistance to insects. Wallingford, Reino Unido: CAB International. [ Links ]
Parra, A., Barona, H., & Vallejo, F. A. (1994). Evaluación de especies silvestres de Lycopersicon sp. como fuente de resistencia al insecto plaga Scrobipalpula absoluta (Meyrick) y su intento de transferencia a la especie cultivada Lycopersicon esculentum Mill. Acta agronómica, 44(1-4), 51-61. [ Links ]
Perrotta, G., Magnano, G., & Bassi, M. (1980). Some anatomical and morphological-functional aspects of the resistance to Phoma tracheiphila in citrus plants. Journal of Phytopathology, 98, 346-358. [ Links ]
Schultes, R. & Cuatrecasas, J. (1953). Notes on the cultivated lulo. Botanical Museum Leaflets, 16(5), 97-105. [ Links ]
Simmons, A. T. & Gurr, G. M. (2005). Trichomes of Lycopersicon species and their hybrids: effects on pests and natural enemies. Agricultural and Forest Entomology, 7, 265-276. [ Links ]
Tamayo, P. (2001). Principales enfermedades del tomate de árbol, mora y lulo en Colombia (Boletín técnico N.° 12). Rionegro, Colombia: Corporación Colombiana de Investigaciones Agropecuarias (Corpoica). [ Links ]
Whalen, M., Costich, D., & Heiser, C. (1981). Taxonomy of Solanum section Lasiocarpa. Gentes Herbarum, 12(2), 41-129. [ Links ]
Received: August 31, 2016; Accepted: February 05, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
